Abstract
Currently, the search for new promising tools of immunotherapy continues. In this regard, microRNAs (miRNAs) that influence immune checkpoint (IC) gene expression in tumor and T-cells and may be important regulators of immune cells are considered. MiRNAs regulate gene expression by blocking mRNA translation. An important feature of miRNA is its ability to affect the expression of several genes simultaneously, which corresponds to the trend toward the use of combination therapy. The article provides a list of miRNAs acting simultaneously on several ICs and miRNAs that, in addition to IC, can regulate the expression of targeted therapy genes. There is dependence of miRNA interactions with IC genes on the type of cancer. The analysis of the accumulated data demonstrates that only about 14% (95% CI: 9.8–20.1%) of the studied miRNAs regulate the expression of specific IC in more than one type of cancer. That is, there is tumor specificity in the miRNA action on ICs. A number of miRNAs demonstrated high efficiency in vitro and in vivo. This indicates the potential of miRNAs as promising agents for cancer immunotherapy. Additional studies of the miRNA–gene interaction features and the search for an optimal miRNA mimic structure are necessary.
1. Introduction
Immunotherapy is an innovative method of cancer treatment. As a result of experiments and clinical trials, it has been found that immunotherapy can increase progression-free survival and overall survival. However, this method of treatment is effective in a limited number of patients, and in addition, it can cause severe adverse reactions due to hyperreactivity of the immune system [1]. In this regard, research is underway to develop new therapeutic approaches based on targeting immune checkpoints (ICs).
ICs are regulators of the immune system, which are divided into stimulating and inhibitory molecules. Stimulating and inhibitory ICs regulate T-cell activation. Tumor cells have the ability to generate ligands that can bind to co-inhibitory receptor molecules. This interaction suppresses the antitumor immune response, allowing the tumor to “escape” from the immune system. As an opportunity to solve this problem, the blockade of ICs is used [2].
IC inhibitors are monoclonal antibodies that affect a specific target [3]. In order to increase the effectiveness of immunotherapy, the FDA approved a number of regimens, including a combination of two IC inhibitors, a combination of IC inhibitors and targeted therapy drugs, as well as antitumor bispecific antibodies [4,5]. It has been shown that in combination therapy regimens, patients experienced a higher response rate compared to monotherapy [6].
In addition, the search for a more promising immunotherapy approach is currently ongoing. In this regard, microRNAs (miRNAs) are considered. MiRNAs participate in tumor-cell signaling pathways and regulate many processes, including the antitumor immune response.
MiRNAs are small non-coding RNAs that carry out post-transcriptional regulation of gene expression. According to recent studies, miRNAs influence IC gene expression and are important regulators in both T-cells and tumor cells [7]. MiRNAs regulate gene expression by binding to the 3’-UTR of their mRNA [8,9,10]. MiRNAs can also affect IC expression indirectly, through molecules of different signaling pathways, such as PTEN, IFR-1, and others [11]. It is also important that one miRNA can affect several genes [7,12].
This article presents a review of miRNAs that interact with IC genes, analyzes their regulating IC expression in tumors of various types of cancer, and identifies miRNAs that act on several IC genes simultaneously. Due to these properties, miRNA-based therapy may become an alternative to the combination of targeted drugs in the future. The effect of miRNAs on the same IC gene in tumors of several types was analyzed. Thus, the tumor specificity of the miRNA–IC gene interaction was assessed. In addition, miRNAs are considered that are capable of simultaneously regulating the expression of targeted therapy genes along with IC genes. These issues have not been previously analyzed in existing reviews of miRNAs as IC regulators [13,14,15,16,17].
2. Immune Checkpoints
2.1. PD-1, PD-L1 and CTLA-4
Programmed cell death protein 1 (PD-1) and programmed death-ligand 1 (PD-L1) are the most studied members of the ICs. PD-1 and CTLA-4 receptors are co-inhibitory molecules. They are normally expressed on activated T-lymphocytes. The PD-1 receptor has PD-L1 and PD-L2 ligands, and the CTLA-4 receptor has CD80/CD86 ligands that can be expressed on tumor cells in various types of cancer. The interaction of PD-1 and CTLA-4 receptors with their ligands leads to suppression of the cytolytic activity of T-lymphocytes, which blocks antitumor immunity. The use of PD-1 and CTLA-4 checkpoint inhibitors increased patient survival compared with traditional chemotherapy in a number of studies, including studies on kidney cancer, melanoma, head and neck squamous cell cancer (HNSCC) and non-small cell lung cancer [18].
Based on preclinical and clinical trials, the FDA approved the use of PD-1 inhibitor-nivolumab in combination with CTLA-4 inhibitor (ipilimumab) for the treatment of several types of metastatic cancer [19].
2.2. Gal-9/Tim-3
One of the relatively new targets of immunotherapy is the T-cell immunoglobulin and mucin domain-3 (Tim-3)/Galectin-9 (Gal-9) pathway. Tim-3 is present on activated effector T-cells. It is an immunosuppressive receptor, causing exhaustion of T-cells [20]. A high level of Tim-3 expression in kidney cancer is associated with an unfavorable prognosis [21,22]. One of the ligands of Tim-3 is Gal-9, which belongs to the galectin family. It is reported that this family of proteins regulates tumor proliferation, migration, and metastasis [23].
Kidney, brain, colon, blood, liver, prostate, lung, and skin cancer cell lines were found to express detectable amounts of Tim-3 and Gal-9 proteins. It is assumed that the Tim-3/Gal-9 pathway is involved in the prevention of antitumor immunity [24]. However, the information regarding Gal-9 is contradictory. According to a number of authors, Gal-9 overexpression correlates with a poor prognosis in many types of cancer [25,26,27,28,29]. On the other hand, increased expression of Gal-9 has been shown to be associated with a favorable prognosis in solid tumors [30].
Currently, Gal-9 is considered as a target of immunotherapy, anti-Gal-9 antibodies have been developed. In the co-cultivation of T-cells and tumor cells, these antibodies contributed significantly to T-cell-mediated destruction of tumor cells [31].
In addition, considering Gal-9 as a target for immunotherapy, it should be taken into account that Gal-9 also interacts with stimulatory receptors, such as the 4-1BB-co-stimulating receptor of the tumor necrosis factor receptor superfamily (TNFRSF). Another member of the TNFRSF—GITR—may also be relevant for the results of Gal-9 inhibition [25].
2.3. VISTA
V-domain Ig-containing suppressor of T-cell activation (VISTA) is an immunosuppressive receptor. It is considered as a potential target of immunotherapy. VISTA expression level is significantly increased in clear cell renal cell carcinoma (ccRCC). In animal models of kidney cancer, VISTA blockade significantly suppressed tumor growth [32].
In addition, it was shown that the combination of a VISTA inhibitor with an TLR agonist led to the development of antitumor immunity associated with T-cells [33]. It is also reported that simultaneous blockade of CTLA-4 and VISTA can enhance the antitumor immune response in HNSCC [34].
2.4. BTLA
The B- and T-lymphocyte attenuator (BTLA) is expressed by most lymphocytes. BTLA belongs to the CD28 superfamily and is similar in structure and function to PD-1 and CTLA-4 [35]. Increased BTLA expression is associated with an unfavorable prognosis [36]. The BTLA ligand is HVEM (TNFRSF14), a mediator of herpes virus penetration, a membrane protein that belongs to the superfamily of tumor necrosis factor receptors. The BTLA/HVEM axis is one of the most important ICs [37].
2.5. B7-H3
B7-H3 (CD276) is a member of the B7 family. It functions as a co-stimulating and as a co-inhibiting immunoregulatory protein [38]. B7-H3 is not only a regulator of the antitumor immune response, but is also involved in angiogenesis and metastasis. Co-expression of B7-H3 and tyrosine kinase receptor of angiopoietin Tie-2 was detected in RCC. It was shown that overexpression of B7-H3 and Tie-2 in the vascular endothelium of RCC was associated with the density of tumor microvessels and disease progression [39]. Additionally, it was found that B7-H3 knockdown eliminated the pro-metastatic effect of fibronectin and significantly suppressed the metastasis of ccRCC cells [40].
Clinical trials have evaluated the effectiveness of B7-H3 inhibitors in the treatment of solid tumors, both as monotherapy and as part of combined therapy [41]. In particular, a bispecific anti-B7-H3/PD-1 fusion protein has been developed. It interacts simultaneously with the tumor-associated marker B7-H3 and the immunosuppressive signaling pathway PD-1/PD-L1, and also enhances antibody-dependent cellular cytotoxicity. Treatment with anti-B7-H3/PD-1 fusion protein leads to effective suppression of tumor growth in animal models of several types of cancer [42]. A bispecific antibody targeting B7-H3 and 4-1BB (BsAb; B7-H3 × 4-1BB) has also been developed, which is a B7-H3 inhibitor and a 4-1BB agonist. It has been shown that BsAb; B7-H3 × 4-1BB, as well as its combination with anti-PD-1 therapy, inhibits tumor growth in animal models of several types of cancer [43].
2.6. ICOS/ICOSL
Inducible co-stimulator (ICOS), a molecule that also belongs to the CD28/CTLA-4/B7 immunoglobulin superfamily. ICOS and its ligand (ICOSL) are involved in various aspects of the T-cell response. The ICOS/ICOSL pathway has been shown to play a significant role in anti-CTLA-4 therapy [44]. ICOS/ICOSL is considered as a target for kidney cancer in combination with anti-PD-L1 therapy (NCT03829501) [45].
2.7. LAG3
Lymphocyte-associated gene 3 (LAG3) is a transmembrane protein that is expressed on immune cells. LAG3 refers to immunosuppressive IC molecules. LAG3 overexpression is associated with overall survival, as is the overexpression of PD-1 and CTLA-4. Currently, bispecific antibody immunotherapy trials that aim to simultaneously inhibit LAG3 and PD-1/PD-L1/CTLA-4 are being conducted [46]. According to Zelba et al., simultaneous blocking of PD-1 and LAG3 is a promising strategy in the treatment of kidney cancer [47]. Fibrinogen-like protein 1 (FGL1) participates in inactivation of T-cells and is considered as the main ligand of LAG3 [48].
3. Simultaneous Inhibition of ICs and Other Targets
3.1. ICs and VEGF
Combined treatment regimens, including IC inhibitors and anti-VEGF therapy, have shown significant efficacy in patients with various types of cancer [49,50,51]. To date, antiangiogenic therapy in combination with anti-PD-1/PD-L1 (Pembrolizumab plus Axitinib, Nivolumab plus Cabozantinib and Pembrolizumab plus Lenvatinib) is recommended by the NCCN and EAU along with anti-PD-1/anti-CTLA-4 treatment (Ipilimumab plus Nivolumab as first-line therapy for metastatic RCC [52]).
3.2. ICs and c-Met
c-Met is a receptor tyrosine kinase that is involved in normal cell development and motility. Aberrant activation of c-Met can lead to tumor growth and metastasis [53]. The combination of ICI with cabozatinib, which inhibits c-Met in addition to VEGFR, is now recognized as the new standard of care for metastatic RCC [54]. There is also a study evaluating the efficacy of a combination of c-Met inhibitor and anti-PD-1 therapy in locally advanced or metastatic hepatocellular carcinoma and RCC (NCT03655613; NCT02795429).
3.3. ICs and HIF
Co-suppression of IC and Hypoxia-inducible factor (HIF) is also being considered as a therapeutic strategy. HIF activates downstream effectors, including vascular endothelial growth factor (VEGF), platelet growth factor (PDGF) and carbonic anhydrase IX (CA IX), which are involved in cell proliferation, angiogenesis, and erythropoiesis [55]. HIF-2α is considered as a target for therapy in RCC. The combination of a HIF-2α inhibitor (belzutifan), an anti-VEGF therapy (lenvatinib), and PD-1 and CTLA-4 inhibitors is currently being investigated in patients with RCC (NCT04736706).
3.4. ICs and PI3K
The efficacy of a combination of PD-L1 inhibitors (atezolizumab) and VEGF (bevacizumab) and a selective phosphoinositide 3-kinase (PI3K)-gamma inhibitor (IPI-549) in the treatment of metastatic RCC is currently being investigated (NCT03961698). Clinical trials are also underway to evaluate the effectiveness of combined inhibition of BRAF and MEK (MAPK and PI3K-Akt-mTOR signaling pathways) in combination with PD-1/PD-L1 blockade in melanoma [56].
3.5. ICs and CXCR4
Overexpression of CXC chemokine receptor 4 (CXCR4) is observed in many types of cancer and is associated with a poor prognosis [57]. CXCR4 is also considered as a promising therapeutic target [58]. A number of clinical trials of the combination of a CXCR4 inhibitor with anti-PD-1/PD-L1 therapy are currently underway in several types of cancer [4].
3.6. ICs and EGFR
EGFR expression is associated with the progression of many types of cancer. EGFR is one of the therapeutic targets. It has been shown that EGFR is expressed in 98.4% of cases in ccRCC [59]. A bispecific antibody that inhibits PD-1 and EGFR (anti-PD-1 x anti-EGFR) has been shown to significantly suppress tumor growth and activate antitumor immunity in animal models of various types of cancer [60].
3.7. ICs and HER2
Overexpression of the human epidermal growth factor receptor 2 (HER2) occurs in breast and gastric cancer (GC) and is associated with a poor prognosis. Currently, clinical trials are underway to evaluate the effectiveness of a combination of IC and HER2 inhibitors at different stages of the disease [61,62]. A bispecific antibody that simultaneously inhibits PD-1 and HER2 demonstrated significant efficacy in both in vitro and in vivo experiments [63].
4. Regulation of IC Genes by miRNAs
Currently, a lot of miRNAs are known to interact with ICs in various types of cancer. Table 1 lists miRNAs that interact with ICs and the type of cancer in which this interaction was shown. More than 50 miRNAs regulate the PD-1/PD-L1 pathway, and about 40 miRNAs regulate B7-H3. For other ICs, fewer miRNA regulators have been described.

Table 1.
The miRNAs interacting with IC genes in different types of cancer.
In particular, miR-497-5p is a direct inhibitor of PD-L1 in kidney cancer. An inverse correlation was noted between miR-497-5p and PD-L1 expression levels in ccRCC samples. In addition, reduced miR-497-5p expression was associated with shorter survival. In vitro experiments demonstrated that miR-497-5p suppressed tumor cell proliferation and migration, simultaneously stimulating their apoptosis [71].
Other miRNAs also can bind to the 3’-UTR of the PD-L1 gene and suppress PD-L1 expression. These include miR-570, miR-34a, miR-200, miR-21, and miR-197. Thus, PD-L1 may be the main target for miRNA control of ICs [7].
MiR-138 and miR-28 inhibit PD-1 expression in T-cells. MiR-138 can enhance immune response and slow down tumor progression in mouse [109]. A low level of miR-28 induces T-cell exhaustion and allows tumor cells to evade immune surveillance in a mouse melanoma model [112].
The impact of miRNAs on other ICs may also be important for the development of new approaches in cancer therapy. MiR-448 and miR-153 inhibit IDO1 in CRC. MiR-448 activates CD8+ T-cells by inhibiting function of IDO1 enzyme [115].
MiR-153 inhibits IDO1 expression in CRC cells; however, overexpression of this miRNA does not have a significant effect on tumor cells. However, overexpression of miR-153 has been shown to enhance the effect of chimeric antigen receptor (CAR) T-cell therapy [116].
MiR-128 is a direct inhibitor of Gal-3. Reduced miR-128 expression in CRC was negatively correlated with Gal-3 expression and was associated with poor prognosis. MiR-128 overexpression increased tumor cell sensitivity to chemotherapy in experiments in vitro and in vivo [118].
MiR-498 is an inhibitor of Tim-3. In experiments in AML cell lines, miR-498 significantly suppressed Tim-3 expression, which led to a decrease in proliferation and an increase in cell apoptosis [114].
B7-H3 inhibitors include miR-145, miR-1301-3p, miR-335-5p, miR-28-5p and miR-187. B7-H3 and miR-145 have been shown to be associated with lymph node metastasis, grade, and TNM stage in lung cancer with malignant pleural effusion [124]. MiR-1301-3p, miR-335-5p and miR-28-5p downregulate B7-H3 and are associated with lymph node metastasis and TNM staging in CRC [125]. MiR-187 is also a direct inhibitor of B7-H3. Reduced miR-187 expression has been shown to be associated with TNM stage in kidney cancer. Overexpression of miR-187 resulted in decreased proliferation and migration of tumor cells in vitro and inhibited tumor growth in vivo [130].
The direct target of miR-32 is BTLA. Experiments in OC cell lines showed that miR-32 overexpression led to a decrease in BTLA expression, resulting in a significant suppression of tumor cell proliferation, migration, and invasion [113].
One miRNA can target several ICs in the same type of cancer (Figure 1). For example, miR-28 interacts with PD-1/PD-L1, BTLA, and Tim-3 in melanoma [112]; miR-424 interacts with PD-1/PD-L1 and CD80/CTLA-4 in OC [108]; miR-128 interacts with B7-H3 and Gal-3 in CRC [118,127]; miR-138 interacts with PD-1 and CTLA-4 in glioma [109].
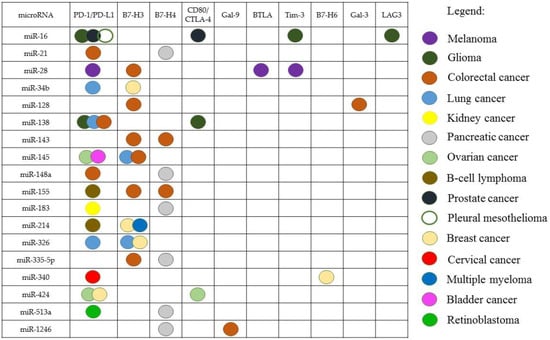
Figure 1.
The miRNAs interacting with several IC genes in some types of cancer.
The spectrum of targets of the same miRNA can be different in different types of cancer. In particular, miR-155 interacts with PD-1/PD-L1 in B-cell lymphoma and with B7-H3, B7-H4 in CRC [84,126]; miR-145 acts on PD-1/PD-L1 in OC and bladder cancer [96,97], meanwhile, in lung cancer and CRC, this miRNA acts on B7-H3 [124,126].
It has also been shown that miRNA can interact with the IC gene, in one type of cancer and not in another. An interaction of miR-34 with PD-L1 has been found in lung cancer, but it has not been seen in pleural mesothelioma [81]. On the other hand, the interaction of a certain miRNA with the IC can be observed in several types of cancer: for example, miR-138 inhibits the PD-1/PD-L1 pathway in glioma, CRC, and lung cancer [68,72,109]
The miRNAs that regulate ICs do so depending on cancer type: there is almost no overlap between these miRNAs in different types of cancer (Figure 2). For example, 60 miRNA regulators of ICs have been identified for PC and 25 for BC. However, among them, no miRNAs have been identified in both PC and BC. The same picture is observed in many other types of cancer. Among 115 miRNAs shown in the diagram, only 15 miRNAs (13%) interact with the IC genes in two or more types of cancer.
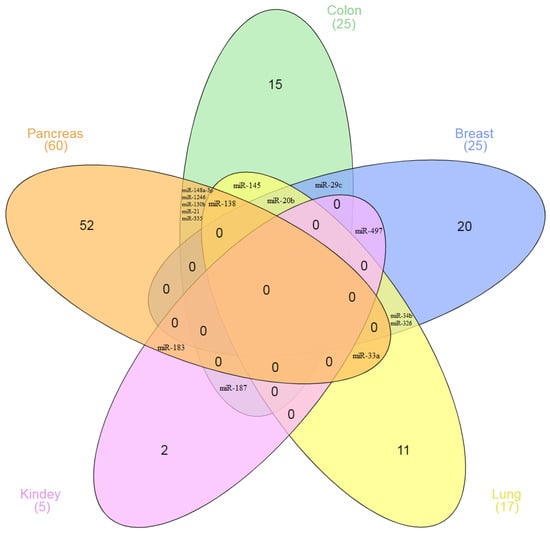
Figure 2.
The Venn diagram of the number of miRNAs interacting with IC genes in various types of cancer.
The confidence interval (CI) for the obtained ratio (15/115) was determined. The 95% CI was from 7.3% to 21.5%. The diagram (Figure 2) shows miRNAs in five types of cancer, which are distributed with the specified characteristics. In total, we have considered 207 miRNAs in the article, 30 of which regulate IC in different types of cancer (Table 1). The analysis of the data demonstrates the only 14.5% (95% CI: 9.8–20.1%) of the studied miRNAs regulate the expression of specific IC in more than one type of cancer. That is, when the number of analyzed miRNAs increased by almost two times, the share of overlapping miRNA and confidence intervals practically did not change.
For the first time, on a large sample of miRNAs, we showed the tumor specificity of their action on the IC genes. Similar results were obtained both in the study of five types of cancer and in the study of more of them. These calculations show that the number of miRNAs studied is sufficient for reliable statistical evaluation. Similar results are also observed in separate experiments. Thus, in the work of Qian et al., 62 miRNAs associated with B7-H4 in pancreatic cancer were identified using the miRCURY LNA™ microchip. Of these miRNAs, only 8 (13%) regulate ICs in other cancers [143]. In lung cancer, it was shown that miR-34 directly interacts with the 3’-UTR of PD-L1; miR-34 overexpression suppressed PD-L1 protein expression [87]. At the same time, in pleural mesothelioma, Kao et al. did not observe the relationship between the expression of miR-34 and PD-L1 [81]. Thus, our conclusion about the tumor-specific interaction of miRNA with ICs is consistent with the results of studies by other authors. Essentially, miRNAs can be differentially expressed depending on cancer type. Korotaeva et al. found that miRNAs are specifically expressed in certain types of neuroendocrine tumors [145].
5. miRNAs as Regulators of ICs and Targeted Therapy Genes
In addition to ICs, miRNAs can also affect other genes (such as mediators of angiogenesis, hypoxia, etc.) and thus regulate different signaling pathways (Table 2). In other words, miRNAs function as part of a complex signaling network [146].
Table 2.
MiRNAs regulating immune checkpoints as well as other cancer relevant genes.
In particular, in CRC, a number of miRNAs may affect both ICs and other genes that play a key role in cancer development. For example, miR-145 inhibits its target B7-H3, miR-143 is a direct inhibitor of B7-H3 and B7-H4. Also, these miRNAs suppress the angiogenesis regulator VEGF, HIF1 and the IRS-1/IGF-IR signaling pathway, which is a potent inhibitor of apoptosis and cell differentiation [126,157,158,159].
MiR-143 indirectly interacts with miR-155. The miR-155 targets C/EBPβ, a transcriptional activator for miR-143. MiR-155, by inhibiting miR-143, has been shown to increase the expression of B7-H3 and B7-H4 [126]. Also, miR-155 is described as a direct inhibitor of HIF-1α, and thus, being a regulator of various signaling pathways, miR-155 functions as an oncomiR and a tumor suppressor [156].
MiR-148a, in addition to PD-L1, inhibits the human epidermal growth factor receptor 3 (HER3), Wnt/β-catenin pathway ligand—WNT10b, apoptosis regulator—BCL2, as well as VEGF and HIF1 in CRC; that is, it acts as a tumor suppressor [73,164,165,166,167].
In addition, miR-148a has been shown to inhibit the calnexin (CANX)/MHC-I signaling pathway in CRC, by downregulating its direct target, CANX. An increased level of CANX expression in the tumor positively correlates with the overall survival of patients with CRC. Thus, miR-148a functions as a tumor promoter in this case [168].
In CRC miR-1246 inhibits Gal-9 and CCNG2, which is a tumor suppressor; decreased expression of CCNG2 occurs in many types of cancer and correlates with lymph node metastasis, clinical stage, and poor prognosis [121,163].
One miRNA can be involved in several signaling pathways that mediate different, sometimes opposite, cell functions. This can lead to the development of adverse events of miRNA-based therapy. Thus, when considering miRNAs as therapeutic targets, the features of their action in a particular type of cancer should be taken into account.
6. Prospects for miRNA-Based Therapy
Currently, a number of therapeutic approaches involving miRNAs are being developed: combinations of miRNAs with other agents [173,174,175,176,177], as well as combinations of two miRNAs [178,179,180] are being investigated.
MiRNAs from the miR-200 family are inhibitors of PD-L1 [90], HIF-1α, and the VEGF pathway [181,182,183]. According to Nguyen et al., nanoparticle therapy containing miR-200c as a PD-L1 inhibitor with a BRAF inhibitor demonstrated significant efficacy in a mouse model of CRC [177].
MiR-15/16 cluster miRNAs have been shown to interact with several genes. In prostate cancer, a negative correlation was found between the expression level of miR-195 and miR-16 and PD-L1, PD-1, CD80, and CTLA-4 [86]. In pleural mesothelioma, miR-15/16 inhibited PD-L1, Bcl-2, and CCND1. Injection of miR-16 mimetics led to a decrease in Bcl-2 and CCND1 expression levels and also inhibited tumor growth in animal models of malignant pleural mesothelioma [81,147]. The efficacy of miR-16 mimetic was evaluated in phase I clinical trials in patients with malignant pleural mesothelioma. The trial results were described as promising and suggest further drug research [184,185].
Experiments on T-cells have shown that direct targets for miR-155 are CTLA-4 and BTLA [186,187]. Administration of antibodies against CTLA-4, PD-1, and PD-L1 activated the antitumor immune response in miR-155 knockout mice [188]. MiR-155 also indirectly inhibited CXCR4 in glioma [156,189]. In chronic lymphocytic leukemia, diffuse large B-cell lymphoma, and liposarcoma, miR-155 acts as an oncomiR by inhibiting its direct targets, the casein kinase CSNK1G2 and the casein kinase 1α (CK1α) isoforms [190,191]. Also, miR-155 upregulates PD-L1 expression [84] and regulates a number of signaling pathways (including JAK/STAT, MAPK/ERK, and PI3K/AKT) in lymphoma [192]. The miR-155 inhibitor has performed well in the clinical trials NCT02580552, NCT03837457 and NCT03713320 in patients with hematologic malignancies in which miR-155 is overexpressed, such as cutaneous T-cell lymphoma, mycosis fungoides, chronic lymphocytic leukemia, diffuse large B-cell lymphoma, ABC subtype, adult T-cell leukemia/lymphoma [193].
MiR-138 acts as a tumor suppressor in many types of cancer and inhibits many target genes. Overexpression of miR-138 may increase the sensitivity of tumor tissue sensitivity to chemotherapy [194]. In lung cancer, miR-138 has been shown to significantly inhibit tumor cell proliferation in vitro by acting on its target, the PD-L1/PD-1 pathway. In experiments on NSCLC xenografts, miR-138 not only inhibited tumor growth, which led to a decrease in its size, but also regulated the tumor microenvironment [68]. In CRC, transfection with miR-138-5p mimetics also led to a decrease in tumor cell proliferation. In experiments on xenografts with the administration of miR-138 tumor size was decreased [72]. In glioma models, miR-138 inhibited CTLA-4 and PD-1, leading to significant regression of subcutaneous tumors. Moreover, tumor regression continued even after treatment was stopped. The administration of miR-138 was also effective in animals with intracerebral tumor localization: the average lifespan of mice treated with miR-138 was 33.5 days, and in the control group it was 23.5 days [109].
MiR-424 is a direct inhibitor of PD-L1 and CD80. It was shown that miR-424 expression in OC tumors negatively correlated with the PD-L1 and CD80 expression level. Also, miR-424 overexpression was correlated with progression-free survival. In experiments in animal models of OC, restoration of miR-424 expression increased tumor sensitivity to chemotherapy. Administration of miR-424 led to tumor regression and decreased tumor cell chemoresistance due to the activation of the T-cell immune response [108].
MiR-142-5p is a direct inhibitor of PD-L1. In vitro experiments revealed that miR-142-5p overexpression did not affect the tumor cell proliferation. However, miR-142-5p has been shown to inhibit the growth of PC and enhance antitumor immunity in vivo [69].
It has been shown that miR-34a inhibits PD-L1 and reduces the proliferation and migration of BC cells [195]. Overall, more than 30 target genes have been described for miR-34a involved in various signaling pathways in cancer. MiR-34a liposomal mimic, MRX34, is the first-in-human miRNA-based drug that has been evaluated in clinical trials in patients with various solid tumors [196]. This trial was terminated due to serious immune-mediated adverse events that may indicate the effect of this miRNA on several immune system regulatory genes. Despite this, miR-34a continues to be considered as a therapeutic target in cancer, as it has significant tumor suppressor potential. It was shown that the induced co-expression of miR-34a with other miRNAs led to a pronounced and stable therapeutic effect in models of various types of cancer, such as CRC, NSCLC, melanoma, etc. [197]. Orellana et al. identified five miRNAs that, in combination with miR-34a, most effectively inhibited tumor cell proliferation [198]. The combination of miR-34a-mimic and antisense-miR-10b is also being studied in BC models [180]. Meng et al. showed that the synthesized miR-34a analog NS-MX3 simultaneously decreased the expression of B7-H3 and PD-L1 and demonstrated superior antitumor activity in CRC models in vitro and in vivo [199].
Thus, the results of in vitro and in vivo experiments on the study of miRNAs as immunotherapeutic agents look encouraging.
In order to use miRNA-based drugs in clinical practice, these drugs must meet the requirements for efficacy and safety. Techniques that provide more specific binding to a target can increase the effectiveness of miRNA-based therapy, as well as reduce the dose of the administered drug, thereby reducing its side effects.
7. Conclusions
We have reviewed more than 200 miRNAs that regulate ICs in tumors of various types. The results of accumulated data analysis for the first time demonstrate a significant relationship between the action of miRNAs on ICs genes and the type of tumor—only about 14% (95% CI: 9.8–20.1%) of the studied miRNAs regulate the expression of specific IC in more than one type of cancer. That is, there is tumor specificity in the miRNA action on ICs.
An important feature of miRNA is its ability to affect the expression of several genes simultaneously. The data described here evidenced that some miRNAs can simultaneously regulate more than one IC gene. In addition, there are miRNAs that can affect both the IC gene and some targeted therapy genes. These results indicate the possibility of using miRNAs in the future as an alternative to combined treatment regimens that use inhibition of both two ICs simultaneously and inhibition of ICs together with targeted therapy genes.
Currently, there are numerous studies underway to identify miRNAs that are the most promising as immunotherapy agents. In vivo experiments have repeatedly shown that miRNA-based therapy leads to significant tumor regression.
Although miRNA has not yet entered the arsenal of antitumor agents used in practice, some results are encouraging. Thus, the miR-155 inhibitor has performed well in clinical trials. The study of miR-138 is promising. Ongoing research on miR-34a may also lead to a positive result. Thus, there is the prospect of using miRNA as a therapeutic agent in cancer immunotherapy regimens.
At the same time, the ability of miRNAs to inhibit several genes can lead to adverse events. To overcome this, it is important to expand data of the spectrum of miRNA targets in a particular type of cancer. Additional studies of the miRNA–genes interaction features and the search for an optimal miRNA mimic structure are necessary, thus allowing an increase in the efficiency and selectivity of interaction with the mRNA of target genes. It can increase the effectiveness of therapy, as well as reduce the dose of the drug, thereby reducing its side effects.
Author Contributions
Conceptualization, F.K. and A.K. (Alexander Karpukhin); methodology, F.K. and A.K. (Alexandra Korotaeva); formal analysis, I.S.; resources, M.N. and O.M.; writing—original draft preparation, F.K. and T.M.; writing—review and editing, D.M.; visualization, D.M. and P.A.; supervision, A.K. (Alexander Karpukhin). All authors have read and agreed to the published version of the manuscript.
Funding
The research was carried out within the state assignment and funding of the Ministry of Science and Higher Education of the Russian Federation FGFF-2022-0011.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The cited works can be found on the PubMed, PMC, Omicsonline, and Embase databases.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tan, S.; Li, D.; Zhu, X. Cancer immunotherapy: Pros, cons and beyond. Biomed. Pharmacother. 2020, 124, 109821. [Google Scholar] [CrossRef]
- Qin, S.; Xu, L.; Yi, M.; Yu, S.; Wu, K.; Luo, S. Novel immune checkpoint targets: Moving beyond PD-1 and CTLA-4. Mol. Cancer 2019, 18, 155. [Google Scholar] [CrossRef]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. AAPS J. 2021, 23, 39. [Google Scholar] [CrossRef]
- Vafaei, S.; Zekiy, A.O.; Khanamir, R.A.; Zaman, B.A.; Ghayourvahdat, A.; Azimizonuzi, H.; Zamani, M. Combination therapy with immune checkpoint inhibitors (ICIs); a new frontier. Cancer Cell Int. 2022, 22, 2. [Google Scholar] [CrossRef]
- Esfandiari, A.; Cassidy, S.; Webster, R.M. Bispecific antibodies in oncology. Nat. Rev. Drug Discov. 2022, 21, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Jin, H.; Guo, K.; Xiang, Y.; Zhang, Y.; Du, W.; Shen, M.; Ruan, S. Results from a Meta-analysis of Combination of PD-1/PD-L1 and CTLA-4 Inhibitors in Malignant Cancer Patients: Does PD-L1 Matter? Front. Pharmacol. 2021, 12, 572845. [Google Scholar] [CrossRef]
- Zhang, Y.; Tanno, T.; Kanellopoulou, C. Cancer therapeutic implications of microRNAs in the regulation of immune checkpoint blockade. ExRNA 2019, 1, 19. [Google Scholar] [CrossRef]
- Shao, L.; He, Q.; Wang, J.; He, F.; Lin, S.; Wu, L.; Gao, Y.; Ma, W.; Dong, J.; Yang, X.; et al. MicroRNA-326 attenuates immune escape and prevents metastasis in lung adenocarcinoma by targeting PD-L1 and B7-H3. Cell Death Discov. 2021, 7, 145. [Google Scholar] [CrossRef]
- Xu, W.; Atkins, M.B.; McDermott, D.F. Checkpoint inhibitor immunotherapy in kidney cancer. Nat. Rev. Urol. 2020, 17, 137–150. [Google Scholar] [CrossRef]
- Soleimani, M.; Thi, M.; Saxena, N.; Khalaf, D.J.; Eigl, B.J.; Chi, K.N.; Kollmannsberger, C.K.; Nappi, L. 693P Plasma exosome microRNA-155-3p expression in patients with metastatic renal cell carcinoma treated with immune checkpoint inhibitors: Potential biomarker of response to systemic therapy. Ann. Oncol. 2021, 32, S708. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, L.; Zou, L.; Yang, P.; Wu, R.; Mao, Y.; Zhou, H.; Li, R.; Wang, K.; Wang, W.; et al. MiR-20b, -21, and -130b inhibit PTEN expression resulting in B7-H1 over-expression in advanced colorectal cancer. Hum. Immunol. 2014, 75, 348–353. [Google Scholar] [CrossRef]
- Cortez, M.A.; Anfossi, S.; Ramapriyan, R.; Menon, H.; Atalar, S.C.; Aliru, M.; Welsh, J.; Calin, G.A. Role of miRNAs in immune responses and immunotherapy in cancer. Genes Chromosomes Cancer 2019, 58, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.-H.T.; Luo, Y.-H.; Li, A.-L.; Tsai, J.-C.; Wu, K.-L.; Chung, P.-J.; Ma, N. miRNA as a Modulator of Immunotherapy and Immune Response in Melanoma. Biomolecules 2021, 11, 1648. [Google Scholar] [CrossRef]
- Yi, M.; Xu, L.; Jiao, Y.; Luo, S.; Li, A.; Wu, K. The role of cancer-derived microRNAs in cancer immune escape. J. Hematol. Oncol. 2020, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Omar, H.A.; El-Serafi, A.T.; Hersi, F.; Arafa, E.A.; Zaher, D.M.; Madkour, M.; Arab, H.H.; Tolba, M.F. Immunomodulatory MicroRNAs in cancer: Targeting immune checkpoints and the tumor microenvironment. FEBS J. 2019, 286, 3540–3557. [Google Scholar] [CrossRef]
- Skafi, N.; Fayyad-Kazan, M.; Badran, B. Immunomodulatory role for MicroRNAs: Regulation of PD-1/PD-L1 and CTLA-4 immune checkpoints expression. Gene 2020, 754, 144888. [Google Scholar] [CrossRef]
- Eichmüller, S.B.; Osen, W.; Mandelboim, O.; Seliger, B. Immune Modulatory microRNAs Involved in Tumor Attack and Tumor Immune Escape. JNCI J. Natl. Cancer Inst. 2017, 109, djx034. [Google Scholar] [CrossRef]
- Seidel, J.A.; Otsuka, A.; Kabashima, K. Anti-PD-1 and Anti-CTLA-4 Therapies in Cancer: Mechanisms of Action, Efficacy, and Limitations. Front. Oncol. 2018, 8, 86. [Google Scholar] [CrossRef]
- Kooshkaki, O.; Derakhshani, A.; Hosseinkhani, N.; Torabi, M.; Safaei, S.; Brunetti, O.; Racanelli, V.; Silvestris, N.; Baradaran, B. Combination of Ipilimumab and Nivolumab in Cancers: From Clinical Practice to Ongoing Clinical Trials. Int. J. Mol. Sci. 2020, 21, 4427. [Google Scholar] [CrossRef]
- Acharya, N.; Sabatos-Peyton, C.; Anderson, A.C. Tim-3 finds its place in the cancer immunotherapy landscape. J. Immunother. Cancer 2020, 8, e000911. [Google Scholar] [CrossRef]
- Yuan, J.; Jiang, B.; Zhao, H.; Huang, Q. Prognostic implication of TIM-3 in clear cell renal cell carcinoma. Neoplasma 2014, 61, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Granier, C.; Dariane, C.; Combe, P.; Verkarre, V.; Urien, S.; Badoual, C.; Roussel, H.; Mandavit, M.; Ravel, P.; Sibony, M.; et al. Tim-3 Expression on Tumor-Infiltrating PD-1+CD8+ T Cells Correlates with Poor Clinical Outcome in Renal Cell Carcinoma. Cancer Res. 2017, 77, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, V.L.; Heusschen, R.; Caers, J.; Griffioen, A.W. Galectin expression in cancer diagnosis and prognosis: A systematic review. Biochim. Biophys. Acta-Rev. Cancer 2015, 1855, 235–247. [Google Scholar] [CrossRef]
- Yasinska, I.M.; Sakhnevych, S.S.; Pavlova, L.; Teo Hansen Selnø, A.; Teuscher Abeleira, A.M.; Benlaouer, O.; Gonçalves Silva, I.; Mosimann, M.; Varani, L.; Bardelli, M.; et al. The Tim-3-Galectin-9 Pathway and Its Regulatory Mechanisms in Human Breast Cancer. Front. Immunol. 2019, 10, 1594. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Sun, L.; Li, C.-F.; Wang, Y.-H.; Yao, J.; Li, H.; Yan, M.; Chang, W.-C.; Hsu, J.-M.; Cha, J.-H.; et al. Galectin-9 interacts with PD-1 and TIM-3 to regulate T cell death and is a target for cancer immunotherapy. Nat. Commun. 2021, 12, 832. [Google Scholar] [CrossRef] [PubMed]
- Jikuya, R.; Kishida, T.; Sakaguchi, M.; Yokose, T.; Yasui, M.; Hashizume, A.; Tatenuma, T.; Mizuno, N.; Muraoka, K.; Umemoto, S.; et al. Galectin-9 expression as a poor prognostic factor in patients with renal cell carcinoma. Cancer Immunol. Immunother. 2020, 69, 2041–2051. [Google Scholar] [CrossRef]
- Fu, H.; Liu, Y.; Xu, L.; Liu, W.; Fu, Q.; Liu, H.; Zhang, W.; Xu, J. Galectin-9 predicts postoperative recurrence and survival of patients with clear-cell renal cell carcinoma. Tumor Biol. 2015, 36, 5791–5799. [Google Scholar] [CrossRef]
- Kawashima, H.; Obayashi, A.; Kawamura, M.; Masaki, S.; Tamada, S.; Iguchi, T.; Uchida, J.; Kuratsukuri, K.; Tanaka, T.; Nakatani, T. Galectin 9 and PINCH, novel immunotherapy targets of renal cell carcinoma: A rationale to find potential tumour antigens and the resulting cytotoxic T lymphocytes induced by the derived peptides. BJU Int. 2014, 113, 320–332. [Google Scholar] [CrossRef]
- Dama, P.; Tang, M.; Fulton, N.; Kline, J.; Liu, H. Gal9/Tim-3 expression level is higher in AML patients who fail chemotherapy. J. Immunother. Cancer 2019, 7, 175. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, L.; Jing, D.; Xu, G.; Zhang, J.; Lin, L.; Zhao, J.; Yao, Z.; Lin, H. Galectin-9 Expression Predicts Favorable Clinical Outcome in Solid Tumors: A Systematic Review and Meta-Analysis. Front. Physiol. 2018, 9, 452. [Google Scholar] [CrossRef]
- Yang, R.; Sun, L.; Li, C.-F.; Wang, Y.-H.; Xia, W.; Liu, B.; Chu, Y.-Y.; Bover, L.; Vien, L.; Hung, M.-C. Development and characterization of anti-galectin-9 antibodies that protect T cells from galectin-9-induced cell death. J. Biol. Chem. 2022, 298, 101821. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Yuan, Q.; Xia, H.; Zhu, G.; Feng, Y.; Wang, Q.; Zhang, Z.; He, W.; Lu, J.; Dong, C.; et al. Analysis of VISTA expression and function in renal cell carcinoma highlights VISTA as a potential target for immunotherapy. Protein Cell 2019, 10, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Dong, J.; Zheng, Y.; Zhou, J.; Yuan, Y.; Ta, H.M.; Miller, H.E.; Olson, M.; Rajasekaran, K.; Ernstoff, M.S.; et al. Immune-Checkpoint Protein VISTA Regulates Antitumor Immunity by Controlling Myeloid Cell–Mediated Inflammation and Immunosuppression. Cancer Immunol. Res. 2019, 7, 1497–1510. [Google Scholar] [CrossRef]
- Kondo, Y.; Ohno, T.; Nishii, N.; Harada, K.; Yagita, H.; Azuma, M. Differential contribution of three immune checkpoint (VISTA, CTLA-4, PD-1) pathways to antitumor responses against squamous cell carcinoma. Oral Oncol. 2016, 57, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Liu, K.; Xiong, H. Roles of BTLA in Immunity and Immune Disorders. Front. Immunol. 2021, 12, 654960. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, J.; Tu, H.; Liang, D.; Chang, D.W.; Ye, Y.; Wu, X. Soluble immune checkpoint-related proteins as predictors of tumor recurrence, survival, and T cell phenotypes in clear cell renal cell carcinoma patients. J. Immunother. Cancer 2019, 7, 334. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Cao, X.; Li, Y.; Li, G.-Q.; He, Q.-H.; Li, S.-J.; Chen, J.; Xu, G.-L.; Zhang, K.-Q. High expression of herpes virus entry mediator is associated with poor prognosis in clear cell renal cell carcinoma. Am. J. Cancer Res. 2019, 9, 975–987. [Google Scholar]
- Hofmeyer, K.A.; Ray, A.; Zang, X. The contrasting role of B7-H3. Proc. Natl. Acad. Sci. USA 2008, 105, 10277–10278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ji, J.; Zhang, G.; Fang, C.; Jiang, F.; Ma, S.; Hou, J. Expression and significance of B7-H3 and Tie-2 in the tumor vasculature of clear cell renal carcinoma. OncoTargets Ther. 2017, 10, 5417–5424. [Google Scholar] [CrossRef]
- Xie, J.; Sun, M.; Zhang, D.; Chen, C.; Lin, S.; Zhang, G. Fibronectin enhances tumor metastasis through B7-H3 in clear cell renal cell carcinoma. FEBS Open Bio 2021, 11, 2977–2987. [Google Scholar] [CrossRef]
- Kontos, F.; Michelakos, T.; Kurokawa, T.; Sadagopan, A.; Schwab, J.H.; Ferrone, C.R.; Ferrone, S. B7-H3: An Attractive Target for Antibody-based Immunotherapy. Clin. Cancer Res. 2021, 27, 1227–1235. [Google Scholar] [CrossRef]
- Xu, Y.; Xiao, Y.; Luo, C.; Liu, Q.; Wei, A.; Yang, Y.; Zhao, L.; Wang, Y. Blocking PD-1/PD-L1 by an ADCC enhanced anti-B7-H3/PD-1 fusion protein engages immune activation and cytotoxicity. Int. Immunopharmacol. 2020, 84, 106584. [Google Scholar] [CrossRef] [PubMed]
- You, G.; Lee, Y.; Kang, Y.-W.; Park, H.W.; Park, K.; Kim, H.; Kim, Y.-M.; Kim, S.; Kim, J.-H.; Moon, D.; et al. B7-H3 × 4-1BB bispecific antibody augments antitumor immunity by enhancing terminally differentiated CD8+ tumor-infiltrating lymphocytes. Sci. Adv. 2021, 7, aax3160. [Google Scholar] [CrossRef]
- Fu, T.; He, Q.; Sharma, P. The ICOS/ICOSL Pathway Is Required for Optimal Antitumor Responses Mediated by Anti–CTLA-4 Therapy. Cancer Res. 2011, 71, 5445–5454. [Google Scholar] [CrossRef]
- Solinas, C.; Gu-Trantien, C.; Willard-Gallo, K. The rationale behind targeting the ICOS-ICOS ligand costimulatory pathway in cancer immunotherapy. ESMO Open 2020, 5, e000544. [Google Scholar] [CrossRef]
- Wang, M.; Du, Q.; Jin, J.; Wei, Y.; Lu, Y.; Li, Q. LAG3 and its emerging role in cancer immunotherapy. Clin. Transl. Med. 2021, 11, e365. [Google Scholar] [CrossRef]
- Zelba, H.; Bedke, J.; Hennenlotter, J.; Mostböck, S.; Zettl, M.; Zichner, T.; Chandran, A.; Stenzl, A.; Rammensee, H.-G.; Gouttefangeas, C. PD-1 and LAG-3 Dominate Checkpoint Receptor–Mediated T-cell Inhibition in Renal Cell Carcinoma. Cancer Immunol. Res. 2019, 7, 1891–1899. [Google Scholar] [CrossRef]
- Wang, J.; Sanmamed, M.F.; Datar, I.; Su, T.T.; Ji, L.; Sun, J.; Chen, L.; Chen, Y.; Zhu, G.; Yin, W.; et al. Fibrinogen-like Protein 1 Is a Major Immune Inhibitory Ligand of LAG-3. Cell 2019, 176, 334–347.e12. [Google Scholar] [CrossRef]
- Braun, D.A.; Bakouny, Z.; Hirsch, L.; Flippot, R.; Van Allen, E.M.; Wu, C.J.; Choueiri, T.K. Beyond conventional immune-checkpoint inhibition—Novel immunotherapies for renal cell carcinoma. Nat. Rev. Clin. Oncol. 2021, 18, 199–214. [Google Scholar] [CrossRef]
- Formisano, L.; Jansen, V.M.; Marciano, R.; Bianco, R. From Biology to Therapy: Improvements of Therapeutic Options in Lung Cancer. Anticancer Agents Med. Chem. 2019, 18, 1235–1240. [Google Scholar] [CrossRef]
- Bagegni, N.A.; Park, H.; Kraft, K.; O-Toole, M.; Gao, F.; Waqar, S.N.; Ratner, L.; Morgensztern, D.; Devarakonda, S.; Amin, M.; et al. Phase 1b trial of anti-VEGF/PDGFR vorolanib combined with immune checkpoint inhibitors in patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2022, 89, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-H.; Lee, H.J. The Frontline Immunotherapy-Based Treatment of Advanced Clear Cell Renal Cell Carcinoma: Current Evidence and Clinical Perspective. Biomedicines 2022, 10, 251. [Google Scholar] [CrossRef]
- Uchikawa, E.; Chen, Z.; Xiao, G.-Y.; Zhang, X.; Bai, X. Structural basis of the activation of c-MET receptor. Nat. Commun. 2021, 12, 4074. [Google Scholar] [CrossRef]
- Lindner, A.K.; Pichler, M.; Thurnher, M.; Pichler, R. Targeting c-Met to Improve Immune Checkpoint Inhibition in Metastatic Renal Cell Carcinoma. Eur. Urol. 2022, 81, 1–2. [Google Scholar] [CrossRef]
- Lai, Y.; Zeng, T.; Liang, X.; Wu, W.; Zhong, F.; Wu, W. Cell death-related molecules and biomarkers for renal cell carcinoma targeted therapy. Cancer Cell Int. 2019, 19, 221. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Wang, G.; Cui, X. Triple Combination Therapy with PD-1/PD-L1, BRAF, and MEK Inhibitor for Stage III–IV Melanoma: A Systematic Review and Meta-Analysis. Front. Oncol. 2021, 11, 2088. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Long, Q.; Guan, B.; Mu, L. Prognostic Value of High CXCR4 Expression in Renal Cell Carcinoma: A System Review and Meta-Analysis. Dis. Markers 2015, 2015, 568980. [Google Scholar] [CrossRef]
- Vaishampayan, U.N.; McDermott, D.F.; Matrana, M.R.; Rha, S.Y.; Zurita, A.J.; Ho, T.H.; Keam, B.; Lee, J.-L.; Joseph, R.W.; Ali, S.; et al. A phase 1/2 study evaluating the efficacy and safety of the oral CXCR4 inhibitor X4P-001 in combination with axitinib in patients with advanced renal cell carcinoma. J. Clin. Oncol. 2018, 36, 4510. [Google Scholar] [CrossRef]
- Cossu-Rocca, P.; Muroni, M.R.; Sanges, F.; Sotgiu, G.; Asunis, A.; Tanca, L.; Onnis, D.; Pira, G.; Manca, A.; Dore, S.; et al. EGFR kinase-dependent and kinase-independent roles in clear cell renal cell carcinoma. Am. J. Cancer Res. 2016, 6, 71–83. [Google Scholar]
- Li, L.; Deng, L.; Meng, X.; Gu, C.; Meng, L.; Li, K.; Zhang, X.; Meng, Y.; Xu, W.; Zhao, L.; et al. Tumor-targeting anti-EGFR x anti-PD1 bispecific antibody inhibits EGFR-overexpressing tumor growth by combining EGFR blockade and immune activation with direct tumor cell killing. Transl. Oncol. 2021, 14, 100916. [Google Scholar] [CrossRef]
- Matusz-Fisher, A.; Tan, A.R. Combination of HER2-targeted agents with immune checkpoint inhibitors in the treatment of HER2-positive breast cancer. Expert Opin. Biol. Ther. 2022, 22, 385–395. [Google Scholar] [CrossRef]
- Catenacci, D.V.T.; Kang, Y.-K.; Park, H.; Uronis, H.E.; Lee, K.-W.; Ng, M.C.H.; Enzinger, P.C.; Park, S.H.; Gold, P.J.; Lacy, J.; et al. Margetuximab plus pembrolizumab in patients with previously treated, HER2-positive gastro-oesophageal adenocarcinoma (CP-MGAH22–05): A single-arm, phase 1b–2 trial. Lancet Oncol. 2020, 21, 1066–1076. [Google Scholar] [CrossRef]
- Gu, C.; Zhu, H.; Deng, L.; Meng, X.; Li, K.; Xu, W.; Zhao, L.; Liu, Y.; Zhu, Z.; Huang, H. Bispecific antibody simultaneously targeting PD1 and HER2 inhibits tumor growth via direct tumor cell killing in combination with PD1/PDL1 blockade and HER2 inhibition. Acta Pharmacol. Sin. 2022, 43, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wang, B.; Zeng, J.; Sang, S.; Lei, J.; Lu, Y. MicroRNA-374b inhibits liver cancer progression via down regulating programmed cell death-1 expression on cytokine-induced killer cells. Oncol. Lett. 2018, 15, 4797–4804. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, N.; Li, Z.; Zhu, Q.; Li, F.; Yang, C.; Han, Q.; Lv, Y.; Zhou, Z.; Liu, Z. microRNA-4717 differentially interacts with its polymorphic target in the PD1 3′ untranslated region: A mechanism for regulating PD-1 expression and function in HBV-associated liver diseases. Oncotarget 2015, 6, 18933–18944. [Google Scholar] [CrossRef]
- Zhang, Q.; Di, W.; Dong, Y.; Lu, G.; Yu, J.; Li, J.; Li, P. High serum miR-183 level is associated with poor responsiveness of renal cancer to natural killer cells. Tumor Biol. 2015, 36, 9245–9249. [Google Scholar] [CrossRef]
- Grenda, A.; Krawczyk, P.; Błach, J.; Chmielewska, I.; Kubiatowski, T.; Kieszko, S.; Wojas-Krawczyk, K.; Kucharczyk, T.; Jarosz, B.; Paśnik, I.; et al. Tissue MicroRNA Expression as a Predictor of Response to Immunotherapy in NSCLC Patients. Front. Oncol. 2021, 10, 563613. [Google Scholar] [CrossRef]
- Song, N.; Li, P.; Song, P.; Li, Y.; Zhou, S.; Su, Q.; Li, X.; Yu, Y.; Li, P.; Feng, M.; et al. MicroRNA-138-5p Suppresses Non-small Cell Lung Cancer Cells by Targeting PD-L1/PD-1 to Regulate Tumor Microenvironment. Front. Cell Dev. Biol. 2020, 8, 540. [Google Scholar] [CrossRef]
- Jia, L.; Xi, Q.; Wang, H.; Zhang, Z.; Liu, H.; Cheng, Y.; Guo, X.; Zhang, J.; Zhang, Q.; Zhang, L.; et al. miR-142-5p regulates tumor cell PD-L1 expression and enhances anti-tumor immunity. Biochem. Biophys. Res. Commun. 2017, 488, 425–431. [Google Scholar] [CrossRef]
- Aichen, Z.; Kun, W.; Xiaochun, S.; Lingling, T. LncRNA FGD5-AS1 promotes the malignant phenotypes of ovarian cancer cells via targeting miR-142-5p. Apoptosis 2021, 26, 348–360. [Google Scholar] [CrossRef]
- Qu, F.; Ye, J.; Pan, X.; Wang, J.; Gan, S.; Chu, C.; Chu, J.; Zhang, X.; Liu, M.; He, H.; et al. MicroRNA-497-5p down-regulation increases PD-L1 expression in clear cell renal cell carcinoma. J. Drug Target. 2019, 27, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yu, H.; Yi, S.; Peng, X.; Su, P.; Xiao, Z.; Liu, R.; Tang, A.; Li, X.; Liu, F.; et al. The tumor suppressor miR-138-5p targets PD-L1 in colorectal cancer. Oncotarget 2016, 7, 45370–45384. [Google Scholar] [CrossRef] [PubMed]
- Ashizawa, M.; Okayama, H.; Ishigame, T.; Thar Min, A.K.; Saito, K.; Ujiie, D.; Murakami, Y.; Kikuchi, T.; Nakayama, Y.; Noda, M.; et al. miRNA-148a-3p Regulates Immunosuppression in DNA Mismatch Repair–Deficient Colorectal Cancer by Targeting PD-L1. Mol. Cancer Res. 2019, 17, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Y.; Zhang, J.; Hou, L.-D.; Zhang, R.; Chen, W.; Fan, H.-N.; Huang, Y.-X.; Liu, H.; Zhu, J.-S. Upregulation of PD-L1 predicts poor prognosis and is associated with miR-191-5p dysregulation in colon adenocarcinoma. Int. J. Immunopathol. Pharmacol. 2018, 32, 205873841879031. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Guo, Q.; Li, X.; Yang, X.; Ni, H.; Wang, T.; Zhao, Q.; Liu, H.; Xing, Y.; Xi, T.; et al. MiR-873/PD-L1 axis regulates the stemness of breast cancer cells. EBioMedicine 2019, 41, 395–407. [Google Scholar] [CrossRef]
- Yang, L.; Cai, Y.; Zhang, D.; Sun, J.; Xu, C.; Zhao, W.; Jiang, W.; Pan, C. miR-195/miR-497 Regulate CD274 Expression of Immune Regulatory Ligands in Triple-Negative Breast Cancer. J. Breast Cancer 2018, 21, 371. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, X.; Yang, M.; Kan, Q.; Duan, Z. miR3609 sensitizes breast cancer cells to adriamycin by blocking the programmed death-ligand 1 immune checkpoint. Exp. Cell Res. 2019, 380, 20–28. [Google Scholar] [CrossRef]
- Dastmalchi, N.; Hosseinpourfeizi, M.A.; Khojasteh, S.M.B.; Baradaran, B.; Safaralizadeh, R. Tumor suppressive activity of miR-424-5p in breast cancer cells through targeting PD-L1 and modulating PTEN/PI3K/AKT/mTOR signaling pathway. Life Sci. 2020, 259, 118239. [Google Scholar] [CrossRef]
- Audrito, V.; Serra, S.; Stingi, A.; Orso, F.; Gaudino, F.; Bologna, C.; Neri, F.; Garaffo, G.; Nassini, R.; Baroni, G.; et al. PD-L1 up-regulation in melanoma increases disease aggressiveness and is mediated through miR-17-5p. Oncotarget 2017, 8, 15894–15911. [Google Scholar] [CrossRef]
- Mastroianni, J.; Stickel, N.; Andrlova, H.; Hanke, K.; Melchinger, W.; Duquesne, S.; Schmidt, D.; Falk, M.; Andrieux, G.; Pfeifer, D.; et al. miR-146a Controls Immune Response in the Melanoma Microenvironment. Cancer Res. 2019, 79, 183–195. [Google Scholar] [CrossRef]
- Kao, S.C.; Cheng, Y.Y.; Williams, M.; Kirschner, M.B.; Madore, J.; Lum, T.; Sarun, K.H.; Linton, A.; McCaughan, B.; Klebe, S.; et al. Tumor Suppressor microRNAs Contribute to the Regulation of PD-L1 Expression in Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2017, 12, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Indovina, P.; Mattioli, E.; Forte, I.M.; Iannuzzi, C.A.; Luzzi, L.; Bellan, C.; De Summa, S.; Bucci, E.; Di Marzo, D.; et al. P53-regulated miR-320a targets PDL1 and is downregulated in malignant mesothelioma. Cell Death Dis. 2020, 11, 748. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Yan, F.; Wu, C. Overexpressed miR-195 attenuated immune escape of diffuse large B-cell lymphoma by targeting PD-L1. Biomed. Pharmacother. 2018, 98, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Sun, R.; Zhao, H.-J.; Fu, D.; Zhong, H.-J.; Weng, X.-Q.; Qu, B.; Zhao, Y.; Wang, L.; Zhao, W.-L. MiR155 sensitized B-lymphoma cells to anti-PD-L1 antibody via PD-1/PD-L1-mediated lymphoma cell interaction with CD8+T cells. Mol. Cancer 2019, 18, 54. [Google Scholar] [CrossRef]
- Sun, J.-R.; Zhang, X.; Zhang, Y. MiR-214 prevents the progression of diffuse large B-cell lymphoma by targeting PD-L1. Cell. Mol. Biol. Lett. 2019, 24, 68. [Google Scholar] [CrossRef]
- Tao, Z.; Xu, S.; Ruan, H.; Wang, T.; Song, W.; Qian, L.; Chen, K. MiR-195/-16 Family Enhances Radiotherapy via T Cell Activation in the Tumor Microenvironment by Blocking the PD-L1 Immune Checkpoint. Cell. Physiol. Biochem. 2018, 48, 801–814. [Google Scholar] [CrossRef]
- Cortez, M.A.; Ivan, C.; Valdecanas, D.; Wang, X.; Peltier, H.J.; Ye, Y.; Araujo, L.; Carbone, D.P.; Shilo, K.; Giri, D.K.; et al. PDL1 Regulation by p53 via miR-34. JNCI J. Natl. Cancer Inst. 2016, 108, djv303. [Google Scholar] [CrossRef]
- Xie, W.-B.; Liang, L.-H.; Wu, K.-G.; Wang, L.-X.; He, X.; Song, C.; Wang, Y.-Q.; Li, Y.-H. MiR-140 Expression Regulates Cell Proliferation and Targets PD-L1 in NSCLC. Cell. Physiol. Biochem. 2018, 46, 654–663. [Google Scholar] [CrossRef]
- Tang, D.; Zhao, D.; Wu, Y.; Yao, R.; Zhou, L.; Lu, L.; Gao, W.; Sun, Y. The miR-3127-5p/p-STAT3 axis up-regulates PD-L1 inducing chemoresistance in non-small-cell lung cancer. J. Cell. Mol. Med. 2018, 22, 3847–3856. [Google Scholar] [CrossRef]
- Chen, L.; Gibbons, D.L.; Goswami, S.; Cortez, M.A.; Ahn, Y.-H.; Byers, L.A.; Zhang, X.; Yi, X.; Dwyer, D.; Lin, W.; et al. Metastasis is regulated via microRNA-200/ZEB1 axis control of tumour cell PD-L1 expression and intratumoral immunosuppression. Nat. Commun. 2014, 5, 5241. [Google Scholar] [CrossRef]
- Wei, S.; Wang, K.; Huang, X.; Zhao, Z.; Zhao, Z. LncRNA MALAT1 contributes to non-small cell lung cancer progression via modulating miR-200a-3p/programmed death-ligand 1 axis. Int. J. Immunopathol. Pharmacol. 2019, 33, 205873841985969. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, J.; Dong, K.; Lin, F.; Long, M.; Ouyang, Y.; Wei, J.; Chen, X.; Weng, Y.; He, T.; et al. Tumor suppressor miR-34a targets PD-L1 and functions as a potential immunotherapeutic target in acute myeloid leukemia. Cell. Signal. 2015, 27, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fan, L.; Yu, H.; Zhang, J.; He, Y.; Feng, D.; Wang, F.; Li, X.; Liu, Q.; Li, Y.; et al. Endoplasmic Reticulum Stress Causes Liver Cancer Cells to Release Exosomal miR-23a-3p and Up-regulate Programmed Death Ligand 1 Expression in Macrophages. Hepatology 2019, 70, hep.30607. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Tan, W.; Liu, S.; Huang, X.; Lin, J.; Liang, R.; Su, L.; Su, Q.; Wang, C. MiR-570 inhibited the cell proliferation and invasion through directly targeting B7-H1 in hepatocellular carcinoma. Tumor Biol. 2015, 36, 9049–9057. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, Y.; Sun, Y.; Yan, X.; Wang, P. miR-375 inhibits IFN-γ-induced programmed death 1 ligand�1 surface expression in head and neck squamous cell carcinoma cells by blocking JAK2/STAT1 signaling. Oncol. Rep. 2018, 39, 1461–1468. [Google Scholar] [CrossRef]
- Sheng, Q.; Zhang, Y.; Wang, Z.; Ding, J.; Song, Y.; Zhao, W. Cisplatin-mediated down-regulation of miR-145 contributes to up-regulation of PD-L1 via the c-Myc transcription factor in cisplatin-resistant ovarian carcinoma cells. Clin. Exp. Immunol. 2020, 200, 45–52. [Google Scholar] [CrossRef]
- Zhu, J.; Li, Y.; Luo, Y.; Xu, J.; Liufu, H.; Tian, Z.; Huang, C.; Li, J.; Huang, C. A Feedback Loop Formed by ATG7/Autophagy, FOXO3a/miR-145 and PD-L1 Regulates Stem-Like Properties and Invasion in Human Bladder Cancer. Cancers 2019, 11, 349. [Google Scholar] [CrossRef]
- Wu, L.; Chen, Z.; Zhang, J.; Xing, Y. Effect of miR-513a-5p on etoposide-stimulating B7-H1 expression in retinoblastoma cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2012, 32, 601–606. [Google Scholar] [CrossRef]
- Miliotis, C.; Slack, F.J. miR-105-5p regulates PD-L1 expression and tumor immunogenicity in gastric cancer. Cancer Lett. 2021, 518, 115–126. [Google Scholar] [CrossRef]
- Wang, W.; Li, F.; Mao, Y.; Zhou, H.; Sun, J.; Li, R.; Liu, C.; Chen, W.; Hua, D.; Zhang, X. A miR-570 binding site polymorphism in the B7-H1 gene is associated with the risk of gastric adenocarcinoma. Hum. Genet. 2013, 132, 641–648. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, D.; Xie, G.; Yin, Y.; Zhao, E.; Tao, K.; Li, R. MicroRNA-152 regulates immune response via targeting B7-H1 in gastric carcinoma. Oncotarget 2017, 8, 28125–28134. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Li, W.; Li, R.; Wu, K.; Zhao, E.; Zhang, Y.; Zhang, P.; Shi, L.; Wang, D.; Yin, Y.; et al. Helicobacter Pylori Promote B7-H1 Expression by Suppressing miR-152 and miR-200b in Gastric Cancer Cells. PLoS ONE 2017, 12, e0168822. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Liu, F.; Chu, Y.; Zhai, Q.; Wei, X.; Shao, J.; Li, R.; Xu, Q.; Yu, L.; Liu, B.; et al. MicroRNA-200c Nanoparticles Sensitized Gastric Cancer Cells to Radiotherapy by Regulating PD-L1 Expression and EMT. Cancer Manag. Res. 2020, 12, 12215–12223. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.; Xiong, Y.; Yu, J.; Chen, L.; Tao, T.; Yi, S.; Hanley, S.J.B.; Yue, J.; Watari, H.; Sakuragi, N. Control of PD-L1 expression by miR-140/142/340/383 and oncogenic activation of the OCT4–miR-18a pathway in cervical cancer. Oncogene 2018, 37, 5257–5268. [Google Scholar] [CrossRef]
- Miao, S.; Mao, X.; Zhao, S.; Song, K.; Xiang, C.; Lv, Y.; Jiang, H.; Wang, L.; Li, B.; Yang, X.; et al. miR-217 inhibits laryngeal cancer metastasis by repressing AEG-1 and PD-L1 expression. Oncotarget 2017, 8, 62143–62153. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Zou, H. microRNA-20b-5p overexpression combing Pembrolizumab potentiates cancer cells to radiation therapy via repressing programmed death-ligand 1. Bioengineered 2022, 13, 917–929. [Google Scholar] [CrossRef]
- Wang, C.; Li, X.; Zhang, L.; Chen, Y.; Dong, R.; Zhang, J.; Zhao, J.; Guo, X.; Yang, G.; Li, Y.; et al. miR-194-5p down-regulates tumor cell PD-L1 expression and promotes anti-tumor immunity in pancreatic cancer. Int. Immunopharmacol. 2021, 97, 107822. [Google Scholar] [CrossRef]
- Xu, S.; Tao, Z.; Hai, B.; Liang, H.; Shi, Y.; Wang, T.; Song, W.; Chen, Y.; OuYang, J.; Chen, J.; et al. miR-424(322) reverses chemoresistance via T-cell immune response activation by blocking the PD-L1 immune checkpoint. Nat. Commun. 2016, 7, 11406. [Google Scholar] [CrossRef]
- Wei, J.; Nduom, E.K.; Kong, L.-Y.; Hashimoto, Y.; Xu, S.; Gabrusiewicz, K.; Ling, X.; Huang, N.; Qiao, W.; Zhou, S.; et al. MiR-138 exerts anti-glioma efficacy by targeting immune checkpoints. Neuro. Oncol. 2016, 18, 639–648. [Google Scholar] [CrossRef]
- Zhao, X.; Yuan, C.; Wangmo, D.; Subramanian, S. Tumor-Secreted Extracellular Vesicles Regulate T-Cell Costimulation and Can Be Manipulated To Induce Tumor-Specific T-Cell Responses. Gastroenterology 2021, 161, 560–574.e11. [Google Scholar] [CrossRef]
- Boldrini, L.; Giordano, M.; Niccoli, C.; Melfi, F.; Lucchi, M.; Mussi, A.; Fontanini, G. Role of microRNA-33a in regulating the expression of PD-1 in lung adenocarcinoma. Cancer Cell Int. 2017, 17, 105. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Johnston, N.; Zheng, X.; Wang, H.; Zhang, X.; Gao, D.; Min, W. miR-28 modulates exhaustive differentiation of T cells through silencing programmed cell death-1 and regulating cytokine secretion. Oncotarget 2016, 7, 53735–53750. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.-R.; Wang, L.-M.; Shen, J.-J. Overexpression of miR-32 inhibits the proliferation and metastasis of ovarian cancer cells by targeting BTLA. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4671–4678. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, Y.; Andalib, A.; Mohammad-Ganji, M.; Homayouni, V.; Sharifi, M.; Ganjalikhani-Hakemi, M. Evaluation of the effect of TIM-3 suppression by miR-498 and its effect on apoptosis and proliferation rate of HL-60 cell line. Pathol. Res. Pract. 2018, 214, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Lou, Q.; Liu, R.; Yang, X.; Li, W.; Huang, L.; Wei, L.; Tan, H.; Xiang, N.; Chan, K.; Chen, J.; et al. miR-448 targets IDO1 and regulates CD8+ T cell response in human colon cancer. J. Immunother. Cancer 2019, 7, 210. [Google Scholar] [CrossRef]
- Huang, Q.; Xia, J.; Wang, L.; Wang, X.; Ma, X.; Deng, Q.; Lu, Y.; Kumar, M.; Zhou, Z.; Li, L.; et al. miR-153 suppresses IDO1 expression and enhances CAR T cell immunotherapy. J. Hematol. Oncol. 2018, 11, 58. [Google Scholar] [CrossRef]
- Bieg, D.; Sypniewski, D.; Nowak, E.; Bednarek, I. MiR-424-3p suppresses galectin-3 expression and sensitizes ovarian cancer cells to cisplatin. Arch. Gynecol. Obstet. 2019, 299, 1077–1087. [Google Scholar] [CrossRef]
- Lu, W.; Wang, J.; Yang, G.; Yu, N.; Huang, Z.; Xu, H.; Li, J.; Qiu, J.; Zeng, X.; Chen, S.; et al. Posttranscriptional regulation of Galectin-3 by miR-128 contributes to colorectal cancer progression. Oncotarget 2017, 8, 15242–15251. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, W.; Zhuang, C.; Geng, Z.; Hou, C.; Huang, D.; Hu, L.; Wang, X. microRNA-22 downregulation of galectin-9 influences lymphocyte apoptosis and tumor cell proliferation in liver cancer. Oncol. Rep. 2015, 34, 1771–1778. [Google Scholar] [CrossRef]
- Yang, Q.; Hou, C.; Huang, D.; Zhuang, C.; Jiang, W.; Geng, Z.; Wang, X.; Hu, L. miR-455-5p functions as a potential oncogene by targeting galectin-9 in colon cancer. Oncol. Lett. 2017, 13, 1958–1964. [Google Scholar] [CrossRef]
- Morishita, A.; Nomura, K.; Tani, J.; Fujita, K.; Iwama, H.; Takuma, K.; Nakahara, M.; Tadokoro, T.; Oura, K.; Chiyo, T.; et al. Galectin-9 suppresses the tumor growth of colon cancer in vitro and in vivo. Oncol. Rep. 2021, 45, 105. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Tang, R.; Zhu, J.; Zou, L.; Wu, R.; Zhou, H.; Mao, Y.; Li, R.; Hua, D.; Wang, W.; et al. A functional variant at miR-24 binding site in B7-H2 alters susceptibility to gastric cancer in a Chinese Han population. Mol. Immunol. 2013, 56, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Cheung, I.Y.; Guo, H.-F.; Cheung, N.-K.V. MicroRNA miR-29 Modulates Expression of Immunoinhibitory Molecule B7-H3: Potential Implications for Immune Based Therapy of Human Solid Tumors. Cancer Res. 2009, 69, 6275–6281. [Google Scholar] [CrossRef] [PubMed]
- Huang, L. The expression and clinical significance of B7-H3 and miR-145 in lung cancer patients with malignant pleural effusion. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6759–6766. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.; Xu, M.; Zhou, F.; Yan, J. Serum miR-1301-3p, miR-335-5p, miR-28-5p, and their target B7-H3 may serve as novel biomarkers for colorectal cancer. J. BUON 2019, 24, 1120–1127. [Google Scholar]
- Zhou, X.; Mao, Y.; Zhu, J.; Meng, F.; Chen, Q.; Tao, L.; Li, R.; Fu, F.; Liu, C.; Hu, Y.; et al. TGF-β1 promotes colorectal cancer immune escape by elevating B7-H3 and B7-H4 via the miR-155/miR-143 axis. Oncotarget 2016, 7, 67196–67211. [Google Scholar] [CrossRef]
- Hu, X.; Xu, M.; Hu, Y.; Li, N.; Zhou, L. B7-H3, Negatively Regulated by miR-128, Promotes Colorectal Cancer Cell Proliferation and Migration. Cell Biochem. Biophys. 2021, 79, 397–405. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Xie, C.; Sun, M.; Hu, C.; Zhang, Z.; Luan, L.; Zhou, J.; Zhou, J.; Zhu, X.; et al. MicroRNA miR-29a Inhibits Colon Cancer Progression by Downregulating B7-H3 Expression: Potential Molecular Targets for Colon Cancer Therapy. Mol. Biotechnol. 2021, 63, 849–861. [Google Scholar] [CrossRef]
- Wang, Z.-S.; Zhong, M.; Bian, Y.-H.; Mu, Y.-F.; Qin, S.-L.; Yu, M.-H.; Qin, J. MicroRNA-187 inhibits tumor growth and invasion by directly targeting CD276 in colorectal cancer. Oncotarget 2016, 7, 44266–44276. [Google Scholar] [CrossRef]
- Zhao, J.; Lei, T.; Xu, C.; Li, H.; Ma, W.; Yang, Y.; Fan, S.; Liu, Y. MicroRNA-187, down-regulated in clear cell renal cell carcinoma and associated with lower survival, inhibits cell growth and migration though targeting B7-H3. Biochem. Biophys. Res. Commun. 2013, 438, 439–444. [Google Scholar] [CrossRef]
- Wang, J.; Chong, K.K.; Nakamura, Y.; Nguyen, L.; Huang, S.K.; Kuo, C.; Zhang, W.; Yu, H.; Morton, D.L.; Hoon, D.S.B. B7-H3 Associated with Tumor Progression and Epigenetic Regulatory Activity in Cutaneous Melanoma. J. Investig. Dermatol. 2013, 133, 2050–2058. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Mou, J.; Cui, L.; Wang, X.; Zhang, Z. Astragaloside IV inhibits cell proliferation of colorectal cancer cell lines through down-regulation of B7-H3. Biomed. Pharmacother. 2018, 102, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Nygren, M.K.; Tekle, C.; Ingebrigtsen, V.A.; Mäkelä, R.; Krohn, M.; Aure, M.R.; Nunes-Xavier, C.E.; Perälä, M.; Tramm, T.; Alsner, J.; et al. Identifying microRNAs regulating B7-H3 in breast cancer: The clinical impact of microRNA-29c. Br. J. Cancer 2014, 110, 2072–2080. [Google Scholar] [CrossRef] [PubMed]
- Li, R.G.; Gao, Z.; Jiang, Y. B7-H3 repression by miR-539 suppresses cell proliferation in human gliomas. Int. J. Clin. Exp. Pathol. 2017, 10, 4363–4369. [Google Scholar]
- Wang, L.; Kang, F.; Sun, N.; Wang, J.; Chen, W.; Li, D.; Shan, B. The tumor suppressor miR-124 inhibits cell proliferation and invasion by targeting B7-H3 in osteosarcoma. Tumor Biol. 2016, 37, 14939–14947. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, J.; Zhu, M.; Wang, Y.; Yang, S.; Ke, X. MicroRNA-506 inhibits the proliferation and invasion of mantle cell lymphoma cells by targeting B7 H3. Biochem. Biophys. Res. Commun. 2019, 508, 1067–1073. [Google Scholar] [CrossRef]
- Gao, Y.; Fang, P.; Li, W.-J.; Zhang, J.; Wang, G.-P.; Jiang, D.-F.; Chen, F.-P. LncRNA NEAT1 sponges miR-214 to regulate M2 macrophage polarization by regulation of B7-H3 in multiple myeloma. Mol. Immunol. 2020, 117, 20–28. [Google Scholar] [CrossRef]
- Purvis, I.J.; Avilala, J.; Guda, M.R.; Venkataraman, S.; Vibhakar, R.; Tsung, A.J.; Velpula, K.K.; Asuthkar, S. Role of MYC-miR-29-B7-H3 in Medulloblastoma Growth and Angiogenesis. J. Clin. Med. 2019, 8, 1158. [Google Scholar] [CrossRef]
- Kanchan, R.K.; Perumal, N.; Atri, P.; Chirravuri Venkata, R.; Thapa, I.; Klinkebiel, D.L.; Donson, A.M.; Perry, D.; Punsoni, M.; Talmon, G.A.; et al. MiR-1253 exerts tumor-suppressive effects in medulloblastoma via inhibition of CDK6 and CD276 (B7-H3). Brain Pathol. 2020, 30, 732–745. [Google Scholar] [CrossRef]
- Yang, X.; Feng, K.-X.; Li, H.; Wang, L.; Xia, H. MicroRNA-199a Inhibits Cell Proliferation, Migration, and Invasion and Activates AKT/mTOR Signaling Pathway by Targeting B7-H3 in Cervical Cancer. Technol. Cancer Res. Treat. 2020, 19, 153303382094224. [Google Scholar] [CrossRef]
- Oliveira, P.; Carvalho, J.; Rocha, S.; Azevedo, M.; Reis, I.; Camilo, V.; Sousa, B.; Valente, S.; Paredes, J.; Almeida, R.; et al. Dies1/VISTA expression loss is a recurrent event in gastric cancer due to epigenetic regulation. Sci. Rep. 2016, 6, 34860. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Tang, R.; Qi, Q.; Zhou, X.; Zhou, H.; Mao, Y.; Li, R.; Liu, C.; Wang, W.; Hua, D.; et al. Five functional polymorphisms of B7/CD28 co-signaling molecules alter susceptibility to colorectal cancer. Cell. Immunol. 2015, 293, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Feng, L.; Wu, W.; Weng, T.; Hu, C.; Hong, B.; Wang, F.X.C.; Shen, L.; Wang, Q.; Jin, X.; et al. MicroRNA Expression Profiling of Pancreatic Cancer Cell Line L3.6p1 Following B7-H4 Knockdown. Cell. Physiol. Biochem. 2017, 44, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Ran, Z.; Liu, M.; Ou, Y. Prognostic Significance of Potential Immune Checkpoint Member HHLA2 in Human Tumors: A Comprehensive Analysis. Front. Immunol. 2019, 10, 1573. [Google Scholar] [CrossRef]
- Korotaeva, A.; Mansorunov, D.; Apanovich, N.; Kuzevanova, A.; Karpukhin, A. MiRNA Expression in Neuroendocrine Neoplasms of Frequent Localizations. Non-Coding RNA 2021, 7, 38. [Google Scholar] [CrossRef]
- Kipkeeva, F.; Muzaffarova, T.; Korotaeva, A.; Nikulin, M.; Grishina, K.; Mansorunov, D.; Apanovich, P.; Karpukhin, A. MicroRNA in Gastric Cancer Development: Mechanisms and Biomarkers. Diagnostics 2020, 10, 891. [Google Scholar] [CrossRef]
- Reid, G.; Pel, M.E.; Kirschner, M.B.; Cheng, Y.Y.; Mugridge, N.; Weiss, J.; Williams, M.; Wright, C.; Edelman, J.J.B.; Vallely, M.P.; et al. Restoring expression of miR-16: A novel approach to therapy for malignant pleural mesothelioma. Ann. Oncol. 2013, 24, 3128–3135. [Google Scholar] [CrossRef]
- Yeh, M.; Wang, Y.-Y.; Yoo, J.Y.; Oh, C.; Otani, Y.; Kang, J.M.; Park, E.S.; Kim, E.; Chung, S.; Jeon, Y.-J.; et al. MicroRNA-138 suppresses glioblastoma proliferation through downregulation of CD44. Sci. Rep. 2021, 11, 9219. [Google Scholar] [CrossRef]
- He, Z.; Ruan, X.; Liu, X.; Zheng, J.; Liu, Y.; Liu, L.; Ma, J.; Shao, L.; Wang, D.; Shen, S.; et al. FUS/circ_002136/miR-138-5p/SOX13 feedback loop regulates angiogenesis in Glioma. J. Exp. Clin. Cancer Res. 2019, 38, 65. [Google Scholar] [CrossRef]
- Kang, X.; Kong, B.; Chen, Q.; Zhao, S. Low expression of miR-138 inhibit the proliferation, migration and invasion of colorectal cancer and affect patient survival by targeting SIRT1. Transl. Cancer Res. 2021, 10, 3548–3559. [Google Scholar] [CrossRef]
- Kasinski, A.L.; Slack, F.J. miRNA-34 Prevents Cancer Initiation and Progression in a Therapeutically Resistant K-ras and p53-Induced Mouse Model of Lung Adenocarcinoma. Cancer Res. 2012, 72, 5576–5587. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-L.; Liu, X.-M.; Zhang, C.-Y.; Zhou, J.-B.; Shao, Y.; Liang, C.; Wang, H.-M.; Hua, Z.-Y.; Lu, S.-D.; Ma, Z.-L. MicroRNA-34a/EGFR axis plays pivotal roles in lung tumorigenesis. Oncogenesis 2017, 6, e372. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Huang, C.; Li, S.; Yang, C.; Xi, Y.; Wang, L.; Zhang, F.; Fu, Y.; Li, D. Hsa-miR-326 targets CCND1 and inhibits non-small cell lung cancer development. Oncotarget 2016, 7, 8341–8359. [Google Scholar] [CrossRef]
- Cai, M.; Wang, Z.; Zhang, J.; Zhou, H.; Jin, L.; Bai, R.; Weng, Y. Adam17, a Target of Mir-326, Promotes Emt-Induced Cells Invasion in Lung Adenocarcinoma. Cell. Physiol. Biochem. 2015, 36, 1175–1185. [Google Scholar] [CrossRef]
- Wu, Z.; Wu, Q.; Wang, C.; Wang, X.; Huang, J.; Zhao, J.; Mao, S.; Zhang, G.; Xu, X.; Zhang, N. miR-340 inhibition of breast cancer cell migration and invasion through targeting of oncoprotein c-Met. Cancer 2011, 117, 2842–2852. [Google Scholar] [CrossRef]
- Bruning, U.; Cerone, L.; Neufeld, Z.; Fitzpatrick, S.F.; Cheong, A.; Scholz, C.C.; Simpson, D.A.; Leonard, M.O.; Tambuwala, M.M.; Cummins, E.P.; et al. MicroRNA-155 Promotes Resolution of Hypoxia-Inducible Factor 1α Activity during Prolonged Hypoxia. Mol. Cell. Biol. 2011, 31, 4087–4096. [Google Scholar] [CrossRef]
- Su, J.; Liang, H.; Yao, W.; Wang, N.; Zhang, S.; Yan, X.; Feng, H.; Pang, W.; Wang, Y.; Wang, X.; et al. MiR-143 and MiR-145 Regulate IGF1R to Suppress Cell Proliferation in Colorectal Cancer. PLoS ONE 2014, 9, e114420. [Google Scholar] [CrossRef]
- Qian, X.; Yu, J.; Yin, Y.; He, J.; Wang, L.; Li, Q.; Zhang, L.-Q.; Li, C.-Y.; Shi, Z.-M.; Xu, Q.; et al. MicroRNA-143 inhibits tumor growth and angiogenesis and sensitizes chemosensitivity to oxaliplatin in colorectal cancers. Cell Cycle 2013, 12, 1385–1394. [Google Scholar] [CrossRef]
- Yin, Y.; Yan, Z.-P.; Lu, N.-N.; Xu, Q.; He, J.; Qian, X.; Yu, J.; Guan, X.; Jiang, B.-H.; Liu, L.-Z. Downregulation of miR-145 associated with cancer progression and VEGF transcriptional activation by targeting N-RAS and IRS1. Biochim. Biophys. Acta-Gene Regul. Mech. 2013, 1829, 239–247. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, Z.; Liu, Z.; Qiu, B.; Liu, K.; Dong, G. MicroRNA-335 inhibits invasion and metastasis of colorectal cancer by targeting ZEB2. Med. Oncol. 2014, 31, 982. [Google Scholar] [CrossRef]
- Lian, B.; Yang, D.; Liu, Y.; Shi, G.; Li, J.; Yan, X.; Jin, K.; Liu, X.; Zhao, J.; Shang, W.; et al. miR-128 Targets the SIRT1/ROS/DR5 Pathway to Sensitize Colorectal Cancer to TRAIL-Induced Apoptosis. Cell. Physiol. Biochem. 2018, 49, 2151–2162. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.I.; Nicoloso, M.S.; Zeng, L.; Ivan, C.; Spizzo, R.; Gafà, R.; Xiao, L.; Zhang, X.; Vannini, I.; Fanini, F.; et al. Strand-Specific miR-28-5p and miR-28-3p Have Distinct Effects in Colorectal Cancer Cells. Gastroenterology 2012, 142, 886–896.e9. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zeng, Y.; Zhou, J.M.; Nie, S.L.; Peng, Q.; Gong, J.; Huo, J.R. MicroRNA-1246 promotes growth and metastasis of colorectal cancer cells involving CCNG2 reduction. Mol. Med. Rep. 2016, 13, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.; Miao, Z.; Chen, Y.; Huang, C.; Yeh, Y.; Yang, I.; Wang, J. miR-148a inhibits early relapsed colorectal cancers and the secretion of VEGF by indirectly targeting HIF-1α under non-hypoxia/hypoxia conditions. J. Cell. Mol. Med. 2019, 23, 3572–3582. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Huang, Q.; Ren, X.; Hu, H.; Sheng, H.; Lai, M. MiR-148a promotes apoptosis by targeting Bcl-2 in colorectal cancer. Cell Death Differ. 2011, 18, 1702–1710. [Google Scholar] [CrossRef]
- Zhao, W.; Zheng, J.; Wei, G.; Yang, K.; Wang, G.; Sun, X. miR-148a inhibits cell proliferation and migration through targeting ErbB3 in colorectal cancer. Oncol. Lett. 2019, 18, 2530–2536. [Google Scholar] [CrossRef]
- Shi, L.; Xi, J.; Xu, X.; Peng, B.; Zhang, B. MiR-148a suppressed cell invasion and migration via targeting WNT10b and modulating β-catenin signaling in cisplatin-resistant colorectal cancer cells. Biomed. Pharmacother. 2019, 109, 902–909. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, T.; Gao, S.; Cheng, M.; Shao, Y.; Xi, Y.; Guo, L.; Zhang, D.; Gao, W.; Zhang, G.; et al. miR-148a-3p silences the CANX/MHC-I pathway and impairs CD8+ T cell-mediated immune attack in colorectal cancer. FASEB J. 2021, 35, e2177. [Google Scholar] [CrossRef]
- Rodriguez-Barrueco, R.; Nekritz, E.A.; Bertucci, F.; Yu, J.; Sanchez-Garcia, F.; Zeleke, T.Z.; Gorbatenko, A.; Birnbaum, D.; Ezhkova, E.; Cordon-Cardo, C.; et al. miR-424(322)/503 is a breast cancer tumor suppressor whose loss promotes resistance to chemotherapy. Genes Dev. 2017, 31, 553–566. [Google Scholar] [CrossRef]
- Wang, F.; Li, L.; Chen, Z.; Zhu, M.; Gu, Y. MicroRNA-214 acts as a potential oncogene in breast cancer by targeting the PTEN-PI3K/Akt signaling pathway. Int. J. Mol. Med. 2016, 37, 1421–1428. [Google Scholar] [CrossRef]
- Lu, Y.-Y.; Zheng, J.-Y.; Liu, J.; Huang, C.-L.; Zhang, W.; Zeng, Y. miR-183 induces cell proliferation, migration, and invasion by regulating PDCD4 expression in the SW1990 pancreatic cancer cell line. Biomed. Pharmacother. 2015, 70, 151–157. [Google Scholar] [CrossRef]
- Li, X.; Chen, W.; Jin, Y.; Xue, R.; Su, J.; Mu, Z.; Li, J.; Jiang, S. miR-142-5p enhances cisplatin-induced apoptosis in ovarian cancer cells by targeting multiple anti-apoptotic genes. Biochem. Pharmacol. 2019, 161, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, G.; Luo, X.; Wang, D.; Zhou, W.; Zhang, Y.; Zhang, W.; Chen, J.; Meng, Q.; Chen, E.; et al. Co-delivery of 5-fluorouracil and miRNA-34a mimics by host-guest self-assembly nanocarriers for efficacious targeted therapy in colorectal cancer patient-derived tumor xenografts. Theranostics 2021, 11, 2475–2489. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, X.; Lv, Y.; Xin, X.; Qin, C.; Han, X.; Yang, L.; He, W.; Yin, L. Cytosolic co-delivery of miRNA-34a and docetaxel with core-shell nanocarriers via caveolae-mediated pathway for the treatment of metastatic breast cancer. Sci. Rep. 2017, 7, 46186. [Google Scholar] [CrossRef] [PubMed]
- Amri, J.; Molaee, N.; Baazm, M.; Karami, H. Targeting Epidermal Growth Factor Receptor by MiRNA-145 Inhibits Cell Growth and Sensitizes NSCLC Cells to Erlotinib. Asian Pac. J. Cancer Prev. 2019, 20, 2781–2787. [Google Scholar] [CrossRef] [PubMed]
- Karaayvaz, M.; Zhai, H.; Ju, J. miR-129 promotes apoptosis and enhances chemosensitivity to 5-fluorouracil in colorectal cancer. Cell Death Dis. 2013, 4, e659. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Phung, C.D.; Tran, T.H.; Pham, T.T.; Pham, L.M.; Nguyen, T.T.; Jeong, J.-H.; Choi, H.-G.; Ku, S.K.; Yong, C.S.; et al. Manipulating immune system using nanoparticles for an effective cancer treatment: Combination of targeted therapy and checkpoint blockage miRNA. J. Control. Release 2021, 329, 524–537. [Google Scholar] [CrossRef]
- Kasinski, A.L.; Kelnar, K.; Stahlhut, C.; Orellana, E.; Zhao, J.; Shimer, E.; Dysart, S.; Chen, X.; Bader, A.G.; Slack, F.J. A combinatorial microRNA therapeutics approach to suppressing non-small cell lung cancer. Oncogene 2015, 34, 3547–3555. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Akiyama, Y.; Yuasa, Y. Multiple-to-Multiple Relationships between MicroRNAs and Target Genes in Gastric Cancer. PLoS ONE 2013, 8, e62589. [Google Scholar] [CrossRef]
- Ahir, M.; Upadhyay, P.; Ghosh, A.; Sarker, S.; Bhattacharya, S.; Gupta, P.; Ghosh, S.; Chattopadhyay, S.; Adhikary, A. Delivery of dual miRNA through CD44-targeted mesoporous silica nanoparticles for enhanced and effective triple-negative breast cancer therapy. Biomater. Sci. 2020, 8, 2939–2954. [Google Scholar] [CrossRef]
- Byun, Y.; Choi, Y.-C.; Jeong, Y.; Lee, G.; Yoon, S.; Jeong, Y.; Yoon, J.; Baek, K. MiR-200c downregulates HIF-1α and inhibits migration of lung cancer cells. Cell. Mol. Biol. Lett. 2019, 24, 28. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, S.; Wu, H.; Zhang, L.; Dai, X.; Hu, J.; Xue, J.; Liu, T.; Liang, Y.; Wu, G. MiR-200c Increases the Radiosensitivity of Non-Small-Cell Lung Cancer Cell Line A549 by Targeting VEGF-VEGFR2 Pathway. PLoS ONE 2013, 8, e78344. [Google Scholar] [CrossRef] [PubMed]
- McArthur, K.; Feng, B.; Wu, Y.; Chen, S.; Chakrabarti, S. MicroRNA-200b Regulates Vascular Endothelial Growth Factor–Mediated Alterations in Diabetic Retinopathy. Diabetes 2011, 60, 1314–1323. [Google Scholar] [CrossRef]
- Van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: A first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- Viteri, S.; Rosell, R. An innovative mesothelioma treatment based on miR-16 mimic loaded EGFR targeted minicells (TargomiRs). Transl. Lung Cancer Res. 2018, 7, S1–S4. [Google Scholar] [CrossRef]
- Jebbawi, F.; Fayyad-Kazan, H.; Merimi, M.; Lewalle, P.; Verougstraete, J.-C.; Leo, O.; Romero, P.; Burny, A.; Badran, B.; Martiat, P.; et al. A microRNA profile of human CD8+ regulatory T cells and characterization of the effects of microRNAs on Treg cell-associated genes. J. Transl. Med. 2014, 12, 218. [Google Scholar] [CrossRef]
- Liu, Y.; Nie, W.; Jin, Y.; Zhuo, A.; Zang, Y.; Xiu, Q. B and T Lymphocyte Attenuator is a Target of miR-155 during Naïve CD4+ T Cell Activation. Iran. J. Immunol. 2016, 13, 89–99. [Google Scholar]
- Huffaker, T.B.; Lee, S.-H.; Tang, W.W.; Wallace, J.A.; Alexander, M.; Runtsch, M.C.; Larsen, D.K.; Thompson, J.; Ramstead, A.G.; Voth, W.P.; et al. Antitumor immunity is defective in T cell–specific microRNA-155–deficient mice and is rescued by immune checkpoint blockade. J. Biol. Chem. 2017, 292, 18530–18541. [Google Scholar] [CrossRef]
- Singh, A.; Srivastava, N.; Yadav, A.; Ateeq, B. Targeting AGTR1/NF-κB/CXCR4 axis by miR-155 attenuates oncogenesis in glioblastoma. Neoplasia 2020, 22, 497–510. [Google Scholar] [CrossRef]
- Zhang, T.; Davidson-Moncada, J.K.; Mukherjee, P.; Furman, R.R.; Bhavsar, E.; Chen, Z.; Hakimpour, P.; Papavasiliou, N.; Tam, W. MicroRNA-155 regulates casein kinase 1 gamma 2: A potential pathogenetic role in chronic lymphocytic leukemia. Blood Cancer J. 2017, 7, e606. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, P.; Bill, K.; Liu, J.; Young, E.; Peng, T.; Bolshakov, S.; Hoffman, A.; Song, Y.; Demicco, E.G.; Terrada, D.L.; et al. MiR-155 Is a Liposarcoma Oncogene That Targets Casein Kinase-1α and Enhances β-Catenin Signaling. Cancer Res. 2012, 72, 1751–1762. [Google Scholar] [CrossRef]
- Seto, A.G.; Beatty, X.; Lynch, J.M.; Hermreck, M.; Tetzlaff, M.; Duvic, M.; Jackson, A.L. Cobomarsen, an oligonucleotide inhibitor of miR-155, co-ordinately regulates multiple survival pathways to reduce cellular proliferation and survival in cutaneous T-cell lymphoma. Br. J. Haematol. 2018, 183, 428–444. [Google Scholar] [CrossRef] [PubMed]
- Witten, L.; Slack, F.J. miR-155 as a novel clinical target for hematological malignancies. Carcinogenesis 2020, 41, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Sha, H.-H.; Wang, D.-D.; Chen, D.; Liu, S.-W.; Wang, Z.; Yan, D.-L.; Dong, S.-C.; Feng, J.-F. MiR-138: A promising therapeutic target for cancer. Tumor Biol. 2017, 39, 101042831769757. [Google Scholar] [CrossRef] [PubMed]
- Shadbad, M.A.; Safaei, S.; Brunetti, O.; Derakhshani, A.; Lotfinejad, P.; Mokhtarzadeh, A.; Hemmat, N.; Racanelli, V.; Solimando, A.G.; Argentiero, A.; et al. A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery. Genes 2021, 12, 1206. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.-K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.-L.; Kim, T.-Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. miRNA-based biomarkers, therapies, and resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef]
- Orellana, E.A.; Li, C.; Lisevick, A.; Kasinski, A.L. Identification and validation of microRNAs that synergize with miR-34a—A basis for combinatorial microRNA therapeutics. Cell Cycle 2019, 18, 1798–1811. [Google Scholar] [CrossRef]
- Meng, F.; Chen, Y.; Yang, M.; Zhang, H.; Wang, W. Concomitant inhibition of B7-H3 and PD-L1 expression by a novel and synthetic microRNA delivers potent antitumor activities in colorectal tumor models. Investig. New Drugs 2021, 39, 1267–1274. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).