
Comparative Ubiquitome Analysis Reveals Deubiquitinating Effects Induced by Wolbachia Infection in Drosophila melanogaster


Abstract
:1. Introduction
2. Results
2.1. Proteomic Profiles of Wolbachia-Infected and Uninfected Drosophila Testes
2.2. Analysis of Differentially Expressed Proteins between DWT and DTT
2.3. Proteome-Wide Analysis of Ubiquitinated Proteins in DWT and DTT
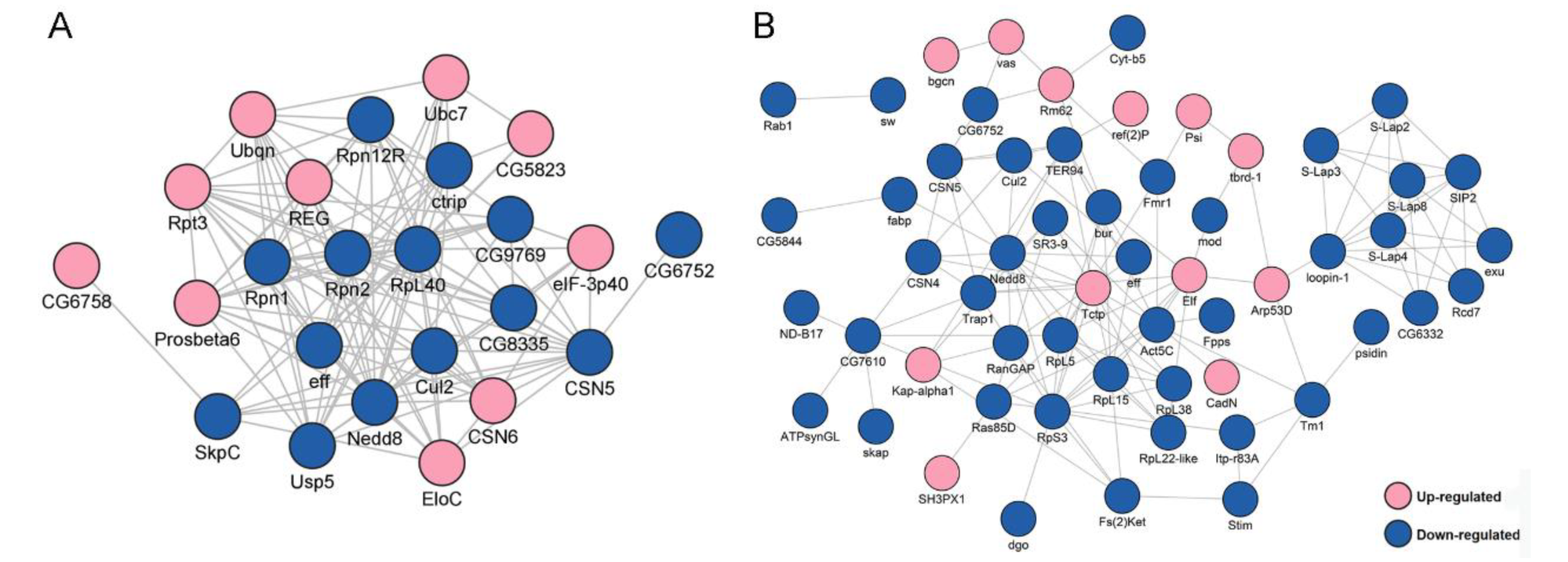
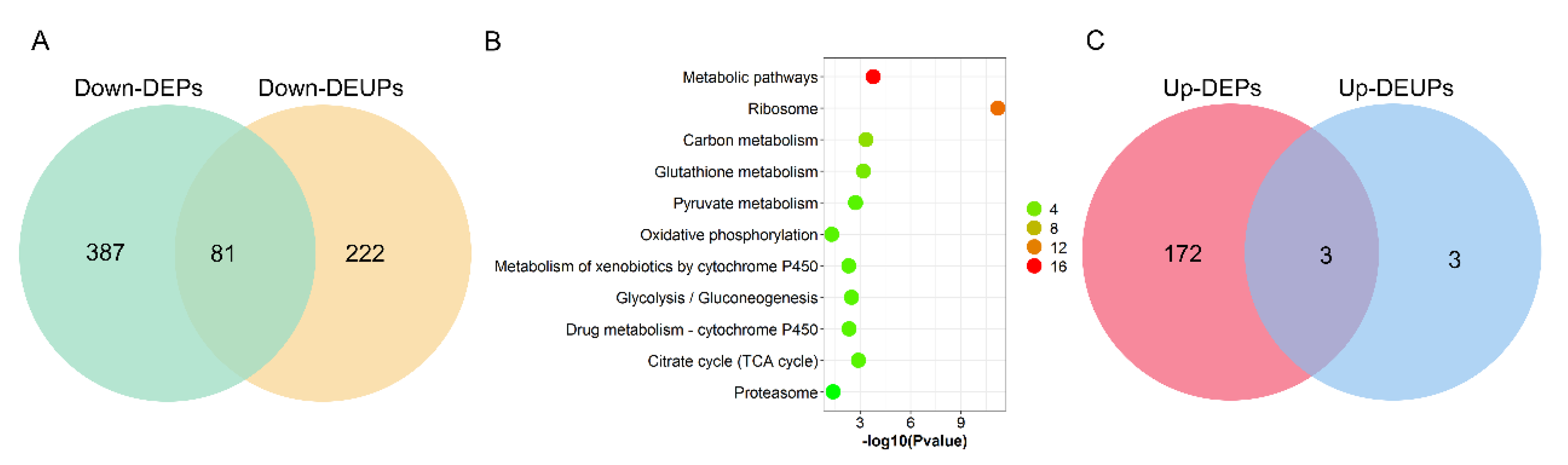
2.4. Analysis of Differentially Expressed Ubiquitinated Proteins (DEUPs) between DTT and DWT
2.5. The Correlation Analysis between DEPs and DEUPs
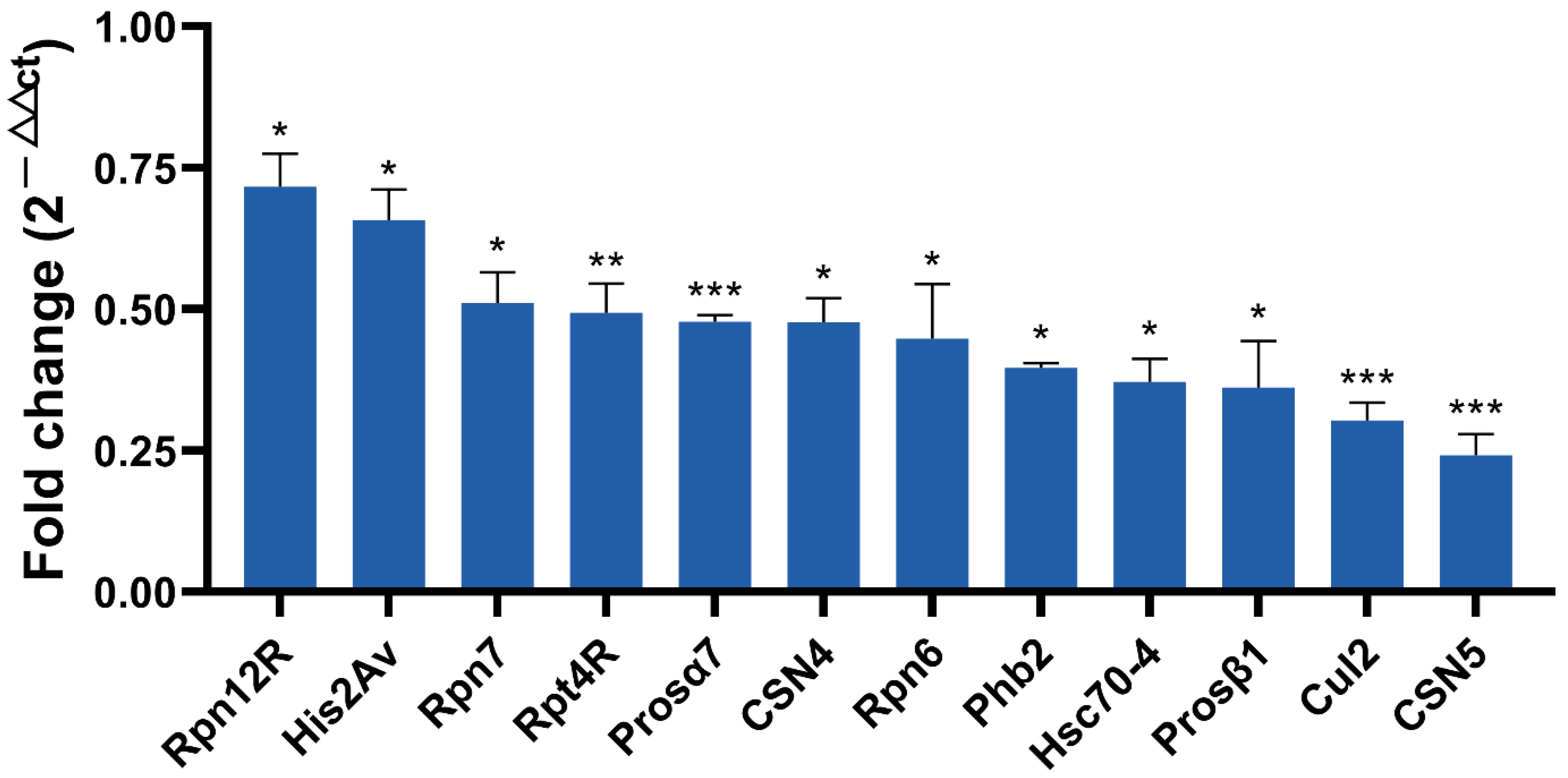
2.6. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Validation and Screening
2.7. Functional Analysis of Candidates in Male Fertility
3. Discussion
4. Materials and Methods
4.1. Fly Lines
4.2. Protein Extraction
4.3. Acetone Precipitation and Trypsin Digestion
4.4. Affinity Enrichment of the Ubiquitinated Protein (IP)
4.5. LC-MS/MS Analysis
4.6. Database Search and Bioinformatics Analysis
4.7. qRT-PCR
4.8. Fertility Test
4.9. Immunofluorescent Staining
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Genet. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Shropshire, J.D.; Cross, K.L.; Leigh, B.; Mansueto, A.J.; Stewart, V.; Bordenstein, S.R.; Bordenstein, S.R. Living in the endosymbiotic world of Wolbachia: A centennial review. Cell Host Microbe 2021, 29, 879–893. [Google Scholar] [CrossRef] [PubMed]
- Horard, B.; Terretaz, K.; Gosselin-Grenet, A.-S.; Sobry, H.; Sicard, M.; Landmann, F.; Loppin, B. Paternal transmission of the Wolbachia CidB toxin underlies cytoplasmic incompatibility. Curr. Biol. 2022, 32, 1319–1331.e5. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Leigh, B.A.; Ritchie, I.T.; Bordenstein, S.R. The Cif proteins from Wolbachia prophage WO modify sperm genome integrity to establish cytoplasmic incompatibility. PLoS Biol. 2022, 20, e3001584. [Google Scholar] [CrossRef]
- Grabbe, C.; Husnjak, K.; Dikic, I. The spatial and temporal organization of ubiquitin networks. Nat. Rev. Mol. Cell Biol. 2011, 12, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J. Biosci. 2006, 31, 137–155. [Google Scholar] [CrossRef]
- Nakamura, N. Ubiquitination Regulates the Morphogenesis and Function of Sperm Organelles. Cells 2013, 2, 732–750. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.-C.; Yang, W.-X. New insights to the ubiquitin–proteasome pathway (UPP) mechanism during spermatogenesis. Mol. Biol. Rep. 2012, 40, 3213–3230. [Google Scholar] [CrossRef]
- Cafe, S.L.; Nixon, B.; Ecroyd, H.; Martin, J.H.; Skerrett-Byrne, D.A.; Bromfield, E.G. Proteostasis in the Male and Female Germline: A New Outlook on the Maintenance of Reproductive Health. Front. Cell Dev. Biol. 2021, 9, 660626. [Google Scholar] [CrossRef]
- Xiong, Y.; Yu, C.; Zhang, Q. Ubiquitin-Proteasome System–Regulated Protein Degradation in Spermatogenesis. Cells 2022, 11, 1058. [Google Scholar] [CrossRef]
- Fabian, L.; Brill, J.A. Drosophila spermiogenesis: Big things come from little packages. Spermatogenesis 2012, 2, 197–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Kim, J.; Fuller, M.T. The polyubiquitin gene Ubi-p63E is essential for male meiotic cell cycle progression and germ cell differentiation in Drosophila. Development 2013, 140, 3522–3531. [Google Scholar] [CrossRef] [Green Version]
- Baarends, W.M.; Hoogerbrugge, J.W.; Roest, H.P.; Ooms, M.; Vreeburg, J.; Hoeijmakers, J.H.; Grootegoed, J. Histone Ubiquitination and Chromatin Remodeling in Mouse Spermatogenesis. Dev. Biol. 1999, 207, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Kavarthapu, R.; Anbazhagan, R.; Sharma, A.; Shiloach, J.; Dufau, M.L. Linking Phospho-Gonadotropin Regulated Testicular RNA Helicase (GRTH/DDX25) to Histone Ubiquitination and Acetylation Essential for Spermatid Development During Spermiogenesis. Front. Cell Dev. Biol. 2020, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.-Y.; Wu, J.; Ye, L.; Gavrilina, G.B.; Saunders, T.L.; Yu, X. RNF8-Dependent Histone Modifications Regulate Nucleosome Removal during Spermatogenesis. Dev. Cell 2010, 18, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavarthapu, R.; Anbazhagan, R.; Raju, M.; Morris, C.-H.T.; Pickel, J.; Dufau, M.L. Targeted knock-in mice with a human mutation in GRTH/DDX25 reveals the essential role of phosphorylated GRTH in spermatid development during spermatogenesis. Hum. Mol. Genet. 2019, 28, 2561–2572. [Google Scholar] [CrossRef]
- Vedelek, V.; Kovács, A.L.; Juhász, G.; Alzyoud, E.; Sinka, R. The tumor suppressor archipelago E3 ligase is required for spermatid differentiation in Drosophila testis. Sci. Rep. 2021, 11, 8422. [Google Scholar] [CrossRef]
- Beckmann, J.F.; Van Vaerenberghe, K.; Akwa, D.E.; Cooper, B.S. A single mutation weakens symbiont-induced reproductive manipulation through reductions in deubiquitylation efficiency. Proc. Natl. Acad. Sci. USA 2021, 118, e2113271118. [Google Scholar] [CrossRef]
- Shropshire, J.D.; Hamant, E.; Cooper, B.S. Male Age and Wolbachia Dynamics: Investigating How Fast and Why Bacterial Densities and Cytoplasmic Incompatibility Strengths Vary. mBio 2021, 12, 6. [Google Scholar] [CrossRef]
- Yuan, L.-L.; Chen, X.; Zong, Q.; Zhao, T.; Wang, J.-L.; Zheng, Y.; Zhang, M.; Wang, Z.; Brownlie, J.C.; Yang, F.; et al. Quantitative Proteomic Analyses of Molecular Mechanisms Associated with Cytoplasmic Incompatibility in Drosophila melanogaster Induced by Wolbachia. J. Proteome Res. 2015, 14, 3835–3847. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, Z.; Huang, X. OSBP- and FAN-mediated sterol requirement for spermatogenesis in Drosophila. Development 2010, 137, 3775–3784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Wang, W.; Salvaterra, P.M. Functional analysis and tissue-specific expression of Drosophila Na+, K+-ATPase subunits. J. Neurochem. 2002, 71, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Copeland, J.M.; Cho, J.; Lo, T.; Hur, J.H.; Bahadorani, S.; Arabyan, T.; Rabie, J.; Soh, J.; Walker, D.W. Extension of Drosophila Life Span by RNAi of the Mitochondrial Respiratory Chain. Curr. Biol. 2009, 19, 1591–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, T.; Amrein, H. Neuronal Gluconeogenesis Regulates Systemic Glucose Homeostasis in Drosophila melanogaster. Curr. Biol. 2019, 29, 1263–1272.e5. [Google Scholar] [CrossRef] [PubMed]
- Tekotte, H.; Berdnik, D.; Török, T.; Buszczak, M.; Jones, L.M.; Cooley, L.; Knoblich, J.; Davis, I. Dcas Is Required for importin-α3 Nuclear Export and Mechano-Sensory Organ Cell Fate Specification in Drosophila. Dev. Biol. 2002, 244, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Kotov, A.A.; Olenkina, O.M.; Kibanov, M.V.; Olenina, L.V. RNA helicase Belle (DDX3) is essential for male germline stem cell maintenance and division in Drosophila. Biochim. Biophys. Acta 2016, 1863, 1093–1105. [Google Scholar] [CrossRef]
- Vedelek, V.; Laurinyecz, B.; Kovács, A.L.; Juhász, G.; Sinka, R. Testis-specific Bb8 is essential in the development of spermatid mitochondria. PLoS ONE 2016, 11, e0161289. [Google Scholar] [CrossRef] [Green Version]
- Laurinyecz, B.; Vedelek, V.; Kovács, A.L.; Szilasi, K.; Lipinszki, Z.; Slezák, C.; Darula, Z.; Juhász, G.; Sinka, R. Sperm-Leucylaminopeptidases are required for male fertility as structural components of mitochondrial paracrystalline material in Drosophila melanogaster sperm. PLoS Genet. 2019, 15, e1007987. [Google Scholar] [CrossRef]
- Eberhart, C.G.; Maines, J.Z.; Wasserman, S.A. Meiotic cell cycle requirement for a fly homologue of human Deleted in Azoospermia. Nature 1996, 381, 783–785. [Google Scholar] [CrossRef]
- Gao, L.; Chang, S.; Xia, W.; Wang, X.; Zhang, C.; Cheng, L.; Liu, X.; Chen, L.; Shi, Q.; Huang, J.; et al. Circular RNAs from BOULE play conserved roles in protection against stress-induced fertility decline. Sci. Adv. 2020, 6, eabb7426. [Google Scholar] [CrossRef]
- Zhang, H.-B.; Cao, Z.; Qiao, J.-X.; Zhong, Z.-Q.; Pan, C.-C.; Liu, C.; Zhang, L.-M.; Wang, Y.-F. Metabolomics provide new insights into mechanisms of Wolbachia-induced paternal defects in Drosophila melanogaster. PLoS Pathog. 2021, 17, e1009859. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.; Miao, Y.; Xiao, J.; Huang, D. Association of Wolbachia with Gene Expression in Drosophila Testes. Microb. Ecol. 2021, 82, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-J.; Cui, J.-R.; Chen, J.; Bing, X.-L.; Hong, X.-Y. Proteomic analysis of Laodelphax striatellus gonads reveals proteins that may manipulate host reproduction by Wolbachia. Insect Biochem. Mol. Biol. 2019, 113, 103211. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Heredia, J.; Estanyol, J.M.; Ballescà, J.L.; Oliva, R. Proteomic identification of human sperm proteins. Proteomics 2006, 6, 4356–4369. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, T.; Xie, F.; Liu, T.; Nicora, C.D.; Yang, F.; Camp, D.G.; Smith, R.; Roberts, K.P. A Systematic Analysis of a Deep Mouse Epididymal Sperm Proteome1. Biol. Reprod. 2012, 87, 141. [Google Scholar] [CrossRef]
- Xu, Y.; Han, Q.; Ma, C.; Wang, Y.; Zhang, P.; Li, C.; Cheng, X.; Xu, H. Comparative Proteomics and Phosphoproteomics Analysis Reveal the Possible Breed Difference in Yorkshire and Duroc Boar Spermatozoa. Front. Cell Dev. Biol. 2021, 9, 652809. [Google Scholar] [CrossRef]
- Nynca, J.; Arnold, G.J.; Fröhlich, T.; Otte, K.; Ciereszko, A. Proteomic identification of rainbow trout sperm proteins. Proteomics 2014, 14, 1569–1573. [Google Scholar] [CrossRef]
- Mageeney, C.M.; Kearse, M.G.; Gershman, B.W.; Pritchard, C.E.; Colquhoun, J.M.; Ware, V.C. Functional interplay between ribosomal protein paralogues in the eRpL22 family in Drosophila melanogaster. Fly 2018, 12, 143–163. [Google Scholar] [CrossRef] [Green Version]
- Mageeney, C.M.; Ware, V.C. Specialized eRpL22 paralogue-specific ribosomes regulate specific mRNA translation in spermatogenesis in Drosophila melanogaster. Mol. Biol. Cell 2019, 30, 2240–2253. [Google Scholar] [CrossRef]
- Sutovsky, P. Sperm proteasome and fertilization. Reproduction 2011, 142, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Ji, S.-Y.; Busayavalasa, K.; Shao, J.; Yu, C. Meiosis I progression in spermatogenesis requires a type of testis-specific 20S core proteasome. Nat. Commun. 2019, 10, 3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Katz, E.; Belote, J.M. Expression of proteasome subunit isoforms during spermatogenesis in Drosophila melanogaster. Insect Mol. Biol. 2002, 11, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Belote, J.M. The testis-specific proteasome subunit Prosalpha6T of D. melanogaster is required for individualization and nuclear maturation during spermatogenesis. Development 2007, 134, 3517–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Lan, X.; Chen, X.; Yu, C.; Xu, Y.; Liu, Y.; Xu, L.; Fan, H.Y.; Tong, C. Protein synthesis and degradation are essential to regulate germline stem cell homeostasis in Drosophila testes. Development 2016, 143, 2930–2945. [Google Scholar] [PubMed] [Green Version]
- Sutovsky, P.; Moreno, R.; Ramalho-Santos, J.; Dominko, T.; Simerly, C.; Schatten, G. Ubiquitinated Sperm Mitochondria, Selective Proteolysis, and the Regulation of Mitochondrial Inheritance in Mammalian Embryos1. Biol. Reprod. 2000, 63, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Berger, K.H.; Yaffe, M.P. Prohibitin Family Members Interact Genetically with Mitochondrial Inheritance Components in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 4043–4052. [Google Scholar] [CrossRef] [Green Version]
- Sutovsky, P.; Van Leyen, K.; McCauley, T.; Day, B.N.; Sutovsky, M. Degradation of paternal mitochondria after fertilization: Implications for heteroplasmy, assisted reproductive technologies and mtDNA inheritance. Reprod. Biomed. Online 2004, 8, 24–33. [Google Scholar] [CrossRef]
- Romero, M.R.; Pérez-Figueroa, A.; Carrera, M.; Swanson, W.J.; Skibinski, D.O.F.; Diz, A.P. RNA-seq coupled to proteomic analysis reveals high sperm proteome variation between two closely related marine mussel species. J. Proteom. 2019, 192, 169–187. [Google Scholar] [CrossRef]
- Shoemaker, D.D.; Katju, V.; Jaenike, J. Wolbachia and the evolution of reproductive isolation between Drosophila recens and Drosophila subquinaria. Evolution 1999, 53, 1157–1164. [Google Scholar] [CrossRef]
- Elias-Costa, A.J.; Confalonieri, V.A.; Lanteri, A.A.; Rodriguero, M.S. Game of clones: Is Wolbachia inducing speciation in a weevil with a mixed reproductive mode? Mol. Phylogenet. Evol. 2019, 133, 42–53. [Google Scholar] [CrossRef]
- Cruz, M.A.; Magalhães, S.; Sucena, É.; Zélé, F. Wolbachia and host intrinsic reproductive barriers contribute additively to postmating isolation in spider mites. Evolution 2021, 75, 2085–2101. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhou, H.; Ponzoni, L.; Luo, A.; Zhu, R.; He, M.; Huang, Y.; Guan, K.-L.; Bahar, I.; Liu, Z.; et al. EIF3H Orchestrates Hippo Pathway–Mediated Oncogenesis via Catalytic Control of YAP Stability. Cancer Res. 2020, 80, 2550–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasanami, T.; Sugiura, K.; Tokumoto, T.; Yoshizaki, N.; Dohra, H.; Nishio, S.; Mizushima, S.; Hiyama, G.; Matsuda, T. Sperm proteasome degrades egg envelope glycoprotein ZP1 during fertilization of Japanese quail (Coturnix japonica). Reproduction 2012, 144, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, T.; Sawada, H. Fertilization of Ascidians: Gamete Interaction, Self/Nonself Recognition and Sperm Penetration of Egg Coat. Front. Cell Dev. Biol. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, F.; Zheng, M.; Wu, R.; Yang, T.; Chen, X.; Shu, J. Effect of Sperm 26S Proteasome on Embryo Formation and Development in In Vitro Fertilization. Reprod. Sci. 2022, 29, 1844–1850. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Mysteries and unsolved problems of mammalian fertilization and related topics. Biol. Reprod. 2022, 106, 644–675. [Google Scholar] [CrossRef] [PubMed]
- Azuma, M.; Ogata, T.; Yamazoe, K.; Tanaka, Y.; Inoue, Y.H. Heat shock cognate 70 genes contribute to Drosophila spermatocyte growth progression possibly through the insulin signaling pathway. Dev. Growth Differ. 2021, 63, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Lier, S.; Paululat, A. The proteasome regulatory particle subunit Rpn6 is required for Drosophila development and interacts physically with signalosome subunit Alien/CSN2. Gene 2002, 298, 109–119. [Google Scholar] [CrossRef]
- Hoffmann, A.; Turelli, M. Unidirectional incompatibility in Drosophila simulans: Inheritance, geographic variation and fitness effects. Genetics 1988, 119, 435–444. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Rpn12R | GGGCAGATACAACAAGATA | CCTCGTCCAGACACCACT |
| Rpn7 | TGCCTTCCGCAAGACCTA | CGCCACCGAGTAAACACC |
| Rpt4R | GGTCGGGGAAATCCTGAAGC | AGTGGTCACATCCAGGGCGA |
| Prosα7 | AGGCAGCCAACTACAGACA | GAAGTAACCGAAGGAGGAG |
| CSN4 | GGAGACGGGTCAGAAACA | TCCAGGACACGAGCATAA |
| Rpn6 | TCACGCACTGAGCAACCT | CTTTGGCGGACAGTAGATAG |
| Prosβ1 | GCGGAGTGGTCATTGGAG | TCGTGATAGTTCAGCGAGTAG |
| Phb2 | CGGCATCCAGAGCGACAT | GCAGATAGGGCAGGTTCA |
| His2Av | GTGGGTCGCATCCATCGT | CCTCGGCGGTCAGGTATT |
| Cul2 | AAGAGTGCGAGGAGAAGT | TGAGATTATCGGGTATGG |
| CSN5 | AGGTGATGGGTCTAATGCT | AATGGCTCCTGGTATGTCT |
| Hsc70-4 | CCTCGGCGGTCAGGTATT | TGCCGAGCAGGTTGTTGT |
| rp49 | CGGTTACGGATCGAACAAGC | CTTGCGCTTCTTGGAGGAGA |
| wsp | TGGTCCAATAAGTGATGAAGAAAC | AAAAATTAAACGCTACTCCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zong, Q.; Mao, B.; Zhang, H.-B.; Wang, B.; Yu, W.-J.; Wang, Z.-W.; Wang, Y.-F. Comparative Ubiquitome Analysis Reveals Deubiquitinating Effects Induced by Wolbachia Infection in Drosophila melanogaster. Int. J. Mol. Sci. 2022, 23, 9459. https://doi.org/10.3390/ijms23169459
Zong Q, Mao B, Zhang H-B, Wang B, Yu W-J, Wang Z-W, Wang Y-F. Comparative Ubiquitome Analysis Reveals Deubiquitinating Effects Induced by Wolbachia Infection in Drosophila melanogaster. International Journal of Molecular Sciences. 2022; 23(16):9459. https://doi.org/10.3390/ijms23169459
Chicago/Turabian StyleZong, Qiong, Bin Mao, Hua-Bao Zhang, Bing Wang, Wen-Juan Yu, Zhi-Wei Wang, and Yu-Feng Wang. 2022. "Comparative Ubiquitome Analysis Reveals Deubiquitinating Effects Induced by Wolbachia Infection in Drosophila melanogaster" International Journal of Molecular Sciences 23, no. 16: 9459. https://doi.org/10.3390/ijms23169459