
Reversible Photocontrol of Dopaminergic Transmission in Wild-Type Animals

 ,
,  , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
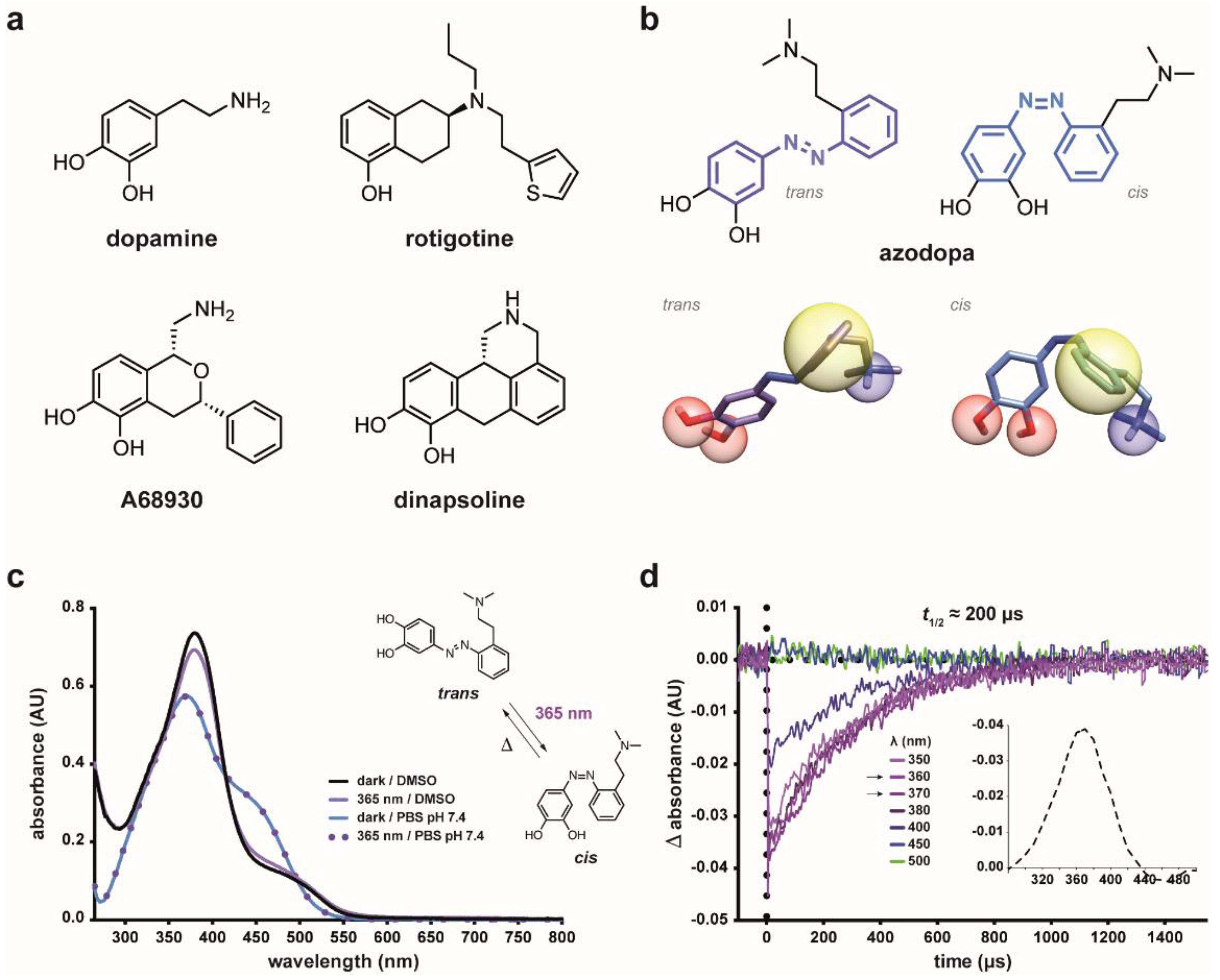
2.1. Design Strategy and Synthesis
2.2. Photochemical Characterization
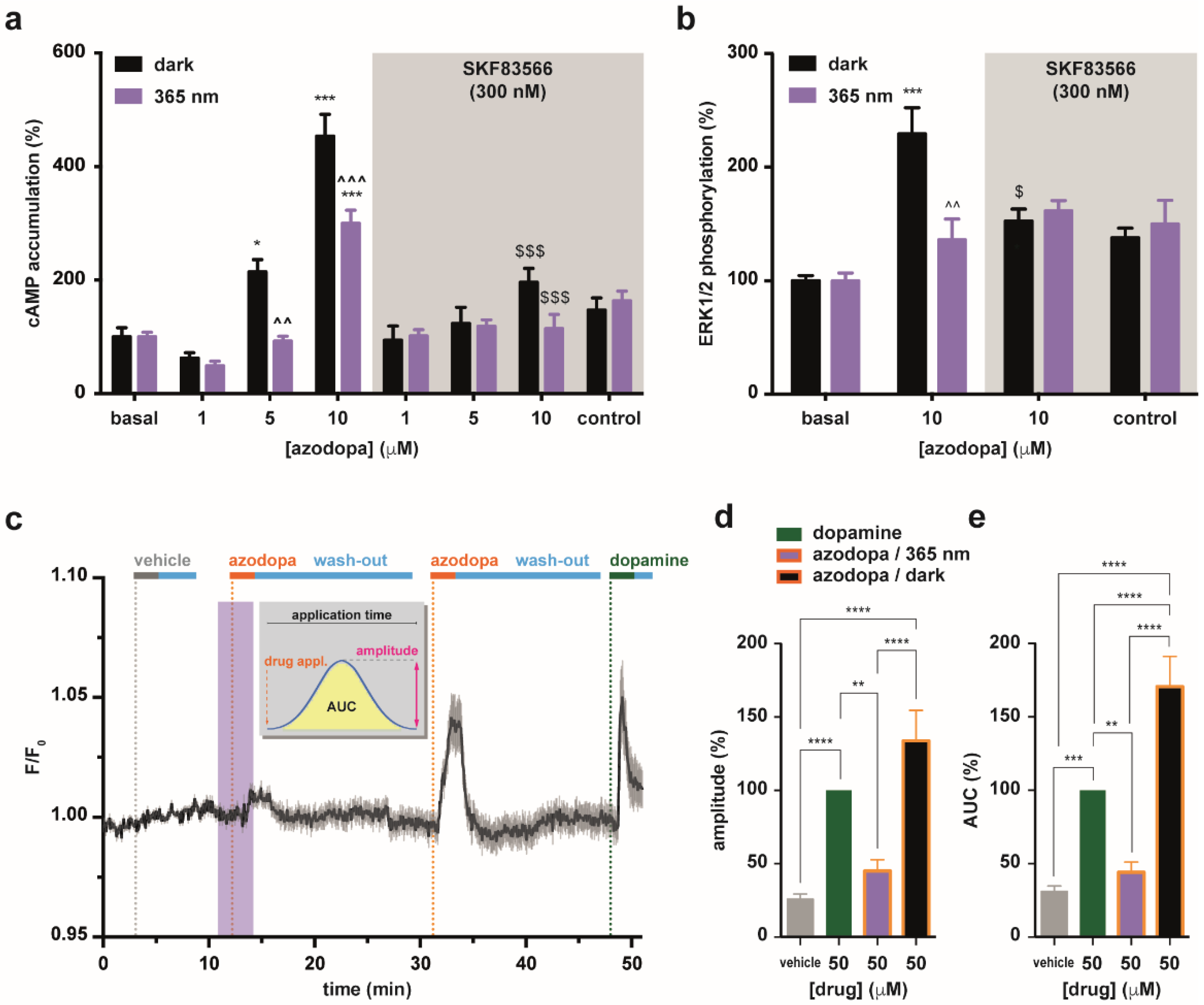
2.3. In Vitro Pharmacology
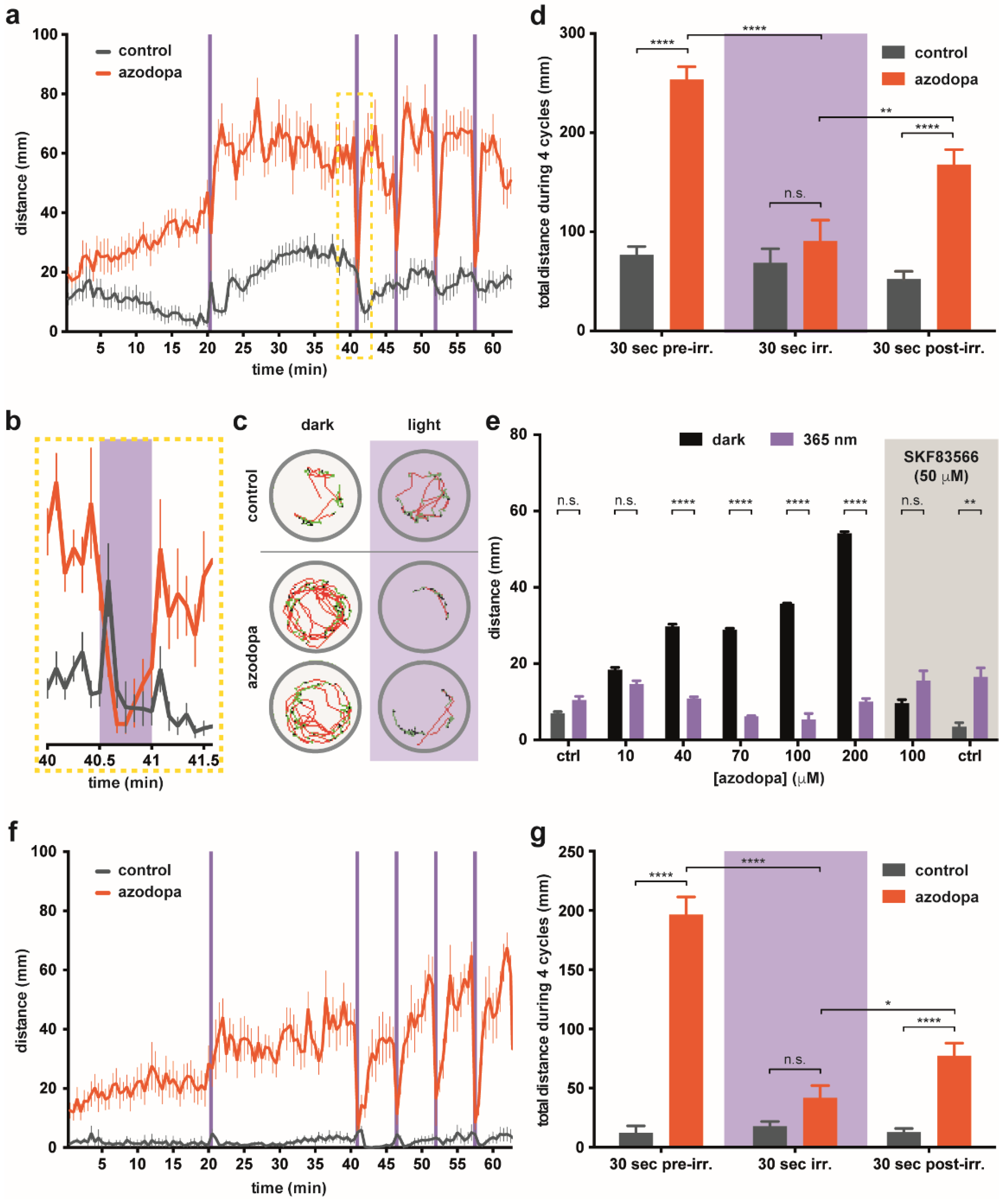
2.4. Behavioral Effects in Zebrafish Larvae
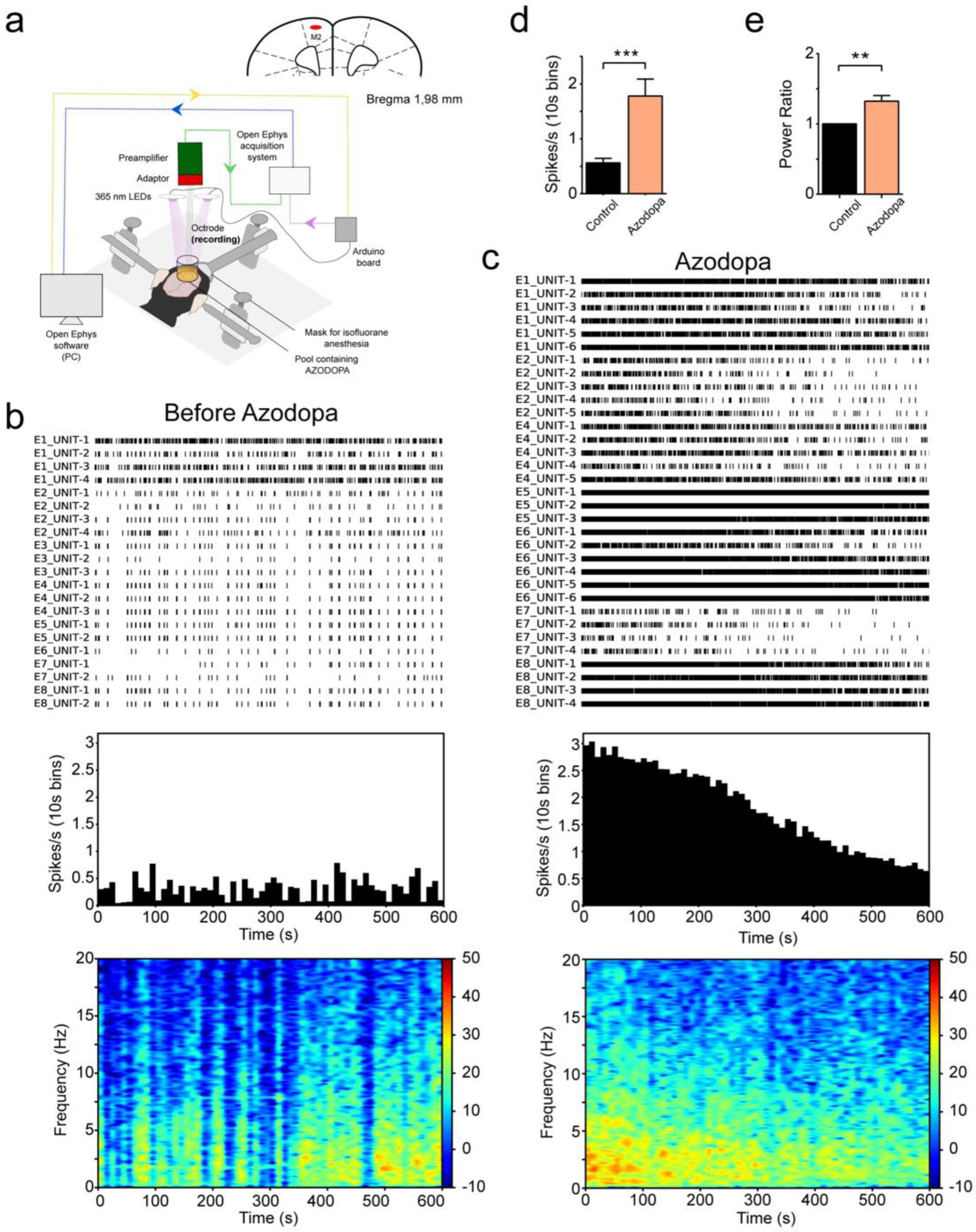
2.5. Electrophysiological Recordings in Anesthetized Mice
3. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beaulieu, J.-M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef] [PubMed]
- Tritsch, N.X.; Sabatini, B.L. Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron 2012, 76, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Firsov, M.L.; Astakhova, L.A. The Role of Dopamine in Controlling Retinal Photoreceptor Function in Vertebrates. Neurosci. Behav. Phys. 2015, 46, 138–145. [Google Scholar] [CrossRef]
- Korshunov, K.S.; Blakemore, L.J.; Trombley, P.Q. Dopamine: A Modulator of Circadian Rhythms in the Central Nervous System. Front. Cell. Neurosci. 2017, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.V.; Miller, E.K. The role of prefrontal dopamine D1 receptors in the neural mechanisms of associative learning. Neuron 2012, 74, 874–886. [Google Scholar] [CrossRef]
- Puig, M.V.; Miller, E.K. Neural Substrates of Dopamine D2 Receptor Modulated Executive Functions in the Monkey Prefrontal Cortex. Cereb. Cortex 2015, 25, 2980–2987. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.-M.; Espinoza, S.; Gainetdinov, R.R. Dopamine receptors—IUPHAR Review 13. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef]
- Lerch, M.M.; Hansen, M.J.; van Dam, G.M.; Szymanski, W.; Feringa, B.L. Emerging Targets in Photopharmacology. Angew. Chem. Int. Ed. Engl. 2016, 55, 10978–10999. [Google Scholar] [CrossRef] [PubMed]
- Hüll, K.; Morstein, J.; Trauner, D. In Vivo Photopharmacology. Chem. Rev. 2018, 118, 10710–10747. [Google Scholar] [CrossRef]
- Pianowski, Z.L. Molecular Photoswitches: Synthesis, Properties, and Applications; VCH: Hoboken, NJ, USA, 2021. [Google Scholar]
- Pittolo, S.; Gómez-Santacana, X.; Eckelt, K.; Rovira, X.; Dalton, J.; Goudet, C.; Pin, J.-P.; Llobet, A.; Giraldo, J.; Llebaria, A.; et al. An allosteric modulator to control endogenous G protein-coupled receptors with light. Nat. Chem. Biol. 2014, 10, 813–815. [Google Scholar] [CrossRef]
- Agnetta, L.; Bermudez, M.; Riefolo, F.; Matera, C.; Claro, E.; Messerer, R.; Littmann, T.; Wolber, G.; Holzgrabe, U.; Decker, M. Fluorination of Photoswitchable Muscarinic Agonists Tunes Receptor Pharmacology and Photochromic Properties. J. Med. Chem. 2019, 62, 3009–3020. [Google Scholar] [CrossRef] [PubMed]
- Riefolo, F.; Matera, C.; Garrido-Charles, A.; Gomila, A.M.J.; Sortino, R.; Agnetta, L.; Claro, E.; Masgrau, R.; Holzgrabe, U.; Batlle, M.; et al. Optical Control of Cardiac Function with a Photoswitchable Muscarinic Agonist. J. Am. Chem. Soc. 2019, 141, 7628–7636. [Google Scholar] [CrossRef] [PubMed]
- Prischich, D.; Gomila, A.M.J.; Milla-Navarro, S.; Sangüesa, G.; Diez-Alarcia, R.; Preda, B.; Matera, C.; Batlle, M.; Ramírez, L.; Giralt, E.; et al. Adrenergic Modulation with Photochromic Ligands. Angew. Chem. Int. Ed. Engl. 2021, 60, 3625–3631. [Google Scholar] [CrossRef] [PubMed]
- Donthamsetti, P.; Winter, N.; Hoagland, A.; Stanley, C.; Visel, M.; Lammel, S.; Trauner, D.; Isacoff, E. Cell specific photoswitchable agonist for reversible control of endogenous dopamine receptors. Nat. Commun. 2021, 12, 4775. [Google Scholar] [CrossRef] [PubMed]
- Berizzi, A.E.; Goudet, C. Strategies and considerations of G-protein-coupled receptor photopharmacology. Adv. Pharm. 2020, 88, 143–172. [Google Scholar]
- Bregestovski, P.; Maleeva, G.; Gorostiza, P. Light-induced regulation of ligand-gated channel activity. Br. J. Pharmacol. 2017, 172, 5870. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Serra, M.; Bautista-Barrufet, A.; Trapero, A.; Garrido-Charles, A.; Díaz-Tahoces, A.; Camarero, N.; Pittolo, S.; Valbuena, S.; Pérez-Jiménez, A.; Gay, M.; et al. Optical control of endogenous receptors and cellular excitability using targeted covalent photoswitches. Nat. Commun. 2016, 7, 12221. [Google Scholar] [CrossRef]
- Szymanski, W.; Ourailidou, M.E.; Velema, W.A.; Dekker, F.J.; Feringa, B.L. Light-Controlled Histone Deacetylase (HDAC) Inhibitors: Towards Photopharmacological Chemotherapy. Chem. Eur. J. 2015, 21, 16517–16524. [Google Scholar] [CrossRef]
- Matera, C.; Gomila, A.M.J.; Camarero, N.; Libergoli, M.; Soler, C.; Gorostiza, P. Photoswitchable Antimetabolite for Targeted Photoactivated Chemotherapy. J. Am. Chem. Soc. 2018, 140, 15764–15773. [Google Scholar] [CrossRef]
- Steinberg, E.E.; Janak, P.H. Establishing causality for dopamine in neural function and behavior with optogenetics. Brain Res. 2013, 1511, 46–64. [Google Scholar] [CrossRef]
- Stauffer, W.R.; Lak, A.; Yang, A.; Borel, M.; Paulsen, O.; Boyden, E.S.; Schultz, W. Dopamine Neuron-Specific Optogenetic Stimulation in Rhesus Macaques. Cell 2016, 166, 1564–1571.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.Y.; Esber, G.R.; Marrero-Garcia, Y.; Yau, H.-J.; Bonci, A.; Schoenbaum, G. Brief optogenetic inhibition of dopamine neurons mimics endogenous negative reward prediction errors. Nat. Neurosci. 2016, 19, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Hare, B.D.; Shinohara, R.; Liu, R.J.; Pothula, S.; DiLeone, R.J.; Duman, R.S. Optogenetic stimulation of medial prefrontal cortex Drd1 neurons produces rapid and long-lasting antidepressant effects. Nat. Commun. 2019, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Donthamsetti, P.C.; Winter, N.; Schönberger, M.; Levitz, J.; Stanley, C.; Javitch, J.A.; Isacoff, E.Y.; Trauner, D. Optical Control of Dopamine Receptors Using a Photoswitchable Tethered Inverse Agonist. J. Am. Chem. Soc. 2017, 139, 18522–18535. [Google Scholar] [CrossRef] [PubMed]
- Araya, R.; Andino-Pavlovsky, V.; Yuste, R.; Etchenique, R. Two-photon optical interrogation of individual dendritic spines with caged dopamine. ACS Chem. Neurosci. 2013, 4, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Gienger, M.; Hübner, H.; Löber, S.; König, B.; Gmeiner, P. Structure-based development of caged dopamine D2/D3 receptor antagonists. Sci. Rep. 2020, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, D.; Studte, C.; Männel, B.; Hübner, H.; Gmeiner, P.; König, B. Photochromic Dopamine Receptor Ligands Based on Dithienylethenes and Fulgides. Chem. Eur. J. 2017, 23, 13423–13434. [Google Scholar] [CrossRef]
- Szymanski, W.; Beierle, J.M.; Kistemaker, H.A.V.; Velema, W.A.; Feringa, B.L. Reversible photocontrol of biological systems by the incorporation of molecular photoswitches. Chem. Rev. 2013, 113, 6114–6178. [Google Scholar] [CrossRef]
- Broichhagen, J.; Frank, J.A.; Trauner, D. A Roadmap to Success in Photopharmacology. Acc. Chem. Res. 2015, 48, 1947–1960. [Google Scholar] [CrossRef]
- Schoenberger, M.; Damijonaitis, A.; Zhang, Z.; Nagel, D.; Trauner, D. Development of a new photochromic ion channel blocker via azologization of fomocaine. ACS Chem. Neurosci. 2014, 5, 514–518. [Google Scholar] [CrossRef]
- Borowiak, M.; Nahaboo, W.; Reynders, M.; Nekolla, K.; Jalinot, P.; Hasserodt, J.; Rehberg, M.; Delattre, M.; Zahler, S.; Vollmar, A.; et al. Photoswitchable Inhibitors of Microtubule Dynamics Optically Control Mitosis and Cell Death. Cell 2015, 162, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morstein, J.; Awale, M.; Reymond, J.-L.; Trauner, D. Mapping the Azolog Space Enables the Optical Control of New Biological Targets. ACS Cent. Sci. 2019, 5, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, C.; Bertolini, G. Perspectives in the design and application of dopamine receptor agonists. In Perspectives in Receptor Research; Elsevier: Amsterdam, The Netherlands, 1996; Volume 24, pp. 67–84. [Google Scholar]
- Rodenhuis, N. New, Centrally Acting Dopaminergic Agents with an Improved Oral Bioavailability: Synthesis and Pharmacological Evaluation. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2000. [Google Scholar]
- Pettersson, I.; Liljefors, T. Structure-activity relationships for apomorphine congeners. Conformational energies vs. biological activities. J. Comput. Aided Mol. Des. 1987, 1, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.G.; Borgman, R.J.; Aleem, M.A.; Long, J.P. Centrally acting emetics. 7. Hofmann and Emde degradation products of apomorphine. J. Med. Chem. 1973, 16, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Malo, M.; Brive, L.; Luthman, K.; Svensson, P. Investigation of D2 receptor-agonist interactions using a combination of pharmacophore and receptor homology modeling. ChemMedChem 2012, 7, 471–82–338. [Google Scholar] [CrossRef] [PubMed]
- Malo, M.; Brive, L.; Luthman, K.; Svensson, P. Investigation of D1 receptor-agonist interactions and D1/D2 agonist selectivity using a combination of pharmacophore and receptor homology modeling. ChemMedChem 2012, 7, 483–94–338. [Google Scholar] [CrossRef] [PubMed]
- Kołaczkowski, M.; Bucki, A.; Feder, M.; Pawłowski, M. Ligand-optimized homology models of D1 and D2 dopamine receptors: Application for virtual screening. J. Chem. Inf. Modeling 2013, 53, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Amorós, J.; Sánchez-Ferrer, A.; Massad, W.A.; Nonell, S.; Velasco, D. Kinetic study of the fast thermal cis-to-trans isomerisation of para-, ortho- and polyhydroxyazobenzenes. Phys. Chem. Chem. Phys. 2010, 12, 13238–13242. [Google Scholar] [CrossRef]
- Izquierdo-Serra, M.; Gascón-Moya, M.; Hirtz, J.J.; Pittolo, S.; Poskanzer, K.E.; Ferrer, È.; Alibés, R.; Busqué, F.; Yuste, R.; Hernando, J.; et al. Two-photon neuronal and astrocytic stimulation with azobenzene-based photoswitches. J. Am. Chem. Soc. 2014, 136, 8693–8701. [Google Scholar] [CrossRef]
- Ferré, S.; Casadó, V.; Devi, L.A.; Filizola, M.; Jockers, R.; Lohse, M.J.; Milligan, G.; Pin, J.-P.; Guitart, X. G protein-coupled receptor oligomerization revisited: Functional and pharmacological perspectives. Pharmacol. Rev. 2014, 66, 413–434. [Google Scholar] [CrossRef]
- Casadó, V.; Ferrada, C.; Bonaventura, J.; Gracia, E.; Mallol, J.; Canela, E.I.; Lluis, C.; Cortés, A.; Franco, R. Useful pharmacological parameters for G-protein-coupled receptor homodimers obtained from competition experiments. Agonist-antagonist binding modulation. Biochem. Pharmacol. 2009, 78, 1456–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.-Q.; Wang, H.-Y.; Friedman, E. Stimulated D1 dopamine receptors couple to multiple Gα proteins in different brain regions. J. Neurochem. 2001, 78, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Chiken, S.; Sato, A.; Ohta, C.; Kurokawa, M.; Arai, S.; Maeshima, J.; Sunayama-Morita, T.; Sasaoka, T.; Nambu, A. Dopamine D1 Receptor-Mediated Transmission Maintains Information Flow through the Cortico-Striato-Entopeduncular Direct Pathway to Release Movements. Cereb. Cortex 2015, 25, 4885–4897. [Google Scholar] [CrossRef] [PubMed]
- Basnet, R.M.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish Larvae as a Behavioral Model in Neuropharmacology. Biomedicines 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Norton, W.; Coolen, M.; Chaminade, M.; Merker, S.; Proft, F.; Schmitt, A.; Vernier, P.; Lesch, K.-P.; Bally-Cuif, L. The ADHD-susceptibility gene lphn3.1 modulates dopaminergic neuron formation and locomotor activity during zebrafish development. Mol. Psychiatry 2012, 17, 946–954. [Google Scholar] [CrossRef]
- Ek, F.; Malo, M.; Åberg Andersson, M.; Wedding, C.; Kronborg, J.; Svensson, P.; Waters, S.; Petersson, P.; Olsson, R. Behavioral Analysis of Dopaminergic Activation in Zebrafish and Rats Reveals Similar Phenotypes. ACS Chem. Neurosci. 2016, 7, 633–646. [Google Scholar] [CrossRef]
- Levitas-Djerbi, T.; Appelbaum, L. Modeling sleep and neuropsychiatric disorders in zebrafish. Curr. Opin. Neurobiol. 2017, 44, 89–93. [Google Scholar] [CrossRef]
- Irons, T.D.; Kelly, P.E.; Hunter, D.L.; Macphail, R.C.; Padilla, S. Acute administration of dopaminergic drugs has differential effects on locomotion in larval zebrafish. Pharmacol. Biochem. Behav. 2013, 103, 792–813. [Google Scholar] [CrossRef]
- Ingebretson, J.J.; Masino, M.A. Quantification of locomotor activity in larval zebrafish: Considerations for the design of high-throughput behavioral studies. Front. Neural Circuits 2013, 7, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, P.A.; Barthel, L.K.; Curran, G.A. Developmental patterning of rod and cone photoreceptors in embryonic zebrafish. J. Comp. Neurol. 1995, 359, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Guggiana-Nilo, D.A.; Engert, F. Properties of the Visible Light Phototaxis and UV Avoidance Behaviors in the Larval Zebrafish. Front. Behav. Neurosci. 2016, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Chen, J.; Luo, J.; Hitchcock, P. Light-induced photoreceptor degeneration in the retina of the zebrafish. Methods Mol. Biol. 2012, 884, 247–254. [Google Scholar] [PubMed]
- Kaufman, D.M.; Milstein, M.J. Neurotransmitters and Drug Abuse. In Kaufman’s Clinical Neurology for Psychiatrists; Elsevier: Amsterdam, The Netherlands, 2013; pp. 501–525. [Google Scholar]
- Roffman, J.L.; Tanner, A.S.; Eryilmaz, H.; Rodriguez-Thompson, A.; Silverstein, N.J.; Ho, N.F.; Nitenson, A.Z.; Chonde, D.B.; Greve, D.N.; Abi-Dargham, A.; et al. Dopamine D1 signaling organizes network dynamics underlying working memory. Sci. Adv. 2016, 2, e1501672. [Google Scholar] [CrossRef]
- Nieh, E.H.; Kim, S.-Y.; Namburi, P.; Tye, K.M. Optogenetic dissection of neural circuits underlying emotional valence and motivated behaviors. Brain Res. 2013, 1511, 73–92. [Google Scholar] [CrossRef]
- Roy, S.; Field, G.D. Dopaminergic modulation of retinal processing from starlight to sunlight. J. Pharm. Sci. 2019, 140, 86–93. [Google Scholar] [CrossRef]
- Alemany-González, M.; Gener, T.; Nebot, P.; Vilademunt, M.; Dierssen, M.; Puig, M.V. Prefrontal-hippocampal functional connectivity encodes recognition memory and is impaired in intellectual disability. Proc. Natl. Acad. Sci. USA 2020, 117, 11788–11798. [Google Scholar] [CrossRef]
- Aubert, I.; Guigoni, C.; Håkansson, K.; Li, Q.; Dovero, S.; Barthe, N.; Bioulac, B.H.; Gross, C.E.; Fisone, G.; Bloch, B.; et al. Increased D1 dopamine receptor signaling in levodopa-induced dyskinesia. Ann. Neurol. 2005, 57, 17–26. [Google Scholar] [CrossRef]
- Cabré, G.; Garrido-Charles, A.; González-Lafont, À.; Moormann, W.; Langbehn, D.; Egea, D.; Lluch, J.M.; Herges, R.; Alibés, R.; Busqué, F.; et al. Synthetic Photoswitchable Neurotransmitters Based on Bridged Azobenzenes. Org. Lett. 2019, 21, 3780–3784. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Charles, A.; Huet, A.; Matera, C.; Thirumalai, A.; Llebaria, A.; Moser, T.; Gorostiza, P. Fast photoswitchable molecular prosthetics control neuronal activity in the cochlea. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Pittolo, S.; Lee, H.; Lladó, A.; Tosi, S.; Bosch, M.; Bardia, L.; Gómez-Santacana, X.; Llebaria, A.; Soriano, E.; Colombelli, J.; et al. Reversible silencing of endogenous receptors in intact brain tissue using 2-photon pharmacology. Proc. Natl. Acad. Sci. USA 2019, 116, 13680–13689. [Google Scholar] [CrossRef] [PubMed]
- Cabré, G.; Garrido-Charles, A.; Moreno, M.; Bosch, M.; Porta-de-la-Riva, M.; Krieg, M.; Gascón-Moya, M.; Camarero, N.; Gelabert, R.; Lluch, J.M.; et al. Rationally designed azobenzene photoswitches for efficient two-photon neuronal excitation. Nat. Commun. 2019, 10, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Casadó, V.; Cortés, A.; Ciruela, F.; Mallol, J.; Ferré, S.; Lluis, C.; Canela, E.I.; Franco, R. Old and new ways to calculate the affinity of agonists and antagonists interacting with G-protein-coupled monomeric and dimeric receptors: The receptor-dimer cooperativity index. Pharmacol. Ther. 2007, 116, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Ferrada, C.; Moreno, E.; Casadó, V.; Bongers, G.; Cortés, A.; Mallol, J.; Canela, E.I.; Leurs, R.; Ferré, S.; Lluís, C.; et al. Marked changes in signal transduction upon heteromerization of dopamine D1 and histamine H3 receptors. Br. J. Pharmacol. 2009, 157, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matera, C.; Calvé, P.; Casadó-Anguera, V.; Sortino, R.; Gomila, A.M.J.; Moreno, E.; Gener, T.; Delgado-Sallent, C.; Nebot, P.; Costazza, D.; et al. Reversible Photocontrol of Dopaminergic Transmission in Wild-Type Animals. Int. J. Mol. Sci. 2022, 23, 10114. https://doi.org/10.3390/ijms231710114
Matera C, Calvé P, Casadó-Anguera V, Sortino R, Gomila AMJ, Moreno E, Gener T, Delgado-Sallent C, Nebot P, Costazza D, et al. Reversible Photocontrol of Dopaminergic Transmission in Wild-Type Animals. International Journal of Molecular Sciences. 2022; 23(17):10114. https://doi.org/10.3390/ijms231710114
Chicago/Turabian StyleMatera, Carlo, Pablo Calvé, Verònica Casadó-Anguera, Rosalba Sortino, Alexandre M. J. Gomila, Estefanía Moreno, Thomas Gener, Cristina Delgado-Sallent, Pau Nebot, Davide Costazza, and et al. 2022. "Reversible Photocontrol of Dopaminergic Transmission in Wild-Type Animals" International Journal of Molecular Sciences 23, no. 17: 10114. https://doi.org/10.3390/ijms231710114
APA StyleMatera, C., Calvé, P., Casadó-Anguera, V., Sortino, R., Gomila, A. M. J., Moreno, E., Gener, T., Delgado-Sallent, C., Nebot, P., Costazza, D., Conde-Berriozabal, S., Masana, M., Hernando, J., Casadó, V., Puig, M. V., & Gorostiza, P. (2022). Reversible Photocontrol of Dopaminergic Transmission in Wild-Type Animals. International Journal of Molecular Sciences, 23(17), 10114. https://doi.org/10.3390/ijms231710114