Abstract
Parkinson’s disease (PD) is a neurodegenerative disease characterized by the accumulation of alpha-synuclein, encoded by the SNCA gene. The main neuropathological hallmark of PD is the degeneration of dopaminergic neurons leading to striatal dopamine depletion. Trophic support by a neurotrophin called glial-derived neurotrophic factor (GDNF) is also lacking in PD. We performed immunohistochemical studies to investigate neuropathological changes in the basal ganglia of a rat transgenic model of PD overexpressing alfa-synuclein. We observed that neuronal loss also occurs in the dorsolateral part of the striatum in the advanced stages of the disease. Moreover, along with the degeneration of the medium spiny projection neurons, we found a dramatic loss of parvalbumin interneurons. A marked decrease in GDNF, which is produced by parvalbumin interneurons, was observed in the striatum and in the substantia nigra of these animals. This confirmed the involvement of the striatum in the pathophysiology of PD and the importance of GDNF in maintaining the health of the substantia nigra.
1. Introduction
Parkinson’s disease (PD) is a neurodegenerative disease characterized by the accumulation of alpha-synuclein, encoded by the SNCA gene [1]. The well-known PD triad of symptoms is characterized by tremor, rigidity, and bradykinesia, and is linked to neuronal loss in the substantia nigra pars compacta. Indeed, the pathognomonic feature of PD is the degeneration of these dopaminergic neurons leading to striatal dopamine depletion. Another essential feature of PD neuropathology is the Lewy pathology, consisting of the presence of Lewy bodies and Lewy neurites [2,3] in neurons.
PD is also characterized by cognitive disturbances such as executive functions impairment, language deficits, visuospatial/visuoconstructive disturbance, and impulse control disorders [4,5,6,7,8]. These non-motor symptoms have been associated with a disruption of frontal-subcortical networks [9]. Moreover, mood disorders often occur in PD patients and are thought to derive from extra-nigral pathology. The latter could constitute the reason for the resistance of some cases to pharmacological dopaminergic treatment [10,11,12,13,14].
In 1993, Lin and coworkers described a specific DA neurotrophic factor that is produced by glial cells in rats, namely, the glial cell line-derived neurotrophic factor (GDNF) [15]. This discovery opened new possibilities for the pathogenesis and therapy of PD.
The survival of adult nigrostriatal dopaminergic neurons is strictly dependent on striatal production of GDNF. GDNF is a trophic factor that has been proven beneficial for nigrostriatal neurons, as it displayed significant neurotrophic effects on mesencephalic dopaminergic neurons as well as on motor neurons [16,17,18,19].
It has recently been demonstrated that the population of parvalbumin containing GABAergic interneurons is responsible for the production of most of the striatal GDNF [19]. Indeed, the authors showed that approximately 95% of GDNF-expressing cells in the striatum are PV-positive interneurons, whereas the vast majority of striatal neurons that receive a strong dopaminergic innervation, namely the projection medium spiny neurons, are devoid of GDNF.
PV interneurons are represented throughout the striatum, and they are distributed in a ventral-to-dorsal, medial-to-lateral, and caudal-to-rostral gradient of increasing density. Their distribution coincides with the area innervated by dopaminergic terminals from the substantia nigra [20]. PV interneurons receive dopaminergic inputs from the substantia nigra and glutamatergic synapses from thalamus and cortex. Their axons generate proximal inhibitory synapses onto medium spiny neurons. As in other areas of the central nervous system, (i.e., cortex or hippocampus), striatal PV+ neurons are “fast-spiking” cells that can sustain high-frequency action potential firing with little afterhyperpolarization or spike frequency adaptation [21]. PV cells create a unique network that is interconnected by electrical synapses thanks to dendrodendritic gap junctions, enabling them to fire almost synchronously.
In an in situ hybridization study, cholinergic interneurons of the striatum were found to express GDNF mRNA [22]. Cholinergic interneurons play a key role in modulating the activity of striatum and in regulating dopaminergic and cholinergic signaling [23]. Interestingly, cholinergic interneurons are relatively resistant to several excitotoxic insults [24].
Braak and coworkers described the neuropathology of PD and its progression and processed it into a staging scheme [6]. The striatum could be affected, but in very late stages [25].
Several transgenic models of PD are currently in use. After disease-causing genes were identified using linkage analysis and association analysis with familial and sporadic cases of PD, SNCA (α-Synuclein) [26,27] and several other familial PD-linked genes were described, such as parkin, DJ-1, PINK1, and LRRK2 [28,29,30].
In this study, we investigated neuropathological changes occurring in the striatum of a transgenic rat model of PD that overexpresses the human physiological α-syn. This model develops behavioral and clinical changes that recapitulate the disease, such as novelty-seeking, avoidance, and smell, as well as, later on, motor deficits. The neuropathological changes observed in this model involve the integrity of dopaminergic system [31]. In this view, we aimed at studying the striatum and its relevance to the disease.
The evaluation of GDNF distribution in the striatum, as related to neuronal degeneration in the rat PD model, was our interest, given its role in the survival of adult nigrostriatal dopaminergic neurons. We studied striatum morphology and the specific subsets of neurons possibly involved in the GDNF role in PD. Thus, if our hypothesis was correct, a depletion in PV GABAergic interneurons would result in a downregulation of GDNF, which in turn would facilitate the degeneration of substantia nigra.
2. Results
2.1. Striatal Neuronal Cell Counts
Nissl staining performed on serial sections of rat brain tissue showed a significant neuronal depletion in the 12-month-old Snca+/+ rats (Figure 1A–D). The analysis carried out on the basis of immunofluorescence experiments using a specific marker for the medium spiny projection neurons, Calbindin, showed medium spiny neurons classically distributed in a patch and matrix compartment [32]. However, a significant numerical reduction in Calbindin-positive neurons was observed in adult rats, mainly in the dorso-lateral and ventral part of striatum (Figure 1L). In the wild type rats, the average number of Calbindin-positive neurons was 80+/−10, and this neuronal population was also observed in the 5-month-old Snca+/+ (Figure 1E,F). Over time, a statistically significant reduction in medium spiny neurons manner was observed in a genotype-dependent in the 12-month-old Snca+/+ rats. The statistical analysis, two-way ANOVA performed for the immunofluorescence intensity of the marker Calbindin, detected a statistically significant reduction in animals at 12 months of age (Figure 1P).
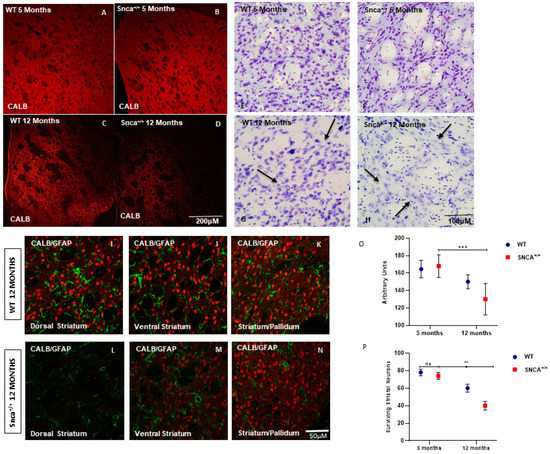
Figure 1.
Striatum pathology. Confocal laser scanning microscopy images of single-label immunofluorescence for the marker of the medium spiny neurons, Calbindin D-28K (red), in the striatum of 5-month-old WT (A), 5-month-old Snca+/+ (B), 12-month-old WT (C), and 12-month-old Snca+/+ (D). Striatum Nissl staining. Representative images of coronal striatal slices (E–H) showing the dramatic reduction in surviving neurons in 12-month-old Snca+/+ (arrows) compared to WT littermate (arrows). WT 12 months: (I) dorsal striatum, (J) ventral striatum, (K) striatum/pallidum; Snca+/+ 12 months: (L) dorsal striatum, (M) ventral striatum, (N) striatum/pallidum; (O,P) Quantification of the number of striatal neurons labeled with CALB. A two-way ANOVA indicated a significant effect of genotype [F(1.24) = 7.86; ** p = 0.0098]; time [F(1.24) = 36.9 *** p < 0.001] and genotype X time interaction (F(1.24) = 3.49; p = 0.05). “ns” means “not significant”.
2.2. Interneuron Subtype Distribution
Medium spiny projecting neurons (MSNs) represent 90–95% of the rodent striatal neurons, and the remaining cells are a mixed population of interneurons [33,34]. Among these, a small percentage (0.3% of all striatal neurons) comprises cholinergic interneurons, and the rest are GABAergic interneurons. GABAergic interneurons are further divided into several neurochemically distinct classes. Approximately 0.7% of all striatal neurons are interneurons that express the calcium binding protein PV [28]. Interestingly, the number of ChAT neurons remained almost unchanged in the WT rats at 5 and 12 months of age, as well as in Snca+/+ rats at the same time points (Figure 2E,F). Conversely, we observed a statistically significant loss the PV+ GABAergic interneurons over time only in the Snca+/+ rats (Figure 3D). Five-month-old WT and Snca+/+ animals showed many PV+ GABAergic interneurons in the dorsal-lateral part of the striatum, where their ontogenic maturation takes place (Figure 3A–C). This condition persisted in the 12-month-old WT rats, while it drastically changed in the 12-month-old rats, whereby most of the PV+ GABAergic interneurons died, mainly in the dorso-lateral part of the striatum. The few surviving PV+ interneurons showed a significantly reduced intensity of immunofluorescence, and a smaller soma with fewer arborizations (Figure 3E,F) (Figure S1).
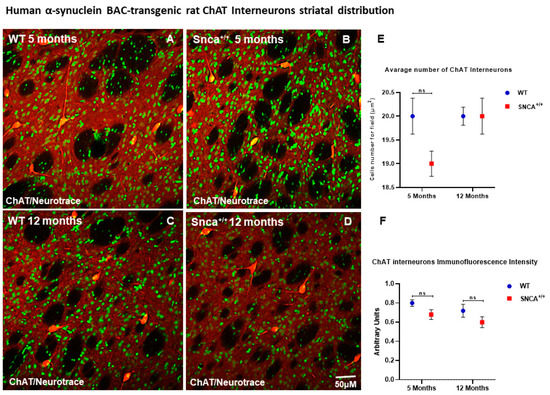
Figure 2.
ChAT Interneuron Distribution. Confocal microscopy-acquired images of single-label immunofluorescence for ChAT and counterstained with fluorescent Nissl staining. ChAT is shown in red fluorescence, Neurotrace is labeled in green. (A–D) Images show the immunoreaction intensity of ChAT in each experimental group. (E,F) Two-way ANOVA analysis performed on data obtained from 5- and 12-month-old WT and Snca+/+ showed no statistically significant (ns) change in ChAT-positive neurons and intensity.
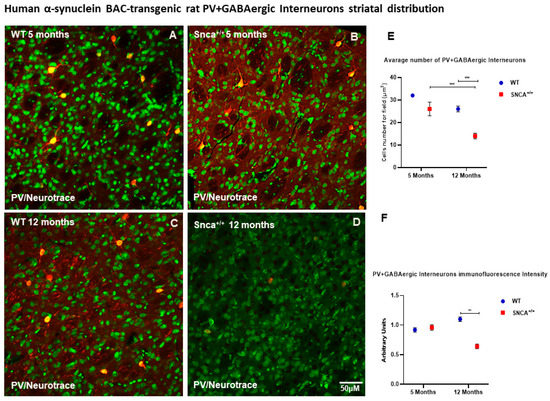
Figure 3.
PV+ GABAergic distribution. Representative confocal microscopy-acquired images of single-label immunofluorescence for PV counterstained with Neurotrace. (A–D) Images show the shape and immunoreaction intensity of Pv+ GABAergic interneurons in each experimental group. (E,F) Two-way ANOVA analysis performed on data obtained from 5- and 12-month-old WT and Snca+/+ showed a statistically significant reduction in number [genotype and time effect F(1.25) = 23.40 *** p < 0.001] and intensity [genotype effect F(1.207) = 10.80 ** p < 0.0012] of Parvalbumin-positive striatal neurons in 12-month-old Snca+/+ rats.
2.3. GDNF Expression in the Snca+/+ Rats’ Striatum
For the first time, the distribution of GDNF expression in the striatal neurons was evaluated in the SNCA genetic model of PD, with a focus on medium spiny projection and ChAT and PV+ GABAergic interneurons. Data obtained by analysis of immunofluorescence images revealed the gradual increase in GDNF between 5- and 12-month-old rats, pointing out the importance of this neurotrophic factor for the neuronal maturation. The immunohistochemistry reaction highlights a specific expression of GDNF in the PV+ GABAergic neurons (Figure 4A,B). Specifically, the immunofluorescence intensity analysis revealed the expression of GDNF in MSNs (Figure 4E–G) and ChAT interneurons, where the accumulation of GDNF takes on the appearance of puncta deposits (Figure 4I–K). Subsequently, we evaluated what happens to the PV+ GABAergic interneurons, whose ability to produce GDNF is known. For the first time, we observed a significant colocalization of GDNF in the PV+ GABAergic interneurons in the 5- and 12-month-old rats, which is only retained in the 12 adult WT (Figure 5G), while it is reduced in the adult Snca+/+, suggesting that the absence of this factor contributes to the specific death of these neurons (Figure 5L). This correlates with the presence of resistant PK aggregates and the increase in the phosphorylated form of human synuclein expression, the function of which could be to alter the ability of PV+ GABAergic interneurons to produce GDNF (Figure S1).
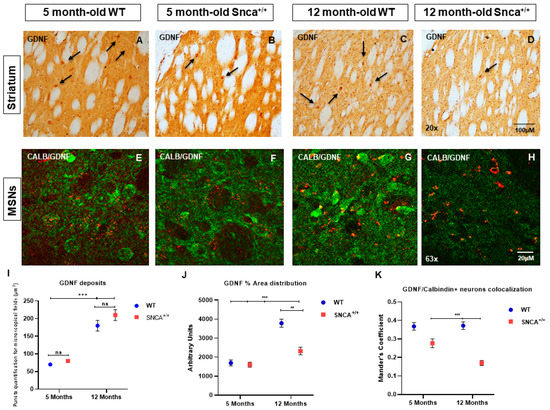
Figure 4.
(A–H) GDNF in striatal projection neurons of WT and Snca+/+ rats. Representative transmitted light microscope images showing Dab staining for GDNF counterstained with Hematoxylin in the striatum of 5- and 12-month-old Wt and Snca+/+ rats. (Arrows show the distribution of GDNF in the interneurons). Two-way ANOVA revealed a statistically significant effect of Time (F1.157 = 115.1; *** p < 0.001) in GDNF deposit formation in the experimental groups of 5- and 12-month-old WT and Snca+/+ rats, and a statistically significant genotype and time effect in GDNF area distribution [genotype F(1.157) = 17.84 ** p < 0.01; time F(1.157) = 57.99; *** p < 0.001]. “ns” means “not significant”. (I) GDNF deposits; (J) GDNF% area distribution; (K) GDNF/Calbindin + neurons colocalization.
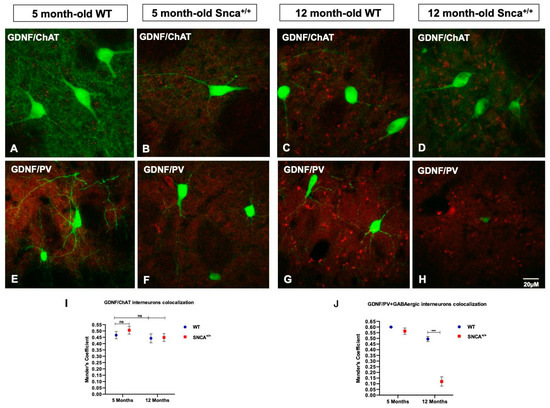
Figure 5.
GDNF in striatal interneurons. (A–D) Representative confocal images show the immunoreaction intensity of ChAT and GDNF in each experimental group. (E–H) Images show the immunoreaction intensity of PV+ GABAergic neurons and GDNF in each experimental group. (I,J) Quantitative analysis of coexpression levels by Mander’s coefficient in all experimental groups. Histogram represents the mean of ROIs calculated on the basis of Mander’s coefficients, revealing the statistically significant colocalization of GDNF in ChAT- and PV-positive neurons, at *** p < 0.001, only in the 5-month-old WT and Snca+/+ rats. “ns” means “not significant”.
2.4. Altered GDNF Protein Expression in the Substantia Nigra
In young rats, we observed an intense immunohistochemical staining of GDNF in the SN pars compacta and reticulata. However, the intensity of staining statistically decreases only in the 12-month-old Snca+/+ rats, in which a reduction in positive cells for GDNF was observed (Figure 6).
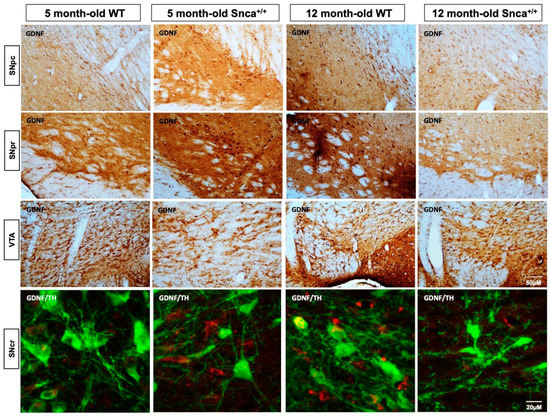
Figure 6.
GDNF in the Snca+/+ Substantia Nigra. Representative photomicrograph of DAB immunohistochemistry for the marker GDNF in rats Substantia Nigra. A qualitative intensity of colorimetric reaction is observed in the SNpr of the 12-month-old WT rats. These data are confirmed by confocal z-stack images of a double immunofluorescence for GDNF and TH in which colocalization analysis show a Mander’s coefficient greater than 0.7 in 5 and 12-month-old Wt that is significantly reduced in 12-month-old Snca+/+, in which it is less than 0.2.
3. Discussion
The striatum is a major component of the basal ganglia, and is involved in controlling motor activity and reward behavior [35,36]. Different cell types compose the striatum, where 90–95% are GABAergic medium spiny neurons (MSNs), and the remaining 5–10% is constituted by interneurons [37,38]. Striatal interneurons exert a powerful pre- and post-synaptic modulation of striatal functions [39], which implicates them in many movement and psychiatric disorders [40,41].
In this study, we investigated changes in the striatum of a transgenic rat model pf PD. We observed a marked neurodegeneration of the dorsolateral part of the striatum, with a relative preservation of the striosome (patch-matrix) distribution [42] in the surviving areas.
DA projections from the substantia nigra innervate both patch and matrix [43,44], even though matrix is enriched with DA and patch is not. Additionally, the two subclasses of medium spiny neurons, the D1 Receptor and D2 Receptor expressing projection neurons, can reside in either the striosome or matrix compartment; therefore, both compartments contribute to the direct and indirect output pathways. PD is characterized by the progressive degeneration of nigrostriatal pathway, consisting of dopaminergic neuronal bodies of the substantia nigra and their projections to the striatum. Thus, the observation of unchanged patch–matrix compartments in this model could be explained by the notion that the degeneration primarily derives from substantia nigra, which evenly innervates both patch and matrix.
As mentioned above, we observed a marked neuronal degeneration in the dorsolateral part of the anterior striatum. The reason for this dramatic loss of neurons in the striatum in a model of PD is yet to be elucidated. However, striatal neuronal degeneration was previously observed in the MPTP/3-NP model of multiple system atrophy [45], and in PD patients [46,47]. Thus, we can speculate that striatal pathology occurs in the later stages of the disease, at least in this model of PD. From a broader perspective, if striatal degeneration precedes the death of SN, in could be inferred that an altered function of DA neurons would provoke the degeneration of their striatal target.
Several cerebral pathways and circuits that could be involved in PD degeneration have emerged over the years. An imbalance in the activity of cerebellar pathways has been reported in PD [48], such as cerebello–thalamo–cortical circuitry [49]. In a voxel-based functional connectivity analysis, Shen and co-workers (2020) showed an alteration in the connectivity of basal ganglia-cortical circuit in patients with PD that was related to postural instability and gait difficulty. They also suggested, in PD patients with predominant tremor and lesser postural instability and gait impairment, a relation with increased functional connectivity between putamen and cerebellum [50].
Neuronal degeneration involves multiple regions of the brain beyond substantia nigra and VTA, such as locus caeruleus, dorsal raphe nucleus [51], and the dorsal motor nucleus of the vagus. Thus, common features explaining neuronal vulnerability to PD have been investigated. Axonal arborization size, iron content, and autonomous pacemaking activity are some of the biological characteristics that have been implied with cell vulnerability [52,53,54]
In our study, neuronal loss involved mainly the calbindin positive medium spiny projection neurons, along with a subset of interneurons, namely, the parvabuminergic interneurons.
In the striatum, PV interneurons primarily downregulate the activity of medium spiny projection neurons by means of a monosynaptic inhibition, with a feed-forward mechanism [55,56]. In SN, excitoxicity and oxidative stress lead to raised intracellular calcium levels. In this situation, an increase in PV expression was protective [57].
Here, we observed a time-sensitive degeneration of striatal parvalbuminergic neurons that was present in 5-month-old SNCA transgenic rats and dramatically worse at 12 months. Other interneurons were relatively preserved, while medium spiny projection neurons decreased progressively. Thus, it appeared that PV neurons were the most vulnerable to the degeneration arising from nigral disfunction due to the SNCA mutation.
PV neurons are evenly distributed throughout the striatum, where they provide a homogenous trophic support to the richly ramified axons of nigrostriatal DA neurons. Part of this important trophic function is exerted through the production of GDNF that is retrogradely transported from the DA nerve terminals to the somata located in the SN [58].
In the striatum, GDNF protein expression was decreased markedly in both projection neurons and in degenerating PV interneurons. GDNF protein appeared to be in the neuropil of SNCA+/+ animals, possibly deriving from other sources [59]. Indeed, in several disease models, neuroinflammation can upregulate GDNF expression in activated astrocytes [60,61].
PV interneurons are indeed the main source of striatal GDNF [62,63]. Cholinergic striatal interneurons are also a source of GDNF, and we observed that this population was relatively spared by the degeneration of neurons in this disease model.
GDNF displays beneficial effects with respect to the survival of mesencephalic dopaminergic neurons and on noradrenergic neurons of the locus coeruleus [64]. Neuroprotective effects of intrastriatally administered GDNF were observed in animal models of PD [65,66,67]. Moreover, the effect of intrastriatal GDNF administration has been tested in several studies performed in PD patients. However, the clinical benefits of GDNF-based therapies have not been demonstrated [68]. Alternative, indirect GDNF-based therapies have been tested over the years, such as lentivirus (LV) vector transgenes fused with a destabilizing domain [69].
We observed a dramatic decrease in GDNF immunostaining in the dopaminergic neurons of substantia nigra (Figure 6). Thus, our hypothesis that PV interneurons are involved in GDNF signaling in this model of disease was confirmed. Transport of GDNF occurs through a retrograde transport [70].
Our data suggest that the damage at the level of PV+ GABAergic interneurons in the striatum not only determines a reduction in the production of GDNF, but also severely compromises their intrinsic ability to retrogradely transport GDNF to the SN, thus promoting the neurodegenerative process.
Therefore, decreased GDNF availability translates into failure by DA nerve endings to take GDNF from the striatum and transport it retrogradely to the SN, thus contributing to the degeneration of SN. In support of this hypothesis, we previously described, in another α-syn rat model of PD, that the main alterations occurred in the DAergic and glutamatergic terminals of the dorsolateral striatum and that these preceded SN dopamine neuron degeneration [71]. Alternatively, primarily degenerating DA neurons would cause their nerve endings to decrease their ability to transport GDNF to SN, thus worsening the neuropathology of the disease.
With both hypotheses being open and needing further investigation, our work has highlighted the importance of GDNF and contributes to the evidence of possible therapeutic use of this trophic factor in PD. Considering the difficulties in the direct administration of trophic factors, [72], possible strategies for PD might include non-pharmacological therapies such as direct current stimulation [73], not only because of its effects on degradation of α-synuclein, but also because of its possible role in determining an increase in trophic factors [74].
4. Materials and Methods
4.1. Genetic Animal Model
All animal experiments, which satisfied ARRIVE guidelines, were performed in accordance with European Communities Council Directive (2010/63 EU) as adopted by the Santa Lucia Foundation Animal Care and Use and approved by the Italian Ministry of Health (Authorization No. 617-2019PR). Homozygous bacterial artificial chromosome (BAC) transgenic rats (Sprague-Dawley background) overexpressing the full-length human Snca locus under the control of the endogenous human regulatory elements (Snca+/+) and wild type (WT) littermates of 5 and 12 months of age were used.
4.2. Histological and Immunohistochemical Studies
- Tissue processing
The study groups included: 5-month-old wild type rats (WT), 5-month-old Snca+/+ rats (Snca+/+) and 12-month-old WT and Snca+/+ rats. Seven rats per group were used, and they were handled under the same conditions by one investigator at the same day and time. All rats were transcardially perfused under deep anesthesia with saline solution containing 0.01 mL heparin, followed by 4% paraformaldehyde (PFA) in phosphate buffer (PB; 0.1 M, pH 7.4). Brains were removed and post-fixed in 4% paraformaldehyde at 4 °C overnight and subsequently transferred into 30% sucrose solution at 4 °C until sinking. Sectioning was performed on a sliding cryostat (Leica, Wetzlar, Germany) at a thickness of 40 μm. Blinded observers collected primary data.
- Histological and immunohistochemical studies
For the immunohistochemical studies, primary omission controls, normal mouse, and rabbit serum controls and preimmune serum controls were used to confirm the specificity of our immunohistochemical labeling.
Striatal neuronal cell count. Single-label immunofluorescence was performed using an antibody against Calbindin D-28K (marker of the Medium Spiny Neurons) (CALB, Immunological Sciences, Rome, Italy) to evaluate the number of surviving projection neurons in the striatum. Cell counts were carried out in each of 24 1.0-mm-square confocal microscope fields, rostrocaudally spaced on both hemispheres of 7 mice from each group. For cell counts, we used the Java image processing and analysis program Image J, developed by Wayne Rasband, available in the public domain (http://imagej.nih.gov/ij/docs/index.html (accessed on 1 January 2020). We used a manual approach, measuring the number of objects by means of the point selection tool.
- Striatal interneuron subtype characterization
Immunohistological staining for striatal interneurons markers was performed. Brain sections were incubated with goat anti-choline acetyl transferase (ChAT; Novus Biological, Bio-Techne, Milan, Italy) and mouse anti-parvalbumin (PARV, Chemicon International, Inc., Temecula, CA, USA). All primary antibodies were used at a dilution of 1:200, in 0.1 M PB containing 0.3% Triton X-100 for 72 h at 4 °C. Sections were rinsed three times for 5 min at room temperature and subsequently incubated with secondary antibodies Alexa Fluor 488 and 555 for 2 h at room temperature at a dilution of 1:200 in a 0.1 M PB solution containing 0.3% Triton X-100. Confocal laser scanner microscopy (Zeiss, Oberkochen, Germany LSM800) was used to acquire images. Immunofluorescence analyses were performed using the Java image processing and plugin analysis program included in Fiji ImageJ.
- Immunohistochemistry for GDNF
Peroxidase-antiperoxidase diaminobenzidine tetrahydrochloride single-label immunohistochemistry for GDNF was performed to identification and quantification of neurons involved in its production in the striatum. Serial sections were incubated with rabbit anti-GDNF (Novus Biological, Bio-techne, Milan, Italy) at a dilution of 1:100 in 0.1 M PB solution containing 0.3% Triton X-100 for 72 h at 4 °C. Subsequently, sections were incubated with rabbit peroxidase–antiperoxidase complex at a dilution of 1:100 in 0.1 M PB solution with 0.3% Triton X-100 at room temperature for 1 h. After peroxidase–antiperoxidase incubation, sections were incubated in Tris-Hcl buffer containing 10 mg diaminobenzidine tetrahydrochloride for 2 min, adding 15 µL of 3% hydrogen peroxidase. The peroxidase–antiperoxidase diaminobenzidine tetrahydrochloride-labeled sections were then washed in distilled water, placed in 0.1 M PB, mounted on gelatin-coated slides, dried, dehydrated and coverslipped. Collected images of GDNF positive neurons were obtained by Olympus U-RFLT 200 software (Olympus, Tokyo, Japan).
- Analysis of colocalization of GDNF in striatal projection neurons, interneurons, and dopaminergic neurons
Double-label immunofluorescence was performed to evaluate the colocalization of GDNF in Calbindin-labeled striatal neurons, ChAT+, PV+ interneurons and TH+ dopaminergic neurons. Antigen retrieval was performed in Citrate Buffer (pH 6–7) for 20 min at 80 °C. Subsequently, sections were incubated with a cocktail of anti-GDNF, ChAT, PARV and TH (monoclonal mouse antibody, Millipore, Italy) antibodies for 72 h at +4 °C. A streptavidin–biotin amplification method for GDNF immunofluorescence staining was used. A confocal laser scanning microscope was used to acquire all the images. Three separate fields (dorsolateral, central, and medial each 1 mm in diameter) in each of three rostrocaudally spaced sections of six mice per group were examined. GDNF, Calbindin, ChAT, PV and TH immunofluorescence intensity was measured and quantified by using the Java image processing and analysis program available in, Fiji ImageJ. All confocal z-stacks images were acquired under no saturation conditions, with a ×63 objective raising a ×1 zoom with an XY resolution of 1024 × 1024 and 0.60 μm step-size along the vertical z-axis. The same set configuration was performed for all samples. Colocalization of GDNF in striatal projection neurons, interneurons, and dopaminergic neurons was calculated using the JACoP tool measures colocalization by performing a correlation coefficient-based analysis of the pixel intensity of different color channels. JACoP allows the use and comparison of some common intensity coefficient-based methods, including Pearson’s coefficient, Manders’ coefficient, Costes’s approach, and object-based analysis. The mean Mander’s coefficients were obtained by performing Costes Autothresholds on the samples’ regions of interest (ROIs) and running the analysis on 100 randomized images, and varied from 0 to 1, corresponding to non-overlapping images and 100% or quite colocalization between the two images, respectively.
4.3. Statistical Analysis
All the collected images were quantified by using the Java image processing and analysis program Fiji ImageJ. Cells of interest were selected using the freehand tool. From the Analyze menu, set measurements Mean “Gray Value”, “Area” and “Min and Max Gray Value” were selected. The region characterized by absence of fluorescence was considered in the background and it was subtracted. Finally, the mean values with SEM were obtained for all measures. ANOVA analysis available in the software GraphPad Prism version 9.0 was performed. p values < 0.05 were considered statistically significant.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms231710126/s1.
Author Contributions
E.P.: study design, experiments, project administration; V.D.: investigation; M.M.C.: investigation; E.G.: colony maintenance, investigation; S.B.: investigation; G.M.: investigation; N.B.: colony maintenance, investigation; O.R.: Resources, review and critique; G.S.: review and critique; N.B.M.: review and critique; F.R.F.: study design, writing—original draft preparation, supervision, final review and critique. All authors have read and agreed to the published version of the manuscript.
Funding
This project was supported by the Italian Ministry of Health (Ricerca Finalizzata #RF-2018-12365509 to N.B.M.).
Institutional Review Board Statement
All animal experiments, which satisfied ARRIVE guidelines, were performed in accordance with European Communities Council Directive (2010/63 EU) as adopted by the Santa Lucia Foundation Animal Care and Use and approved by the Italian Ministry of Health (Authorization No. 617-2019PR).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Monchi, O.; Petrides, M.; Mejia-Constain, B.; Strafella, A.P. Cortical activity in Parkinson’s disease during executive processing depends on striatal involvement. Brain 2007, 130, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A. Neuropathology of sporadic Parkinson’s disease evaluation and changes of concepts. Mov. Disord. 2012, 27, 8–30. [Google Scholar] [CrossRef] [PubMed]
- Dauer, W.; Przedborski, S. Parkinson’s diease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef]
- Owen, A.M. Cognitive dysfunction in Parkinson’s disease: The role of frontostriatal circuitry. Neuroscientist 2004, 10, 525–537. [Google Scholar] [CrossRef]
- Braak, H.; Ghebremedhin, E.; Rub, U.; Bratzke, H.; Del Tredici, K. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res. 2004, 318, 121–134. [Google Scholar] [CrossRef]
- Tsuboi, Y.; Dickson, D.W. Dementia with Lewy bodies and Parkinson’s disease with dementia: Are they different? Parkinsonism Relat. Disord. 2005, 11 (Suppl. 1), S47–S51. [Google Scholar] [CrossRef]
- Steeves, T.D.; Miyasaki, J.; Zurowski, M.; Lang, A.E.; Pellecchia, G.; Van Eimeren, T.; Rusjan, P.; Houle, S.; Strafella, A.P. Increased striatal dopamine release in Parkinsonian patients with pathological gambling: A [11C] raclopride PET study. Brain 2009, 132, 1376–1385. [Google Scholar] [CrossRef]
- Taylor, A.E.; Saint-Cyr, J.A.; Lang, A.E. Frontal lobe dysfunction in Parkinson’s disease. The cortical focus of neostriatal outflow. Brain 1986, 109, 845–883. [Google Scholar] [CrossRef]
- Zgaljardic, D.J.; Foldi, N.S.; Borod, J.C. Cognitive and behavioral dysfunction in Parkinson’s disease: Neurochemical and clinicopathological contributions. J. Neural Transm. 2004, 111, 1287–1301. [Google Scholar] [CrossRef]
- Alexander, G.E.; DeLong, M.R.; Strick, P.L. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 1986, 9, 357–381. [Google Scholar] [CrossRef] [PubMed]
- Connolly, B.; Fox, S.H. Treatment of cognitive, psychiatric, and affective disorders associated with Parkinson’s disease. Neurother. J. Am. Soc. Exp. Neurother. 2014, 11, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Duncan, G.W.; Khoo, T.K.; Yarnall, A.J.; O’Brien, J.T.; Coleman, S.Y.; Brooks, D.J.; Barker, R.A.; Burn, D.J. Health-related quality of life in early Parkinson’s disease: The impact of nonmotor symptoms. Mov. Disord. Off. J. Mov. Disord. Soc. 2014, 29, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Mufti, S.; LaFaver, K. Mood Disorders in Parkinsons Disease. Psychiatr. Ann. 2020, 50, 95–99. [Google Scholar] [CrossRef]
- Lin, L.F.; Doherty, D.H.; Lile, J.D.; Bektesh, S.; Collins, F. GDNF: A glial cell line-derived neurotrophic factor for midbrain dopaminergic neurons. Science 1993, 260, 1130–1132. [Google Scholar] [CrossRef]
- Ibáñez, C.F. Catecholaminergic neuron survival: Getting hooked on GDNF. Nat. Neurosci. 2008, 11, 735–736. [Google Scholar] [CrossRef]
- Pascual, A.; Hidalgo-Figueroa, M.; Piruat, J.I.; Pintado, C.O.; Gómez-Díaz, R.; López-Barneo, J. Absolute requirement of GDNF for adult cat- echolaminergic neuron survival. Nat. Neurosci. 2008, 11, 755–761. [Google Scholar] [CrossRef]
- Beck, K.D.; Valverde, J.; Alexi, T.; Poulsen, K.; Moffat, B.; Vandlen, R.A.; Rosenthal, A.; Hefti, F. Mesencephalic dopaminergic neurons protected by GDNF from axotomy-induced degeneration in the adult brain. Nature 1995, 373, 339–341. [Google Scholar] [CrossRef]
- Hidalgo-Figueroa, M.; Bonilla, S.; Gutiérrez, F.; Pascual, A.; López-Barneo, J. GDNF Is Predominantly Expressed in the PV+ Neostriatal Interneuronal Ensemble in Normal Mouse and after Injury of the Nigrostriatal Pathway. J. Neurosci. 2012, 32, 864–872. [Google Scholar] [CrossRef]
- Björklund, A.; Dunnett, S.B. Dopamine neuron systems in the brain: An update. Trends Neurosci. 2007, 30, 194–202. [Google Scholar] [CrossRef]
- Tepper, J.M.; Koós, T.; Ibanez-Sandoval, O.; Tecuapetla, F.; Faust, T.W.; Assous, M. Heterogeneity and Diversity of Striatal GABAergic Interneurons: Update 2018. Front. Neuroanat. 2018, 12, 91. [Google Scholar] [CrossRef] [PubMed]
- Bizon, J.L.; Lauterborn, J.C.; Gall, C.M. Subpopulations of striatal interneurons can be distinguished on the basis of neurotrophic factor expression. J. Comp. Neurol. 1999, 408, 283–298. [Google Scholar] [CrossRef]
- Tanimura, A.; Pancani, T.; Lim, S.A.O.; Tubert, C.; Melendez, A.E.; Shen, W.; Surmeier, D.J. Striatal cholinergic interneurons and Parkinson’s disease. Eur. J. Neurosci. 2018, 47, 1148–1158. [Google Scholar] [CrossRef]
- Fusco, F.R.; Zuccato, C.; Tartari, M.; Martorana, A.; de March, Z.; Giampà, C.; Cattaneo, E.; Bernardi, G. Co-localization of brain-derived neurotrophic factor (BDNF) and wild-type huntingtin in normal and quinolinic acid-lesioned rat brain. Eur. J. Neurosci. 2003, 18, 1093–1102. [Google Scholar] [CrossRef]
- Graybiel, A.M.; Hirsch, E.C.; Agid, Y. The nigrostriatal system in Parkinson’s disease. Adv. Neurol. 1990, 53, 17–29. [Google Scholar]
- Billingsley, K.J.; Bandres-Ciga, S.; Saez-Atienzar, S.; Singleton, A.B. Genetic risk factors in Parkinson’s disease. Cell Tissue Res. 2018, 373, 9–20. [Google Scholar] [CrossRef]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [Google Scholar] [CrossRef]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef]
- Zimprich, A.; Biskup, S.; Leitner, P.; Lichtner, P.; Farrer, M.; Lincoln, S.; Kachergus, J.; Hulihan, M.; Uitti, R.J.; Calne, D.B.; et al. Mutations in LRRK2 cause autosomal-dominant parkinsonism with pleomorphic pathology. Neuron 2004, 44, 601–607. [Google Scholar] [CrossRef]
- Nuber, S.; Harmuth, F.; Kohl, Z.; Adame, A.; Trejo, M.; Schönig, K.; Zimmermann, F.; Bauer, C.; Casadei, N.; Giel, C.; et al. A progressive dopaminergic phenotype associated with neurotoxic conversion of α-synuclein in BAC-transgenic rats. Brain J. Neurol. 2013, 136, 412–432. [Google Scholar] [CrossRef] [PubMed]
- Brimblecombe, K.R.; Cragg, S.J. The Striosome and Matrix Compartments of the Striatum: A Path through the Labyrinth from Neurochemistry toward Function. ACS Chem Neurosci. 2017, 8, 235–242. [Google Scholar] [CrossRef]
- Prensa, L.; Giménez-Amaya, J.M.; Parent, A. Chemical heterogeneity of the striosomal compartment in the human striatum. J. Comp. Neurol. 1999, 413, 603–618. [Google Scholar] [CrossRef]
- Johnston, J.G.; Gerfen, C.R.; Haber, S.N.; van der Kooy, D. Mechanisms of striatal pattern formation: Conservation of mammalian compartmentalization. Dev. Brain Res. 1990, 57, 93–102. [Google Scholar] [CrossRef]
- Ferrante, R.J.; Kowall, N.W. Tyrosine hydroxylase-like immunoreactivity is distributed in the matrix compartment of normal human and Huntington’s disease striatum. Brain Res. 1987, 416, 141–146. [Google Scholar] [CrossRef]
- Prensa, L.; Cossette, M.; Parent, A. Dopaminergic innervation of human basal ganglia. J. Chem. Neuroanat. 2000, 20, 207–213. [Google Scholar] [CrossRef]
- Huot, P.; Lévesque, M.; Parent, A. The fate of striatal dopaminergic neurons in Parkinson’s disease and Huntington’s chorea. Brain 2007, 130, 222–232. [Google Scholar] [CrossRef]
- Graybiel, A.M. The basal ganglia and chunking of action repertoires. Neurobiol. Learn. Mem. 1998, 70, 119–136. [Google Scholar] [CrossRef]
- Schultz, W. Getting formal with dopamine and reward. Neuron 2002, 36, 241–263. [Google Scholar] [CrossRef]
- Gerfen, C.R.; Bolam, J.P. Chapter 1-The Neuroanatomical Organization of the Basal Ganglia. Handb. Behav. Neurosci. 2010, 20, 3–28. [Google Scholar]
- Petryszyn, S.; Parent, A.; Parent, M. The calretinin interneurons of the striatum: Comparisons between rodents and primates under normal and pathological conditions. J. Neural. Transm. 2018, 128, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.A.; Ding, J.B.; Surmeier, D.J. Muscarinic Modulation of Striatal Function and Circuitry. In Muscarinic Receptors; Springer: Berlin/Heidelberg, Germany, 2012; pp. 223–241. [Google Scholar]
- Pisani, A.; Bernardi, G.; Ding, J.; Surmeier, D.J. Re-emergence of striatal cholinergic interneurons in movement disorders. Trends Neurosci. 2007, 30, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Won, L.; Britt, J.P.; Lim, S.A.O.; McGehee, D.S.; Kang, U.J. Enhanced striatal cholinergic neuronal activity mediates l-DOPA–induced dyskinesia in parkinsonian mice. Proc. Natl. Acad. Sci. USA 2011, 108, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Gerfen, C.R.; Herkenham, M.; Thibault, J. The neostriatal mosaic: II. Patch- and matrix-directed mesostriatal dopaminergic and non-dopaminergic systems. J. Neurosci. 1987, 7, 3915–3934. [Google Scholar] [CrossRef]
- Matsuda, W.; Furuta, T.; Nakamura, K.C.; Hioki, H.; Fujiyama, F.; Arai, R.; Kaneko, T. Single nigrostriatal dopaminergic neurons form widely spread and highly dense axonal arborizations in the neostriatum. J. Neurosci. 2009, 29, 444–453. [Google Scholar] [CrossRef]
- Stefanova, N.; Puschban, Z.; Fernagut, P.O.; Brouillet, E.; Tison, F.; Reindl, M.; Jellinger, K.A.; Poewe, W.; Wenning, G.K. Neuropathological and behavioral changes induced by various treatment paradigms with MPTP and 3-nitropropionic acid in mice: Towards a model of striatonigral degeneration (multiple system atrophy). Acta. Neuropathol. 2003, 106, 157–166. [Google Scholar] [CrossRef]
- Simioni, A.C.; Dagher, A.; Fellows, L.K. Compensatory striatal-cerebel- lar connectivity in mild-moderate Parkinson’s disease. Neuroimage Clin. 2016, 10, 54–62. [Google Scholar] [CrossRef]
- Horne, M.K.; Butler, E.G. The role of the cerebello-thalamo-cortical pathway in skilled movement. Prog. Neurogibol. 1995, 46, 199–213. [Google Scholar] [CrossRef]
- Shen, B.; Pan, Y.; Jiang, X.; Wu, Z.; Zhu, J.; Dong, J.; Zhang, W.; Xu, P.; Dai, Y.; Gao, Y.; et al. Altered putamen and cerebellum connectivity among different subtypes of Parkinson’s disease. CNS Neurosci. Ther. 2020, 26, 207–214. [Google Scholar] [CrossRef]
- Cheshire, P.; Ayton, S.; Bertram, K.L.; Ling, H.; Li, A.; McLean, C.; Halliday, G.M.; O’Sullivan, S.S.; Revesz, T.; Finkelstein, D.I.; et al. Serotonergic markers in Parkinson’s disease and levodopa-induced dyskinesias. Mov. Disord. 2015, 30, 796–804. [Google Scholar] [CrossRef]
- Sian-Hulsmann, J.; Mandel, S.; Youdim, M.B.; Riederer, P. The relevance of iron in the pathogenesis of Parkinson’s disease. J. Neurochem. 2011, 118, 939–957. [Google Scholar] [CrossRef] [PubMed]
- Michel, P.P.; Alvarez-Fischer, D.; Guerreiro, S.; Hild, A.; Hartmann, A.; Hirsch, E.C. Role of activity-dependent mechanisms in the control of dopaminergic neuron survival. J. Neurochem. 2007, 101, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Bugiani, O.; Perdelli, F.; Salvarani, S.; Leonardi, A.; Mancardi, G.L. Loss of striatal neurons in Parkinson’s disease: A cytometric study. Eur. Neurol. 1980, 19, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Burguière, E.; Monteiro, P.; Feng, G.; Graybiel, A.M. Optogenetic stimulation of lateral orbitofronto-striatal pathway suppresses compulsive behaviors. Science 2013, 340, 1243–1246. [Google Scholar] [CrossRef]
- Qi, J.; Zhang, S.; Wang, H.L.; Barker, D.J.; Miranda-Barrientos, J.; Morales, M. VTA glutamatergic inputs to nucleus accumbens drive aversion by acting on GABAergic interneurons. Nat. Neurosci. 2016, 19, 725–733. [Google Scholar] [CrossRef]
- Soós, J.; Engelhardt, J.I.; Siklós, L.; Havas, L.; Majtényi, K. The expression of PARP, NF-κB and parvalbumin is increased in Parkinson disease. Neuroreport 2004, 15, 1715–1718. [Google Scholar] [CrossRef]
- d’anglemont de Tassigny, X.; Pascual, A.; López-Barneo, J. GDNF-based therapies, GDNF-producing interneurons, and trophic support of the dopaminergic nigrostriatal pathway. Implications for Parkinson’s disease. Front. Neuroanat. 2015, 9, 10. [Google Scholar]
- Duarte Azevedo, M.; Sander, S.; Tenenbaum, L. GDNF, A Neuron-Derived Factor Upregulated in Glial Cells during Disease. J. Clin. Med. 2020, 9, 456. [Google Scholar] [CrossRef]
- Merienne, N.; Meunier, C.; Schneider, A.; Seguin, J.; Nair, S.S.; Rocher, A.B.; Le Gras, S.; Keime, C.; Faull, R.; Pellerin, L.; et al. Cell-type-specific gene expression profiling in adult mouse brain reveals normal and disease-state signatures. Cell Rep. 2019, 26, 2477–2493. [Google Scholar] [CrossRef]
- Marco, S.; Canudas, A.M.; Canals, J.M.; Gavalda, N.; Perez-Navarro, E.; Alberch, J. Excitatory amino acids differentially regulate the expression of GDNF, neurturin, and their receptors in the adult rat striatum. Exp. Neurol. 2002, 174, 243–252. [Google Scholar] [CrossRef]
- Arenas, E.; Trupp, M.; Akerud, P.; Ibáñez, C.F. GDNF prevents degeneration and promotes the phenotype of brain noradrenergic neurons in vivo. Neuron 1995, 15, 1465–1473. [Google Scholar] [CrossRef]
- Sauer, H.; Rosenblad, C.; Bjorklund, A. Glial cell line-derived neurotrophic factor but not transforming growth factor beta 3 prevents delayed degeneration of nigral dopaminergic neurons following striatal 6-hydroxydopamine lesion. Proc. Natl. Acad. Sci. USA 1995, 92, 8935–8939. [Google Scholar] [CrossRef] [PubMed]
- Gash, D.M.; Zhang, Z.; Ovadia, A.; Cass, W.A.; Yi, A.; Simmerman, L.; Russell, D.; Martin, D.; Lapchak, P.A.; Collins, F.; et al. Functional recovery in parkinsonian monkeys treated with GDNF. Nature 1996, 380, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Whone, A.; Luz, M.; Boca, M.; Woolley, M.; Mooney, L.; Dharia, S.; Broadfoot, J.; Cronin, D.; Schroers, C.; Barua, N.U.; et al. Randomized trial of intermittent intraputamenal glial cell line-derived neurotrophic factor in Parkinson’s disease. Brain 2019, 142, 512–525. [Google Scholar] [CrossRef]
- Tomac, A.; Lindqvist, E.; Lin, L.F.; Ogren, S.O.; Young, D.; Hoffer, B.J.; Olson, L. Protection and repair of the nigrostriatal dopaminergic system by GDNF in vivo. Nature 1995, 373, 335–339. [Google Scholar] [CrossRef]
- Oppenheim, R.W.; Houenou, L.J.; Johnson, J.E.; Lin, L.F.; Li, L.; Lo, A.C.; Newsome, A.L.; Prevette, D.M.; Wang, S. Developing motor neurons rescued from programmed and axotomy-induced cell death by GDNF. Nature 1995, 373, 344–346. [Google Scholar] [CrossRef]
- Guatteo, E.; Rizzo, F.R.; Federici, M.; Cordella, A.; Ledonne, A.; Latini, L.; Nobili, A.; Viscomi, M.T.; Biamonte, F.; Landrock, K.K.; et al. Functional alterations of the dopaminergic and glutamatergic systems in spontaneous α-synuclein overexpressing rats. Exp. Neurol. 2017, 287, 21–33. [Google Scholar] [CrossRef]
- Yan, Q.; Matheson, C.; Lopez, O.T. In vivo neurotrophic effects of GDNF on neonatal and adult facial motor neurons. Nature 1995, 373, 341–344. [Google Scholar] [CrossRef]
- Tai, M.H.; Cheng, H.; Wu, J.P.; Liu, Y.L.; Lin, P.R.; Kuo, J.S.; Tseng, C.J.; Tzeng, S.F. Gene transfer of glial cell line-derived neurotrophic factor promotes functional recovery following spinal cord contusion. Exp. Neurol. 2003, 183, 508–515. [Google Scholar] [CrossRef]
- Sala, G.; Bocci, T.; Borzì, V.; Parazzini, M.; Priori, A.; Ferrarese, C. Direct current stimulation enhances neuronal alpha-synuclein degradation in vitro. Sci. Rep. 2021, 11, 2197. [Google Scholar] [CrossRef]
- Winkler, C.; Reis, J.; Hoffmann, N.; Gellner, A.K.; Münkel, C.; Curado, M.R.; Furlanetti, L.; Garcia, J.; Döbrössy, M.D.; Fritsch, B. Anodal Transcranial Direct Current Stimulation Enhances Survival and Integration of Dopaminergic Cell Transplants in a Rat Parkinson Model. eNeuro 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.R.; Machado-Vieira, R.; Zarate, C.A., Jr.; Vieira, E.L.; Valiengo, L.; Benseñor, I.M.; Lotufo, P.A.; Gattaz, W.F.; Teixeira, A.L. Assessment of non-BDNF neurotrophins and GDNF levels after depression treatment with sertraline and transcranial direct current stimulation in a factorial, randomized, sham-controlled trial (SELECT-TDCS): An exploratory analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 56, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Surguchov, A. Biomarkers in Parkinson’s Disease. In Neurodegenerative Diseases Biomarkers; Peplow, P.V., Martinez, B., Gennarelli, T.A., Eds.; Neuromethods; Humana: New York, NY, USA, 2022; Volume 173, pp. 155–180. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).