Abstract
Ca2+ is a ubiquitous second messenger, which allows eukaryotic cells to respond to external stimuli. The use of genetically encoded Ca2+ indicators allows real-time monitoring of cytosolic Ca2+ levels to study such responses. Here we explored the possibility of using the ratiometric Ca2+ indicator GEM-GECO for monitoring cytosolic Ca2+ concentration ([Ca2+]cyt) in the yeast Ogataea parapolymorpha. High-level production of GEM-GECO led to a severe growth defect in cells lacking the vacuolar Ca2+ ATPase Pmc1, which is involved in [Ca2+]cyt control, and prompted a phenotype resembling that of Pmc1 deficiency, in a strain with wild-type PMC1. This was likely due to the presence of the calmodulin domain in GEM-GECO. In contrast to previous studies of genetically-encoded calcium indicators in neuronal cells, our results suggest that physiological effects of GEM-GECO expression in yeast cells are due not to Ca2+ depletion, but to excessive Ca2+ signaling. Despite these drawbacks, study of fluorescence in individual cells revealed switching of GEM-GECO from the Ca2+-free to Ca2+-bound state minutes after external addition of CaCl2. This was followed by gradual return of GEM-GECO to a Ca2+-free-state that was impaired in the pmc1-Δ mutant. These results demonstrate GEM-GECO usability for [Ca2+]cyt monitoring in budding yeast.
1. Introduction
Ca2+ is a ubiquitous second messenger involved in the response to external stimuli in different types of living cells. In particular, it is involved in neuron activity, muscular contraction, cell cycle regulation, etc. [1,2,3,4]. Development of techniques for Ca2+ imaging in living cells has provided powerful tools to study its signaling roles.
Intracellular Ca2+ concentration can be monitored either by staining with fluorescent dyes (e.g., Fura or Fluo-2,3,4) or by using genetically encoded calcium sensors such as the Ca2+- coelenterazine-dependent luciferase aequorin [5,6] or artificially developed indicators based on fluorescent proteins (for a review see [7]). Among the latter, the most widely used type of genetically encoded Ca2+ indicators (GECI) is represented by circularly-permuted fluorescent proteins fused with calmodulin (CaM) at the C-terminus and possessing a Ca2+/CaM binding peptide at the N-terminus (reviewed in [7,8]). Efforts to improve their characteristics resulted in highly efficient GECIs (e.g., described in [9,10,11]) which are applicable for monitoring cytosolic Ca2+ concentration ([Ca2+]cyt) in different types of animal cells. One of these indicators was used to monitor [Ca2+]cyt in dividing cells of the Schizosaccharomyces pombe yeast [4]. In such indicators, the Ca2+ dependent binding of the N-terminal peptide to the CaM domain leads to an alteration in the fluorescence parameters. As a rule, this is an increase in the fluorescence intensity; however, ratiometric GECI, which change either excitation (GEX-GECO) or emission (GEM-GECO) wavelength upon Ca2+ binding, have also been developed [9]. This property should allow monitoring [Ca2+]cyt in individual cells independently of variation in the GECI production level. Notably, a GECI is effective in this kind of experiments if its presence in predominantly Ca2+-free form switches to predominantly Ca2+ -bound form within the range of the physiological [Ca2+]cyt variations. This means that the Kd of its Ca2+-bound form must be between the resting [Ca2+]cyt level and the level reached after stimulation.
Notably, high levels of GECI production were shown to affect neuronal cell physiology due to depletion of cytosolic Ca2+ [12], which may hamper its use in certain experiments. Although there is an example of the use of a GECI in fission yeast Sch. pombe [4], to our knowledge, no data are available on the effects of such proteins on yeast cell physiology and their applicability for [Ca2+]cyt monitoring in various species of budding yeasts.
Similar to other eukaryotes, yeasts use Ca2+ as a second messenger allowing cells to respond to different external stimuli. This requires maintaining [Ca2+]cyt at a low level (50–200 nM) by pumping it from the cytosol into intracellular depots. Studies of Saccharomyces cerevisiae have demonstrated that the main Ca2+ depot in the yeast cell is the vacuole, which accumulates Ca2+ due to action of the vacuolar calcium ATPase Pmc1 and the Ca2+/H+ exchanger Vcx1. The proton gradient, which empowers Vcx1 action, is generated by Vma1 (for a review see [13,14]).
Certain extracellular stimuli cause an increase in [Ca2+]cyt, activating the Ca2+ binding protein CaM, which interacts with CaM-dependent proteins, thus modulating their activity. One of these proteins is the protein phosphatase calcineurin which is activated both by binding Ca2+ directly and by interaction with Ca2+-bound CaM. Calcineurin dephosphorylates transcription factor Crz1 to trigger its translocation to the nucleus. This alters the expression of a number of genes including those involved in maintaining Ca2+ homeostasis [15], thus allowing the cell to restore [Ca2+]cyt to the resting level.
Previously we have observed that inactivation of the vacuolar Ca2+ ATPase Pmc1 in methylotrophic yeast Ogataea (Hansenula) polymorpha and O. parapolymorpha has a wider impact on cell physiology than in the conventional yeast S. cerevisiae. In particular, besides the inability to grow at high concentration of Ca2+, in Ogataea yeasts this mutation causes hypersensitivity to sodium dodecyl sulfate (SDS), which is related to G2-M cell cycle transition arrest [16]. Here we used O. parapolymorpha to study whether the GEM-GECO indicator is suitable for [Ca2+]cyt monitoring in this yeast and characterize the effects of its production on cell physiology in wild-type and pmc1-Δ strains.
2. Results and Discussion
2.1. GEM-GECO Expression Affects Cell Growth in a Pmc1-Dependent Manner
Efficient monitoring of Ca2+ using a genetically encoded fluorescent indicator such as GEM-GECO demands that its fluorescence be observable and quantifiable over the background fluorescence of the host cells. To achieve this in O. parapolymorpha, we used a codon-optimized gene coding for GEM-GECO, which was placed under control of a strong regulatable promoter of the O. parapolymorpha MAL1 gene. The latter codes for maltase, which is required for utilization of some disaccharides including sucrose and is strongly induced upon growth on these carbon sources [17]. At the same time, utilization of sucrose supports growth of O. parapolymorpha as efficiently as that of glucose (our unpublished observation). In our study, this provides an advantage compared to the well-known promoters of methylotrophic yeasts, which are induced during utilization of the highly unfavorable carbon source methanol. The GEM-GECO expression cassette was placed into a vector possessing the G418 resistance marker and integrated into the O. polymorpha DL1-L genome. To select clones efficiently expressing GEM-GECO, the obtained transformants were grown on YP-Suc plates and irradiated by a 402 nm light emitting diode that excited GEM-GECO fluorescence. A clone with the brightest fluorescence which demonstrated high mitotic stability was designated DL5 and used in further experiments.
To test applicability of GEM-GECO for monitoring [Ca2+]cyt in O. parapolymorpha, apart from the wild-type strain we also used a derivative of DL5 lacking the vacuolar Ca2+ ATPase Pmc1, which is responsible for maintaining [Ca2+]cyt at a low level.
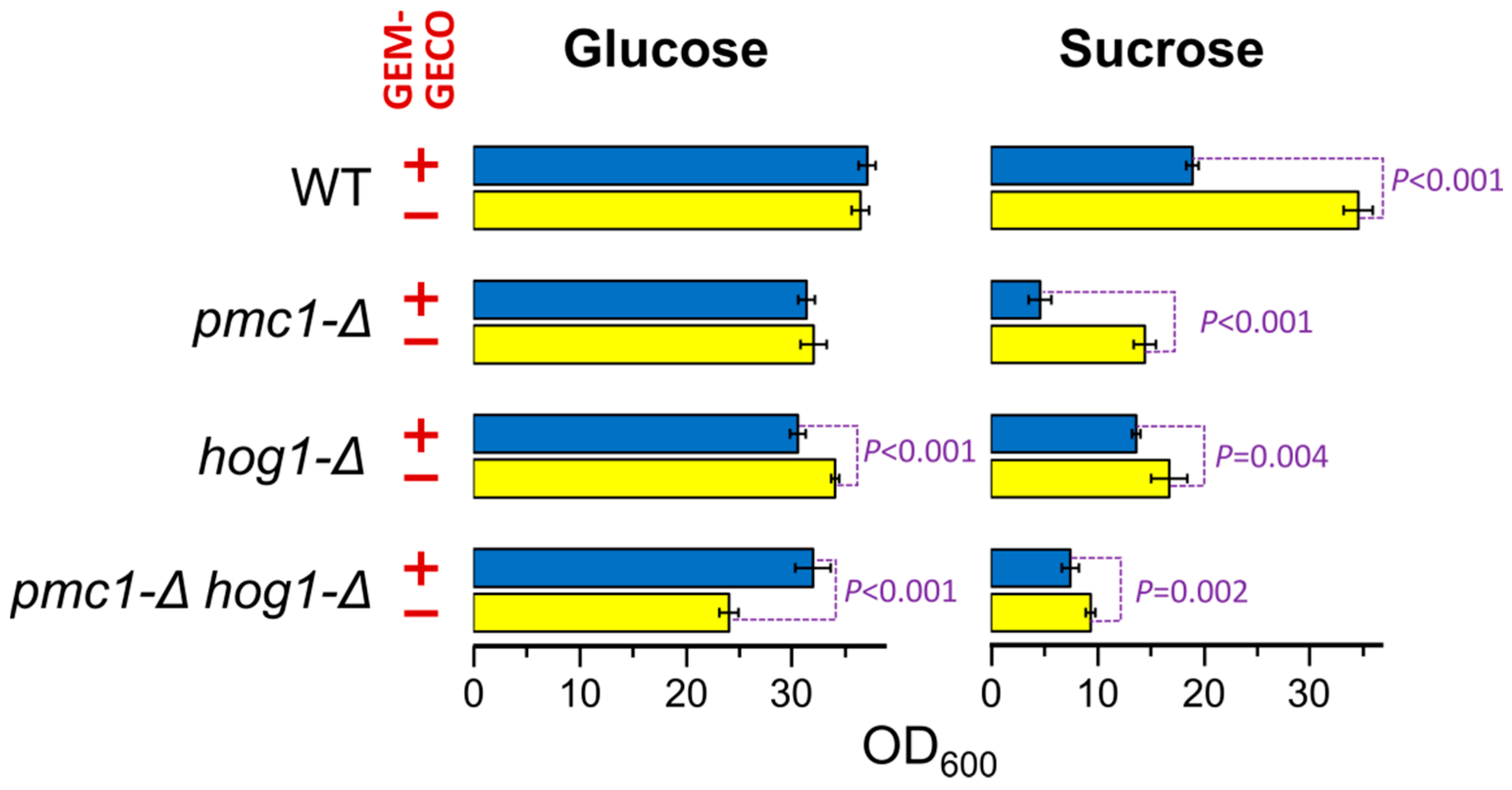
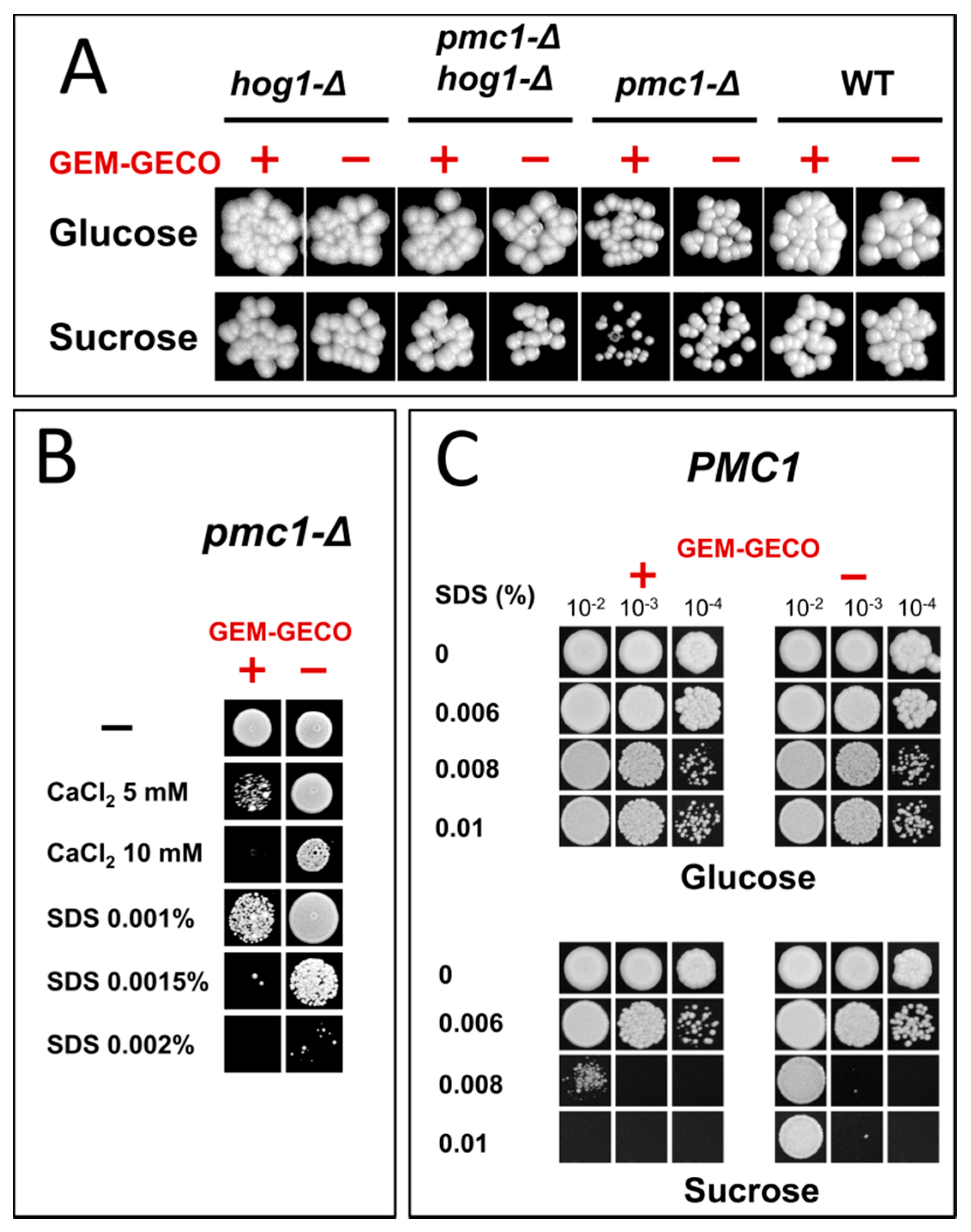
Presence of the GEM-GECO expression cassette did not noticeably affect cell growth in medium containing glucose, which represses the MAL1 promoter. However, the culture of the wild-type strain possessing the GEM-GECO expression cassette did not reach the same density as the control strain lacking this cassette when cells were grown in sucrose-containing medium to stationary phase (Figure 1). Surprisingly, inactivation of PMC1 noticeably decreased final cell density in sucrose-containing medium even in the absence of the GEM-GECO expression cassette, while its presence led to a deeper decrease in the final optical density of the pmc1-Δ mutant than in the strain with wild-type PMC1 (Figure 1). Colonies of the pmc1-Δ mutant bearing the GEM-GECO expression cassette grew on sucrose-containing medium noticeably slower than those of the pmc1-Δ strain lacking this cassette (Figure 2A). This indicated that GEM-GECO expression aggravates some negative effects of the pmc1-Δ mutation. It was previously shown, that the Pmc1 deficiency in O. polymorpha and O. parapolymorpha causes hypersensitivity to SDS, which is related to the Hog1 and Wee1 dependent cell cycle regulation [16]. Although deletion of the HOG1 gene itself affected cell growth, it reduced the negative effect of the GEM-GECO expression, since there was only small difference in stationary OD600 (Figure 1) and no noticeable difference in size of colonies (Figure 2A) between hog1-Δ mutant strains with and without the GEM-GECO expression cassette. The presence of the GEM-GECO expression cassette aggravated phenotypes of Pmc1 deficiency even when its promoter was repressed. Indeed, sensitivities to SDS and Ca2+ of the pmc1-Δ mutant possessing the GEM-GECO expression cassette were increased even on glucose-containing medium (Figure 2B). Expression of GEM-GECO in the strain with wild-type PMC1 diminished the resistance to SDS (Figure 2C) resembling the manifestation of PMC1 inactivation [16], which is most likely due to increased [Ca2+]cyt. This may indicate that GEM-GECO expression provokes activation of some Ca2+-dependent signaling pathway(s) that are usually triggered via increased, rather than depleted Ca2+ in the cytosol.
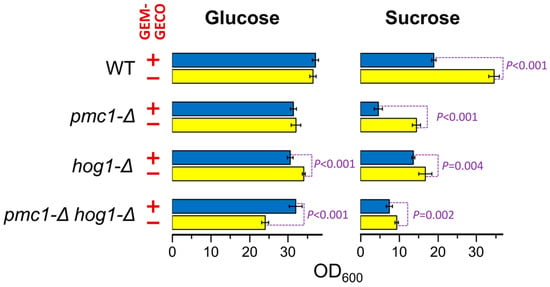
Figure 1.
Effect of GEM-GECO expression on OD600 of saturated cultures of strains bearing wild-type or deletion alleles of the PMC1 and HOG1 genes. Overnight YPD cultures were 100-fold diluted with YPD (Glucose) or YP-Suc (Sucrose) medium and incubated at 37 °C for 24 h. WT, DL5-LC (+) and DL1-LC (−) strains; pmc1-Δ, DL5-pmc1-LC (+) and DL-pmc1-LC (−) strains; hog1-Δ, DL5-hog1 (+) and DLdaduA-hog1-UC (−) strains; pmc1-Δ hog1-Δ, DL5-pmc1-hog1 (+) and DLdaduA-pmc1-hog1 (−) strains. The means with standard deviations and P-values were calculated from data obtained in six replicates.
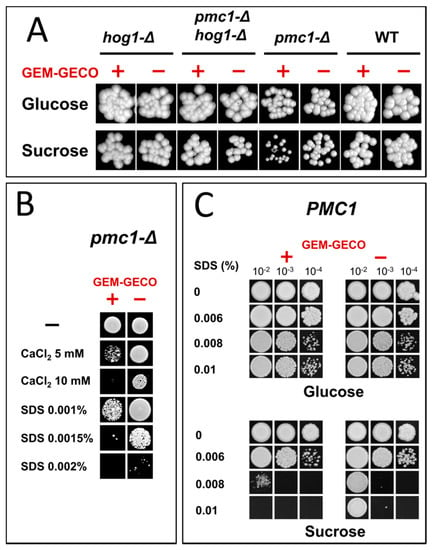
Figure 2.
Growth of strains differing in the expression of the GEM-GECO cassette on YPD (Glucose) and YP-Suc (Sucrose) solid media. (A), Spotting of 10−4-fold diluted YPD cultures on regular YPD and YP-Suc. WT, DL1-LC and DL5-LC strains; pmc1-Δ, DL5-pmc1-LC and DLdaduA-pmc1-LC strains; hog1-Δ, DL5-hog1 and DLdaduA-hog1-UC strains; pmc1-Δ hog1-Δ, DL5-pmc1-hog1 and DLdaduA-pmc1-hog1 strains. (B), Spotting of 10−4-fold diluted YPD cultures of DL-pmc1 (GEM-GECO −) and DL5-pmc1 (GEM-GECO +) on YPD with indicated supplements. (C), Spotting of 10−2-, 10−3-, and 10−4-fold diluted YPD cultures of DL1-L (GEM-GECO −) and DL5 (GEM-GECO +) on YPD and YP-Suc with indicated supplements.
2.2. GEM-GECO Does Not Complement the Loss of Endogenous CaM in Ogataea Yeast
How could the presence of GEM-GECO cause induction of pathways usually triggered by increased [Ca2+]cyt? We proposed that the CaM domain of this protein somehow participates in Ca2+ dependent signaling. While it was previously shown that GECIs possessing this domain can affect cell physiology by depleting Ca2+ from cytosol [12], we suggested that this could be due to functioning of the GEM-GECO calmodulin domain in Ca2+-dependent regulation same as the endogenous calmodulin. The gene encoding CaM in S. cerevisiae (CMD1) is essential for viability [18] and we expected that it would also indispensable in Ogataea yeast. To explore whether GEM-GECO can complement the absence of the endogenous CaM in O. parapolymorpha, the DL5 strain (leu2 PMAL1-GEM-GECO) was transformed with a CMD1 disruption cassette. PCR analysis of 24 transformants revealed integration of the disruption cassette into the CMD1 locus in two of them; however, the wild-type locus was also detected in all cases (Figure S1). This was likely due to locus duplication or aneuploidy. Absence of transformants lacking the wild type CMD1 gene strongly indicated that this gene is essential in O. parapolymorpha, as it is in S. cerevisiae. However, we were unable to test this via gene disruption in a O. parapolymorpha diploid strain since the DL-1 strain whose derivatives were used in this study, and commonly used in other laboratories, was an O. parapolymorpha asporogenous mutant [19]. To overcome this problem, we disrupted the CMD1 gene in a diploid strain of O. polymorpha, which is the closest relative of this yeast. This diploid strain was obtained by crossing strains with and without the GEM-GECO expression cassette. The transformants with the CMD1 disruption allele were incubated on sporulation medium and haploid segregants were selected using diethyl ether enrichment. Colonies of the segregants were obtained on both YPD and YP-Suc media, which provided repression or induction of GEM-GECO expression, respectively. None of the segregants possessed the disruption marker even though approximately half of them carried the expression cassette of GEM-GECO (Tables S1 and S2), which was expected to function as calmodulin. This proved that the CMD1 gene is indispensable in Ogataea yeasts and that GEM-GECO does not rescue the absence of this gene. We hypothesize that the vital pathway depending on endogenous calmodulin and the pathway that exerts Ca2+-dependent SDS sensitivity are different, and that the latter pathway can react to both the endogenous calmodulin and GEM-GECO.
2.3. GEM-GECO Allows Monitoring Ca2+ Concentration in Cytosol in O. parapolymorpha
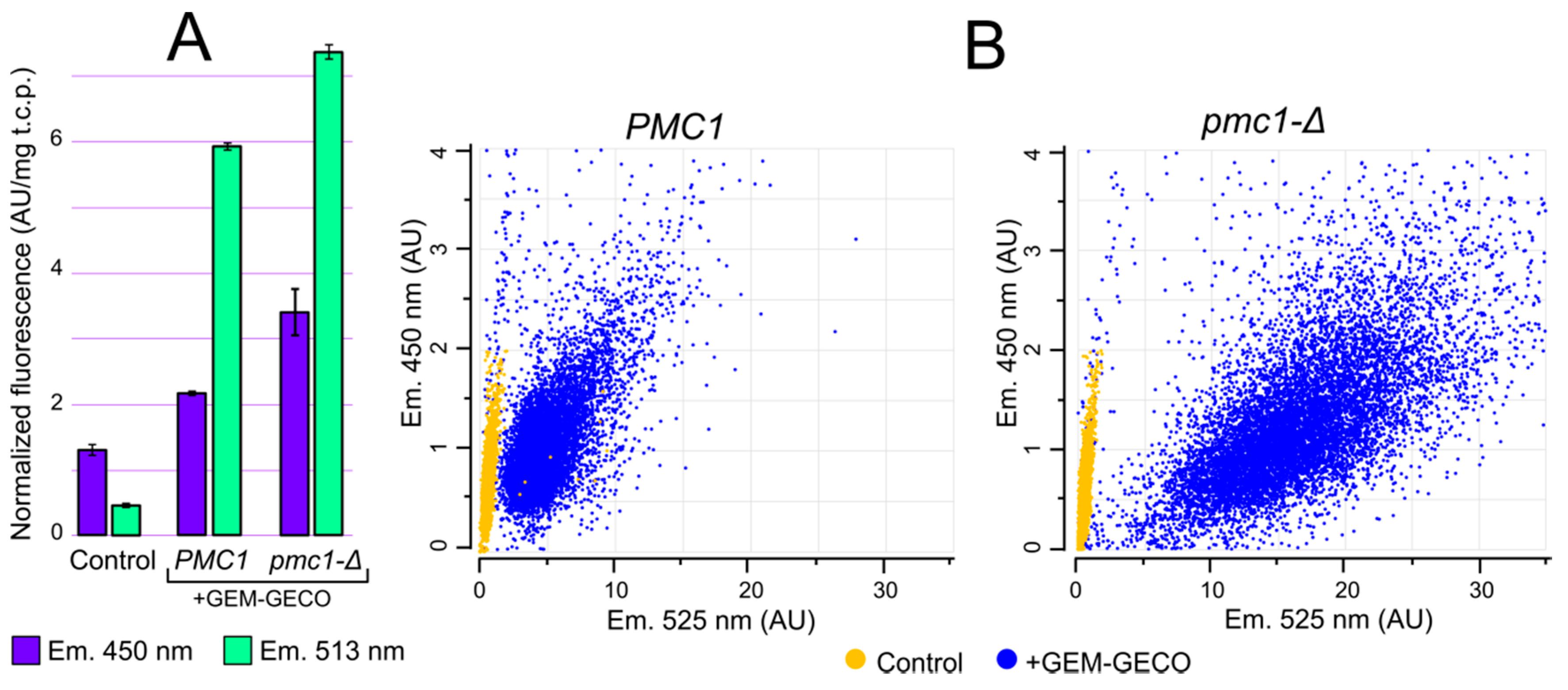
Although GEM-GECO expression affected cell growth in some cases, it did not cause cell death. This allowed us to study whether physiological changes in cytosolic Ca2+ concentration can be monitored by GEM-GECO in O. parapolymorpha. GEM-GECO fluorescence is excited at ~400 nm. The emission peak of the GEM-GECO Ca2+-free form is 513 nm, while its Ca2+-bound form emits with a peak at ~450 nm the quantum yield of the Ca2+-bound form at 450 nm is lower than that of Ca2+-free form at 513 nm [9]. Fluorescent microscopy analysis of cells expressing GEM-GECO revealed a cytosolic intracellular distribution pattern (Figure S2). We expected that inactivation of PMC1 could lead to an increase in [Ca2+]cyt that should increase fluorescence intensity at 450 nm and decrease it at 513 nm. Surprisingly we observed an increase in the fluorescence intensity in cell suspensions at both wavelengths in response to this mutation (Figure 3A). This could be due to higher GEM-GECO production level in the pmc1-Δ mutant.
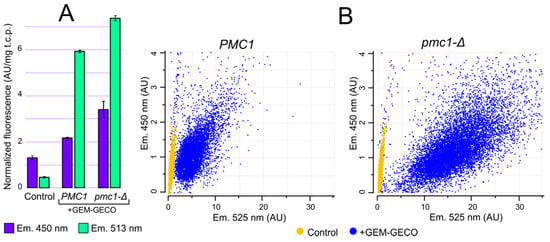
Figure 3.
Comparison of fluorescence characteristics of cells with (+GEM-GECO) and without (Control) the GEM-GECO expression cassette. (A), Fluorescence normalized to the total cell protein in the cell suspensions of DL1-LC (Control), DL5-LC (PMC1) and DL5-pmc1-LC (pmc1-Δ) strains. The fluorescence was excited by 395 nm light. (B), Flowcytometric analysis of cell fluorescence of strains: DL1-LC (Control, PMC1), DL1-pmc1-LC (Control, pmc1-Δ), DL5-LC (PMC1) and DL5-pmc1-LC (pmc1-Δ) strains (450 nm and 525 nm emission, 405 nm excitation).
To analyze GEM-GECO fluorescence in individual cells we used flowcytometry with a 405 nm excitation laser and fluorescence detection with 450 and 525 nm band-pass filters. Although the 450 nm fluorescence of majority of the GEM-GECO-expressing cells in regular conditions surpassed fluorescence of control cells lacking this expression cassette (Figure 3B), the autofluorescence at this wavelength was still considerable compared to the GEM-GECO signal. In contrast to 450 nm, the 525 nm fluorescence of GEM-GECO expressing cells strongly exceeded that of the control cells (Figure 3B). At the same time, when a considerable portion of GEM-GECO is in the Ca2+ bound form, which does not fluoresce at 525 nm, the auto fluorescence may noticeably distort the obtained results by overestimation of Ca2+-free form. To overcome this problem, calculated autofluorescence was subtracted from fluorescence intensities in individual cells (see Materials and Methods) and then the ratios between 450 nm and 525 nm fluorescence (FL450/FL525) were calculated as a characteristic of [Ca2+]cyt.
We suggested that a significant increase in external Ca2+ concentration would lead to some increase in [Ca2+]cyt. This would allow us to test applicability of GEM-GECO for monitoring [Ca2+]cyt changes in yeast cells. To demonstrate this effect with an alternative method we used the fluorescent dye Fluo4-AM, which is often used to detect increases in intracellular [Ca2+] [20]. There was no detectable difference in Fluo-4 fluorescence between the stained and unstained cells incubated in the regular medium indicating that amount of Fluo-4 Ca2+-bound form is below the detection level. Five-minute incubation in the medium containing 100 mM CaCl2 led to noticeable increase in the cell fluorescence, which could be detected by fluorescent microscopy (Figures S3). At the same time incubation of the Fluo-4-loaded cells in CaCl2-supplemented medium led to their flocculation since a significant portion of events detected by flow cytometry had very high forward scattering (Figure S4). Nevertheless, analysis of events whose forward scattering corresponds to non-flocculating cells also showed a noticeable increase in the fluorescence (Figure S4).
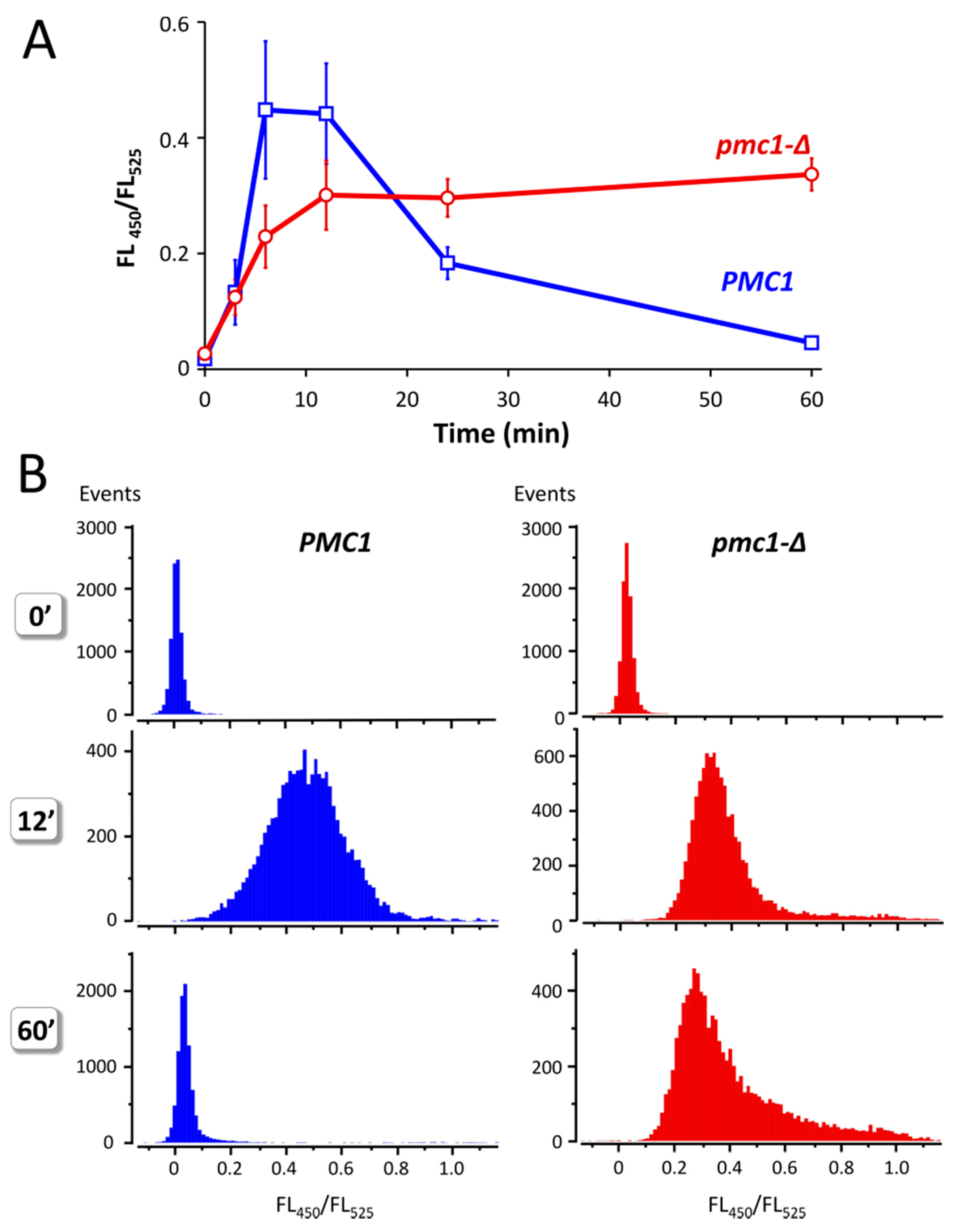
Yeast cells defective in the vacuolar Ca2+ ATPase Pmc1 are unable to grow at high Ca2+ concentration in culture medium [16,21]. This indicates that increased Ca2+ concentration in culture medium should lead to some increase in [Ca2+]cyt and that ability to resist it depends on Pmc1. Indeed, addition of 100 mM CaCl2 to the culture medium of strains expressing GEM-GECO led to a significant increase in FL450/FL525 in several minutes, which was followed by a decrease in this value during longer incubation in cells with wild-type PMC1 gene (Figure 4A). In the pmc1-Δ mutant, similar increase in FL450/FL525 was also observed; however, this was not followed by a decrease in this median value. Moreover, a noticeable portion of cells continued to accumulate Ca2+ according to the increasing FL450/FL525 (Figure 4B). Yet, the peak value of FL450/FL525 in the population was somewhat reduced, which indicates that a portion of cells retain some ability to reduce [Ca2+]cyt most likely due to action of the Vcx1 Ca2+/H+ exchanger. Notably, the FL450/FL525 value did not reach the original level after 1 h incubation even in the wild-type strain.
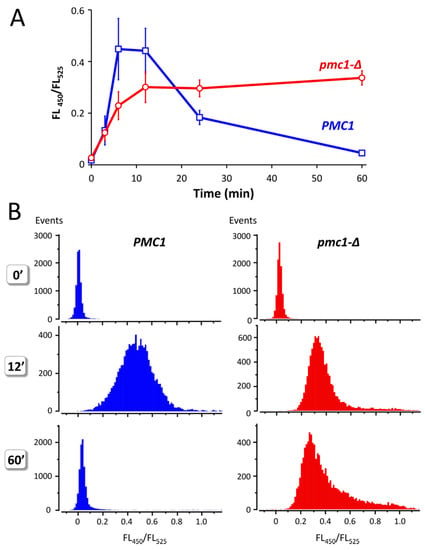
Figure 4.
Effect of Ca2+ concentration rise in culture medium on the ratio between fluorescence at 450 nm and 525 nm (FL450/FL525) excited by 405 nm laser in individual cells of strains DL5 (PMC1) and DL5-pmc1 (pmc1-Δ) producing GEM-GECO. Overnight YP-Suc cultures were 3-fold diluted with fresh YP-Suc medium, grown for 6 h. The obtained cultures were diluted 120-fold with YP-Suc medium supplemented with 100 mM CaCl2. Fluorescence in individual cells was measured using flowcytometry after 3′, 6′, 12′ and 60′ incubation. The 0′ samples are represented by the same cultures, which were diluted by YP-Suc without the CaCl2 supplement. (A), Dynamics of FL450/FL525 median values during incubation with 100 mM CaCl2 in the culture medium. Mean values of medians with standard deviations of these means were calculated using data from five and six independent replicates in the pmc1-Δ mutant and wild-type strain, respectively. (B), distribution of cells with noted FL450/FL525 values in cultures before (0′) and after 12′ and 60′ incubation in the culture medium supplemented with 100 mM CaCl2.
3. Materials and Methods
3.1. Yeast and Escherichia coli Strains
The O. parapolymorpha strains used in this study (Table 1) were constructed from the DL1-L strain (leu2) [22] or its derivative DLdaduA (leu2 ade2-Δ ura3::ADE2) [23]. The GEM-GECO expressing strain DL5 was obtained by transformation of DL1-L with the pKAM738 plasmid digested with BglII to facilitate its genome integration. The PMC1 gene was inactivated in the O. parapolymorpha genome by replacement with either the LEU2 gene as described in [16] or Cre recombinase recognition sequence loxP as described in [24]. EcoRI-digested empty vectors pCHLX [22] or pCCUR1 [23] possessing LEU2 or URA3 gene as a selectable marker, respectively, were integrated into a unidentified genome locus when complementing leu2 or ura3 auxotrophic mutations was required. The O. polymorpha diploid strain, in which the CMD1 gene was inactivated, was obtained by crossing of 1B27-740G1M strain (leu2 ade2 ura3::ADE2 {PMAL1:GEM-GECO}) possessing the GEM-GECO expression cassette integrated into unidentified locus with the 1B/SM/C2LX4 strain (leu2 ade2 mox::uPA [LEU2]) possessing an autonomously replicating plasmid with the LEU2 selectable marker. The 1B27-740G1M strain was obtained by transformation of the 1B27 strain [16] with the EcoRV-cleaved pAM740 GEM-GECO expression vector, whose LEU2-containing backbone was then excised by Cre-recombination. The 1B/SM strain was obtained from the 1B strain [25] by replacement of the MOX gene with the uPA expression cassette as described previously [26]. Then the obtained strain was transformed with an autonomously replicating plasmid pC2LX4. The diploid strain was selected on leucine, uracil and adenine omission medium. To obtain a leu2 auxotroph, which could be transformed with the CMD1 disruption cassette, the autonomously-replicating plasmid was lost from the obtained diploid. The E. coli DH-5α strain was used in plasmid construction procedures.

Table 1.
O. parapolymorpha strains used in this study. Names of plasmid genes integrated into unidentified genome locus as a part of a plasmid are shown in brackets.
3.2. Culture Media and Transformation Procedures
Complex media contained 2% Peptone, 1% Yeast extract and 2% glucose (YPD) or 1% sucrose (YP-Suc) as a carbon source. The synthetic medium SC-D (2% glucose, 0.67% Yeast Nitrogen Base with ammonium sulfate) was used for selection of transformants obtained with an auxotrophic selectable marker. The LB medium (1% Tryptone, 0.5% Yeast extract, 1% NaCl) was used to cultivate E. coli. E. coli was transformed with plasmids by method described in [27]. Yeast cells were transformed by Li-acetate method [28] with some modifications [29] as follows. Cells from 300 μL of an exponentially grown culture were harvested by centrifugation in a bench-top microcentrifuge (Eppendorf, Hamburg, Germany) at 5000 rpm for 30 sec, washed and re-suspended in 42 μL of sterile water. 2 μL of the DNA-carrier solution (10 mg/mL, shared and denatured by boiling) and 6 drops (approx. 90 μL) of 70% PEG 4000 were added and mixed well. After that the suspension was mixed with 9 ul of 1 M Li-acetate solution and dispensed by 20–23 μL to add 1 μL of transforming DNA to each portion. Suspensions were incubated first at 30 °C for 30 min than at 45 °C for 30 min. Cells were washed with YPD medium and spread on plates for selection of transformants.
3.3. Plasmid Construction
The DNA fragment with codon-optimized sequence encoding GEM-GECO was synthesized by Biomatik Corporation (Kitchener, ON, Canada). This fragment was fused with O. parapolymorpha MAL1 promoter sequence (EcoRV-SspI 579 bp fragment of PCR product obtained with primers AGGAATTAATATTGTCAAGAGGG and TGGACACGCGTGTGTCGAGAA) and inserted between PvuII and CfrI sites of the pKAM555 vector [30] to obtain pKAM738 plasmid. This vector possesses a selectable marker, which allows selection of transformants by kanamycin resistance in E. coli and G418 resistance in yeast. The pAM740 GEM-GECO expression vector was obtained by insertion of a 1.6 kb EcoRI-NheI fragment of pKAM738 between EcoRI-SpeI sites of the pAM631 self-excisable vector [24].
3.4. Analysis of GEM-GECO and Fluo-4 Fluorescence
GEM-GECO production in yeast transformants was induced by growing cells in the YP-Suc medium. Flow cytometric analysis of fluorescence (FL) in individual cells was performed using the CytoFLEX S flow cytometer (Beckman Coulter, Pasadena, CA, USA). The GEM-GECO fluorescence was excited by a 405 nm laser and measured using 450/20 nm (FL450) and 525/40 nm (FL525) bandpass filters. 10,000 cells were acquired in each sample. Cell fluorescence excited by 488 nm laser and measured using 525/40 nm bandpass filters (“FITC channel”) was found not to depend on presence of GEM-GECO. At the same time there was a good correlation between the “FITC channel” fluorescence and 405 nm-excited 525 nm and 450 nm autofluorescence (Figure S5). This allowed us to use the “FITC channel” fluorescence to estimate autofluorescence in GEM-GECO-expressing cells. To do this, trend lines of dependence of autofluorescences at 525 nm and 450 nm on “FITC channel” fluorescence in strains lacking GEM-GECO were calculated using MS Excel. Based on the obtained functions and “FITC channel” fluorescence values in the strains producing GEM-GECO, values of autofluorescence were calculated for individual cells and subtracted from the measured 525 nm and 450 nm fluorescence values. The obtained values were normalized to autofluorescence at each wavelength. The FL450/FL525 ratio was used as a characteristic of [Ca2+]cyt [9]. Fluorescence of cell suspensions was measured using an EnSpire plate reader (Perkin Elmer, Waltham, MA, USA). The obtained values were normalized to the total cell protein, which was assayed using the biuretic method described elsewhere [31].
Loading cells with Fluo-4 was performed according to the published protocol [32] with minor modifications as follows. Yeast cells were grown in YPD to OD600 between 1.0 and 2.0, washed twice with PBS (137 mM NaCl, 2.7 mM KCl, 8 mM Na2HPO4, and 2 mM KH2PO4) and resuspended in PBS to achieve the original cell density. The cell suspension was supplemented with 3 μM Fluo-4-AM (Thermo Fisher Scientific, Waltham, MA, USA) and incubated for 1 h at 37 °C. To test effect of external Ca2+ concentration rise, the cell suspension was diluted either with regular YPD, or YPD supplemented with 100 mM CaCl2. In the case of microscopic imaging, the cell suspension was loaded onto solid medium pads [33] made up of regular YPD, or YPD supplemented with 100 mM CaCl2.
4. Conclusions
In this work we have studied the applicability of the ratiometric GEM-GECO Ca2+ indicator for monitoring [Ca2+]cyt in yeast O. parapolymorpha. Production of this protein in cells with wild type Ca2+ homeostasis had a negligible effect on cell physiology. However, inactivation of the vacuolar Ca2+ ion pump Pmc1 manifested a pronounced interaction with GEM-GECO expression, which was revealed as aggravation of pmc1-Δ mutant phenotypes. Although previous studies indicated that physiological effects of such indicators in neuronal cells are related to Ca2+ depletion in cytosol, the effects we observed here were more likely to be related to an enhanced response to [Ca2+]cyt rise. We hypothesize that this is due to the CaM domain of GEM-GECO, which might interact with the host Ca2+-dependent machinery. Nevertheless, this indicator is applicable for monitoring [Ca2+]cyt in yeast cells. In particular, it is predominantly present in the Ca2+-free form in cells during normal conditions and converts to the Ca2+-bound form when [Ca2+]cyt rises in response to external Ca2+ concentration rise. This means that the [Ca2+]cyt in yeast cells is essentially below the Kd of the GEM-GECO-Ca2+ complex and can rise above this value in response to various treatments. We also observed that GEM-GECO expression under control of MAL1 promoter somehow depended on Ca2+ homeostasis, since it became elevated after inactivation of the PMC1 gene encoding the vacuolar Ca2+ ATPase. However, this did not affect [Ca2+]cyt monitoring since this indicator is ratiometric.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms231710004/s1.
Author Contributions
M.O.A. conceived and supervised the study; M.O.A., A.I.A. and A.V.K. designed experiments; M.O.A., A.V.K. and M.V.K. performed experiments; M.O.A. and A.I.A. wrote and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially supported by a grant from Russian Foundation for Basic Research (grant # 20-04-00330) (creation of the GECI construct, fluorescence quantification using cell suspension photometry and flow cytometry for time-resolved single-cell analysis of the changes in GECI fluorescence), a Grant for the Development of genomic editing technologies for innovation in industrial biotechnology # 075-15-2021-1071 from the Ministry of Science and Higher Education of the Russian Federation (introduction of GECI into various strains of O. polymorpha for determining the role of CMD1, cytometry data analysis and drafting and editing of the manuscript by A.I.A.), and base funding from the Ministry of Science and Higher Education of the Russian Federation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Acknowledgments
Flow cytometry was performed at the Shared-Access Equipment Centre “Industrial Biotechnology” of FRC “Fundamentals of Biotechnology” of the RAS. DNA sequencing was performed using equipment provided by the Shared-Access Equipment Centre “Bioengineering” of FRC “Fundamentals of Biotechnology” of the RAS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Eisner, D.A.; Caldwell, J.L.; Kistamás, K.; Trafford, A.W. Calcium and Excitation-Contraction Coupling in the Heart. Circ. Res. 2017, 121, 181–195. [Google Scholar] [CrossRef]
- Giorgi, C.; Danese, A.; Missiroli, S.; Patergnani, S.; Pinton, P. Calcium Dynamics as a Machine for Decoding Signals. Trends Cell Biol. 2018, 28, 258–273. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Poddar, A.; Sidibe, O.; Ray, A.; Chen, Q. Calcium Spikes Accompany Cleavage Furrow Ingression and Cell Separation during Fission Yeast Cytokinesis. Mol. Biol. Cell 2021, 32, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O.; Johnson, F.H. Calcium Binding, Quantum Yield, and Emitting Molecule in Aequorin Bioluminescence. Nature 1970, 227, 1356–1357. [Google Scholar] [CrossRef]
- Nakajima-Shimada, J.; Iida, H.; Tsuji, F.I.; Anraku, Y. Monitoring of Intracellular Calcium in Saccharomyces Cerevisiae with an Apoaequorin CDNA Expression System. Proc. Natl. Acad. Sci. USA 1991, 88, 6878–6882. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Nasu, Y.; Shkolnikov, I.; Kim, A.; Campbell, R.E. Engineering Genetically Encoded Fluorescent Indicators for Imaging of Neuronal Activity: Progress and Prospects. Neurosci. Res. 2020, 152, 3–14. [Google Scholar] [CrossRef]
- Mank, M.; Griesbeck, O. Genetically Encoded Calcium Indicators. Chem. Rev. 2008, 108, 1550–1564. [Google Scholar] [CrossRef]
- Zhao, Y.; Araki, S.; Wu, J.; Teramoto, T.; Chang, Y.-F.; Nakano, M.; Abdelfattah, A.S.; Fujiwara, M.; Ishihara, T.; Nagai, T.; et al. An Expanded Palette of Genetically Encoded Ca2+ Indicators. Science 2011, 333, 1888–1891. [Google Scholar] [CrossRef]
- Dana, H.; Mohar, B.; Sun, Y.; Narayan, S.; Gordus, A.; Hasseman, J.P.; Tsegaye, G.; Holt, G.T.; Hu, A.; Walpita, D.; et al. Sensitive Red Protein Calcium Indicators for Imaging Neural Activity. eLife 2016, 5, e12727. [Google Scholar] [CrossRef]
- Subach, O.M.; Barykina, N.V.; Chefanova, E.S.; Vlaskina, A.V.; Sotskov, V.P.; Ivashkina, O.I.; Anokhin, K.V.; Subach, F.V. FRCaMP, a Red Fluorescent Genetically Encoded Calcium Indicator Based on Calmodulin from Schizosaccharomyces Pombe Fungus. Int. J. Mol. Sci. 2020, 22, 111. [Google Scholar] [CrossRef]
- Gasterstädt, I.; Jack, A.; Stahlhut, T.; Rennau, L.-M.; Gonda, S.; Wahle, P. Genetically Encoded Calcium Indicators Can Impair Dendrite Growth of Cortical Neurons. Front. Cell. Neurosci. 2020, 14, 570596. [Google Scholar] [CrossRef]
- Liu, S.; Hou, Y.; Liu, W.; Lu, C.; Wang, W.; Sun, S. Components of the Calcium-Calcineurin Signaling Pathway in Fungal Cells and Their Potential as Antifungal Targets. Eukaryot. Cell 2015, 14, 324–334. [Google Scholar] [CrossRef]
- Cui, J.; Kaandorp, J.A.; Sloot, P.M.A.; Lloyd, C.M.; Filatov, M.V. Calcium Homeostasis and Signaling in Yeast Cells and Cardiac Myocytes. FEMS Yeast Res. 2009, 9, 1137–1147. [Google Scholar] [CrossRef]
- Yoshimoto, H.; Saltsman, K.; Gasch, A.P.; Li, H.X.; Ogawa, N.; Botstein, D.; Brown, P.O.; Cyert, M.S. Genome-Wide Analysis of Gene Expression Regulated by the Calcineurin/Crz1p Signaling Pathway in Saccharomyces Cerevisiae. J. Biol. Chem. 2002, 277, 31079–31088. [Google Scholar] [CrossRef]
- Fokina, A.V.; Sokolov, S.S.; Kang, H.A.; Kalebina, T.S.; Ter-Avanesyan, M.D.; Agaphonov, M.O. Inactivation of Pmc1 Vacuolar Ca2+ ATPase Causes G2 Cell Cycle Delay in Hansenula Polymorpha. Cell Cycle 2012, 11, 778–784. [Google Scholar] [CrossRef]
- Suppi, S.; Michelson, T.; Viigand, K.; Alamäe, T. Repression vs. Activation of MOX, FMD, MPP1 and MAL1 Promoters by Sugars in Hansenula Polymorpha: The Outcome Depends on Cell’s Ability to Phosphorylate Sugar. FEMS Yeast Res. 2013, 13, 219–232. [Google Scholar] [CrossRef]
- Davis, T.N.; Urdea, M.S.; Masiarz, F.R.; Thorner, J. Isolation of the Yeast Calmodulin Gene: Calmodulin Is an Essential Protein. Cell 1986, 47, 423–431. [Google Scholar] [CrossRef]
- Kurtzman, C.P. A New Methanol Assimilating Yeast, Ogataea Parapolymorpha, the Ascosporic State of Candida Parapolymorpha. Antonie van Leeuwenhoek 2011, 100, 455–462. [Google Scholar] [CrossRef]
- Gee, K.R.; Brown, K.A.; Chen, W.-N.U.; Bishop-Stewart, J.; Gray, D.; Johnson, I. Chemical and Physiological Characterization of Fluo-4 Ca2+-Indicator Dyes. Cell Calcium 2000, 27, 97–106. [Google Scholar] [CrossRef]
- Cunningham, K.W.; Fink, G.R. Calcineurin-Dependent Growth Control in Saccharomyces Cerevisiae Mutants Lacking PMC1, a Homolog of Plasma Membrane Ca2+ ATPases. J. Cell Biol. 1994, 124, 351–363. [Google Scholar] [CrossRef]
- Sohn, J.H.; Choi, E.S.; Kim, C.H.; Agaphonov, M.O.; Ter-Avanesyan, M.D.; Rhee, J.S.; Rhee, S.K. A Novel Autonomously Replicating Sequence (ARS) for Multiple Integration in the Yeast Hansenula Polymorpha DL-1. J. Bacteriol. 1996, 178, 4420–4428. [Google Scholar] [CrossRef]
- Karginov, A.V.; Fokina, A.V.; Kang, H.A.; Kalebina, T.S.; Sabirzyanova, T.A.; Ter-Avanesyan, M.D.; Agaphonov, M.O. Dissection of Differential Vanadate Sensitivity in Two Ogataea Species Links Protein Glycosylation and Phosphate Transport Regulation. Sci. Rep. 2018, 8, 16428. [Google Scholar] [CrossRef]
- Agaphonov, M.; Alexandrov, A. Self-Excising Integrative Yeast Plasmid Vectors Containing an Intronated Recombinase Gene. FEMS Yeast Res. 2014, 14, 1048–1054. [Google Scholar] [CrossRef]
- Bogdanova, A.I.; Agaphonov, M.O.; Ter-Avanesyan, M.D. Plasmid Reorganization during Integrative Transformation InHansenula Polymorpha. Yeast 1995, 11, 343–353. [Google Scholar] [CrossRef]
- Agaphonov, M.O.; Beburov, M.Y.; Ter-Avanesyan, M.D.; Smirnov, V.N. A Disruption-Replacement Approach for the Targeted Integration of Foreign Genes in Hansenula Polymorpha. Yeast 1995, 11, 1241–1247. [Google Scholar] [CrossRef]
- Inoue, H.; Nojima, H.; Okayama, H. High Efficiency Transformation of Escherichia Coli with Plasmids. Gene 1990, 96, 23–28. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. Quick and Easy Yeast Transformation Using the LiAc/SS Carrier DNA/PEG Method. Nat. Protoc. 2007, 2, 35–37. [Google Scholar] [CrossRef]
- Karginov, A.V.; Alexandrov, A.I.; Kushnirov, V.V.; Agaphonov, M.O. Perturbations in the Heme and Siroheme Biosynthesis Pathways Causing Accumulation of Fluorescent Free Base Porphyrins and Auxotrophy in Ogataea Yeasts. J. Fungi 2021, 7, 884. [Google Scholar] [CrossRef]
- Agaphonov, M.; Romanova, N.; Choi, E.-S.; Ter-Avanesyan, M. A Novel Kanamycin/G418 Resistance Marker for Direct Selection of Transformants in Escherichia Coli and Different Yeast Species. Yeast 2010, 27, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Herbert, D.; Phipps, P.J.; Strange, R.E. Chapter III Chemical Analysis of Microbial Cells. In Methods in Microbiology; Academic Press: Cambridge, MA, USA, 1971; pp. 209–344. [Google Scholar]
- Varela, J.N.; Yadav, V.G. A Pichia Biosensor for High-throughput Analyses of Compounds That Can Influence Mosquito Behavior. Microbiologyopen 2021, 10, e1139. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, A.I.; Dergalev, A.A. Increasing Throughput of Manual Microscopy of Cell Suspensions Using Solid Medium Pads. MethodsX 2019, 6, 329–332. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).