
Efficacy Comparison of LPA2 Antagonist H2L5186303 and Agonist GRI977143 on Ovalbumin-Induced Allergic Asthma in BALB/c Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
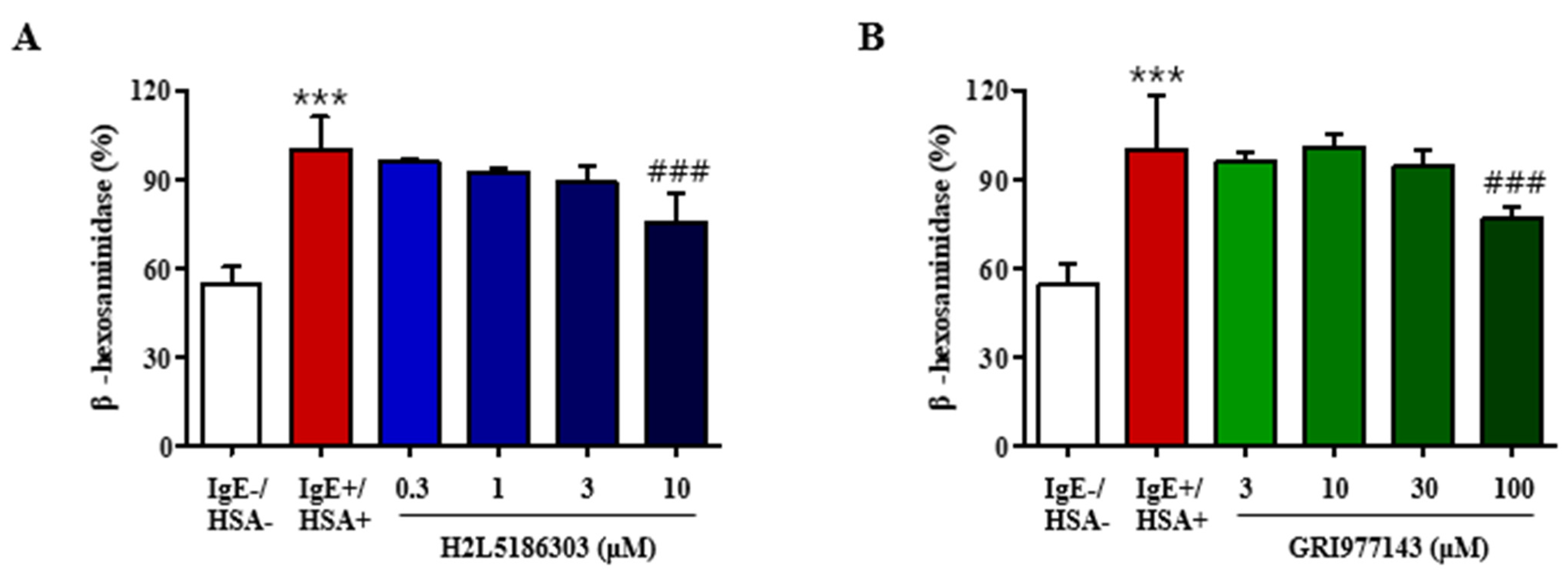
2.1. GRI977143 and H2L5186303 Suppressed Degranulation of Mast Cells
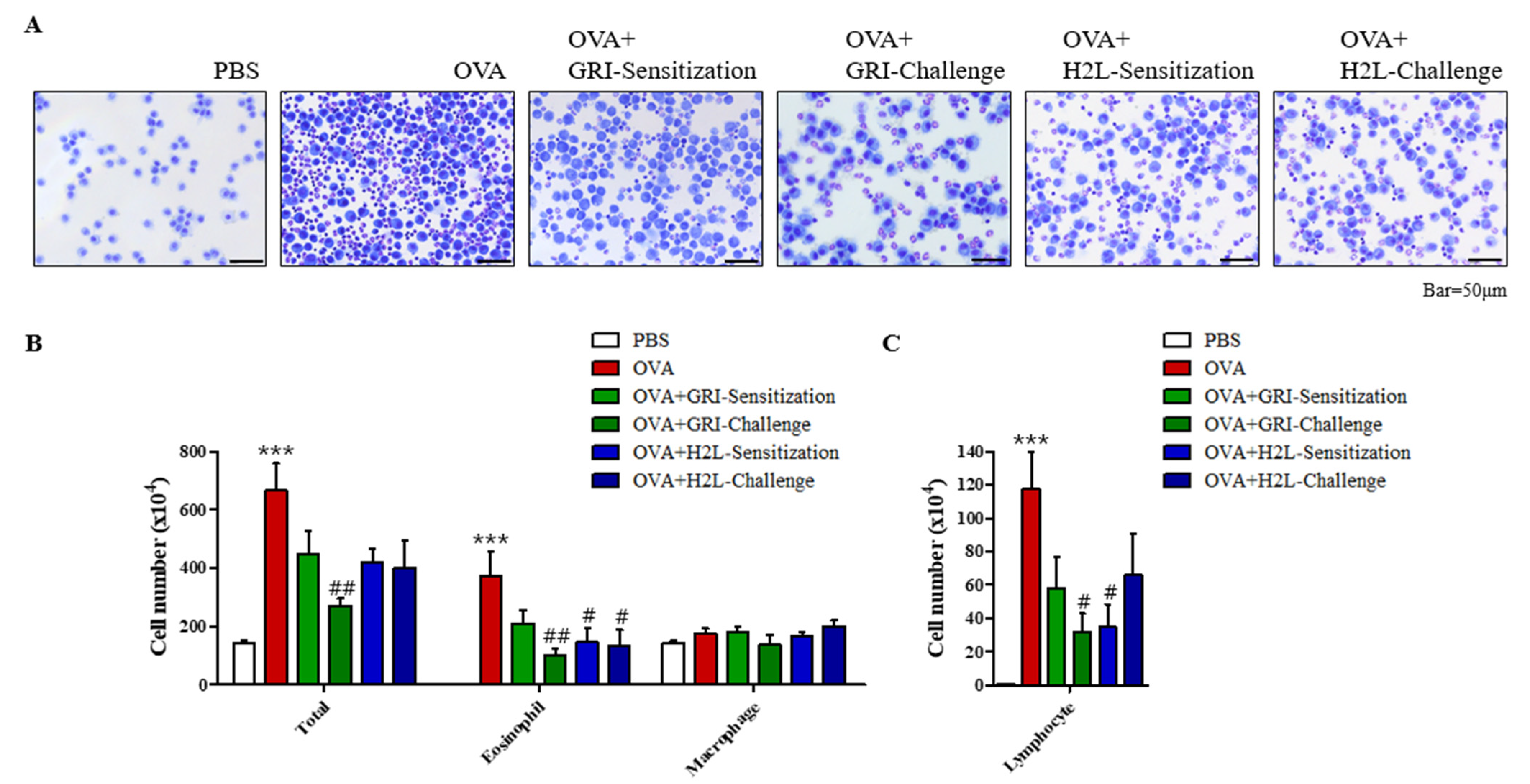
2.2. GRI977143 and H2L5186303 Suppressed the Elivated Eosinophil and Lymphocyte Levels
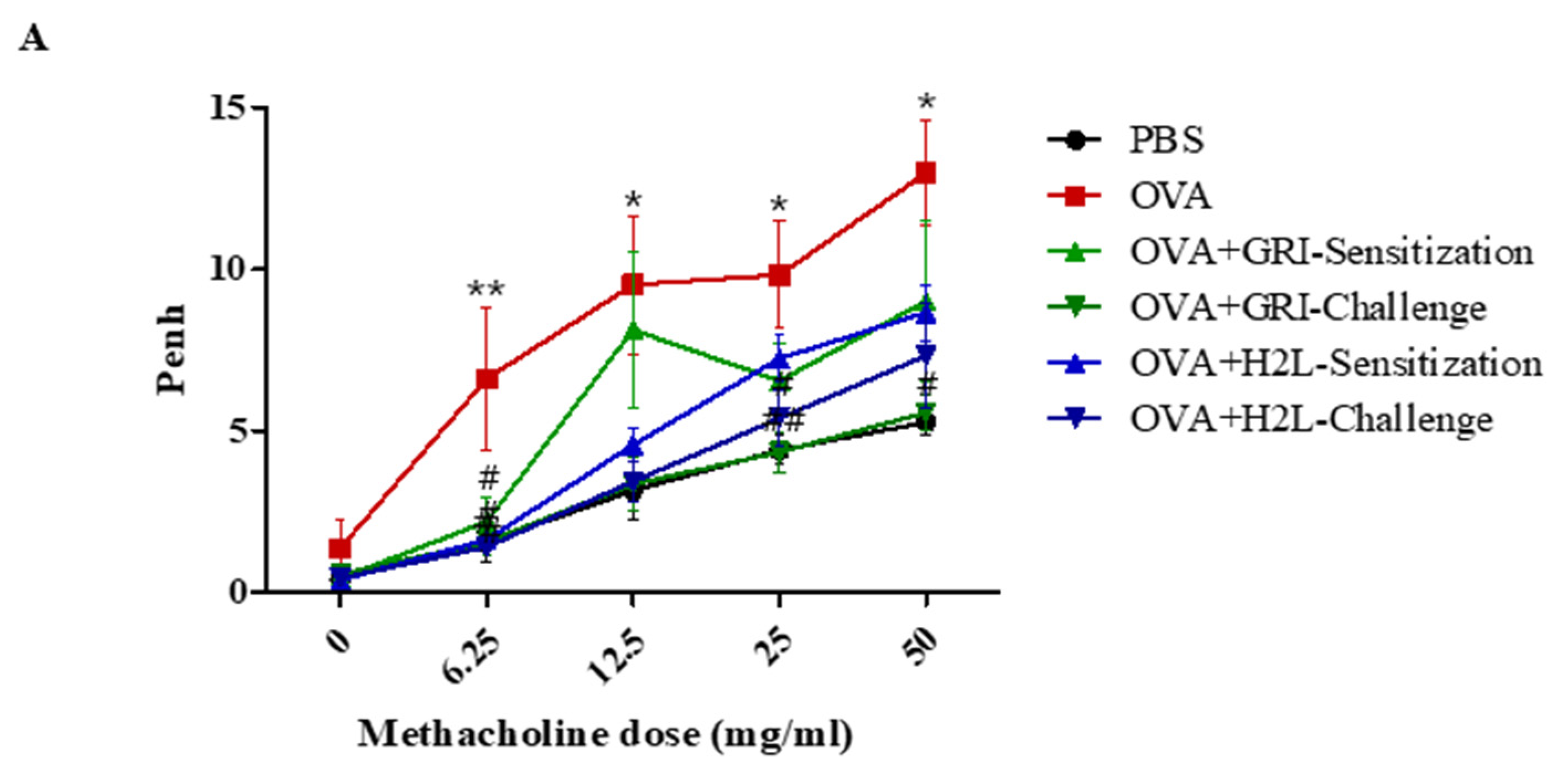
2.3. GRI977143 and H2L5186303 Suppressed AHR
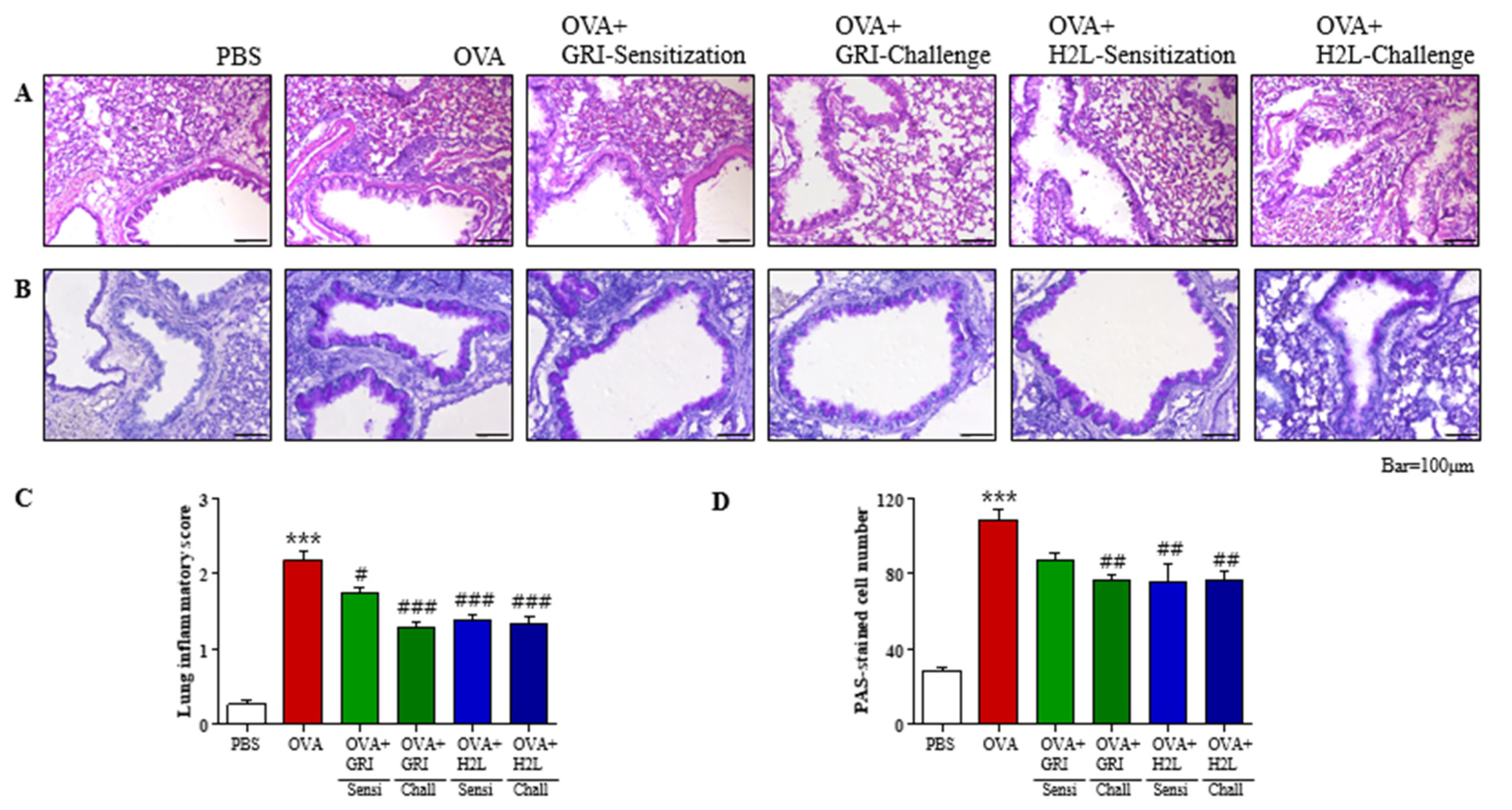
2.4. GRI977143 and H2L5186303 Suppressed Mucin Secretion and Lung Inflammation
2.5. GRI977143 and H2L5186303 Suppressed the Production of Th2 and Th1 Cytokines in the BALF and Lungs
2.6. GRI977143 and H2L5186303 Treatments Suppressed OVA-Induced Elevated IL-13 Levels in BALF Unlike Serum IgE Levels
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Animals
4.4. Assessment of Degranulation
4.5. Asthma Induction in Mice and Administration of H2L5186303 and GRI977143
4.6. BALF Cell Counting and Analysis
4.7. Histological Examination of the Lung Tissues
4.8. Measurement of Airway Hyperresponsiveness
4.9. Measurement of Total Serum IgE and IL-13 Level
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bereda, G. Bronchial Asthma: Etiology, Pathophysiology, Diagnosis and Management. Austin J. Pulm. Respir. Med. 2022, 9, 1085. [Google Scholar]
- Gauvreau, G.M.; Davis, B.E.; Scadding, G.; Boulet, L.-P.; Bjermer, L.; Chaker, A.; Cockcroft, D.W.; Dahlén, B.; Fokkens, W.; Hellings, P. Allergen Provocation Tests in Respiratory Research: Building on 50 Years of Experience. Eur. Respir. J. 2022, 60, 2102782. [Google Scholar] [CrossRef]
- Georas, S.; Berdyshev, E.; Hubbard, W.; Gorshkova, I.; Usatyuk, P.; Saatian, B.; Myers, A.; Williams, M.; Xiao, H.; Liu, M. Lysophosphatidic acid is detectable in human bronchoalveolar lavage fluids at baseline and increased after segmental allergen challenge. Clin. Exp. Allergy 2007, 37, 311–322. [Google Scholar] [CrossRef]
- Park, G.Y.; Lee, Y.G.; Berdyshev, E.; Nyenhuis, S.; Du, J.; Fu, P.; Gorshkova, I.A.; Li, Y.; Chung, S.; Karpurapu, M. Autotaxin production of lysophosphatidic acid mediates allergic asthmatic inflammation. Am. J. Respir. Crit. Care Med. 2013, 188, 928–940. [Google Scholar] [CrossRef]
- Kondo, M.; Tezuka, T.; Ogawa, H.; Koyama, K.; Bando, H.; Azuma, M.; Nishioka, Y. Lysophosphatidic acid regulates the differentiation of Th2 cells and its antagonist suppresses allergic airway inflammation. Int. Arch. Allergy Immunol. 2021, 182, 1–13. [Google Scholar] [CrossRef]
- Hashimoto, T.; Nakano, Y.; Yamashita, M.; Fang, Y.-I.; Ohata, H.; Momose, K. Role of Rho-associated protein kinase and histamine in lysophosphatidic acid-induced airway hyperresponsiveness in guinea pigs. Jpn. J. Pharmacol. 2002, 88, 256–261. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yamashita, M.; Ohata, H.; Momose, K. Lysophosphatidic acid enhances in vivo infiltration and activation of guinea pig eosinophils and neutrophils via a Rho/Rho-associated protein kinase-mediated pathway. J. Pharmacol. Sci. 2003, 91, 8–14. [Google Scholar] [CrossRef]
- Idzko, M.; Laut, M.; Panther, E.; Sorichter, S.; Dürk, T.; Fluhr, J.W.; Herouy, Y.; Mockenhaupt, M.; Myrtek, D.; Elsner, P. Lysophosphatidic acid induces chemotaxis, oxygen radical production, CD11b up-regulation, Ca2+ mobilization, and actin reorganization in human eosinophils via pertussis toxin-sensitive G proteins. J. Immunol. 2004, 172, 4480–4485. [Google Scholar]
- Hashimoto, T.; Ohata, H.; Momose, K.; Honda, K. Lysophosphatidic acid induces histamine release from mast cells and skin fragments. Pharmacology 2005, 75, 13–20. [Google Scholar] [CrossRef]
- Lin, D.A.; Boyce, J.A. IL-4 regulates MEK expression required for lysophosphatidic acid-mediated chemokine generation by human mast cells. J. Immunol. 2005, 175, 5430–5438. [Google Scholar] [CrossRef]
- Rubenfeld, J.; Guo, J.; Sookrung, N.; Chen, R.; Chaicumpa, W.; Casolaro, V.; Zhao, Y.; Natarajan, V.; Georas, S. Lysophosphatidic acid enhances interleukin-13 gene expression and promoter activity in T cells. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2006, 290, L66–L74. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Knudsen, E.; Jin, Y.; Gessani, S.; Maghazachi, A.A. Lysophospholipids and chemokines activate distinct signal transduction pathways in T helper 1 and T helper 2 cells. Cell. Signal. 2004, 16, 991–1000. [Google Scholar] [CrossRef]
- Panther, E.; Idzko, M.; Corinti, S.; Ferrari, D.; Herouy, Y.; Mockenhaupt, M.; Dichmann, S.; Gebicke-Haerter, P.; Di Virgilio, F.; Girolomoni, G. The influence of lysophosphatidic acid on the functions of human dendritic cells. J. Immunol. 2002, 169, 4129–4135. [Google Scholar] [CrossRef]
- Chan, L.C.; Peters, W.; Xu, Y.; Chun, J.; Farese Jr, R.V.; Cases, S. LPA3 receptor mediates chemotaxis of immature murine dendritic cells to unsaturated lysophosphatidic acid (LPA). J. Leukoc. Biol. 2007, 82, 1193–1200. [Google Scholar] [CrossRef]
- Medoff, B.D.; Landry, A.L.; Wittbold, K.A.; Sandall, B.P.; Derby, M.C.; Cao, Z.; Adams, J.C.; Xavier, R.J. CARMA3 mediates lysophosphatidic acid–stimulated cytokine secretion by bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 2009, 40, 286–294. [Google Scholar] [CrossRef]
- Saatian, B.; Zhao, Y.; He, D.; Georas, S.N.; Watkins, T.; Spannhake, E.W.; Natarajan, V. Transcriptional regulation of lysophosphatidic acid-induced interleukin-8 expression and secretion by p38 MAPK and JNK in human bronchial epithelial cells. Biochem. J. 2006, 393, 657–668. [Google Scholar] [CrossRef]
- Zhao, Y.; Tong, J.; He, D.; Pendyala, S.; Evgeny, B.; Chun, J.; Sperling, A.I.; Natarajan, V. Role of lysophosphatidic acid receptor LPA 2 in the development of allergic airway inflammation in a murine model of asthma. Respir. Res. 2009, 10, 114. [Google Scholar] [CrossRef]
- Huang, L.S.; Fu, P.; Patel, P.; Harijith, A.; Sun, T.; Zhao, Y.; Garcia, J.G.; Chun, J.; Natarajan, V. Lysophosphatidic acid receptor–2 deficiency confers protection against bleomycin-induced lung injury and fibrosis in mice. Am. J. Respir. Cell Mol. Biol. 2013, 49, 912–922. [Google Scholar] [CrossRef]
- Emo, J.; Meednu, N.; Chapman, T.J.; Rezaee, F.; Balys, M.; Randall, T.; Rangasamy, T.; Georas, S.N. Lpa2 is a negative regulator of both dendritic cell activation and murine models of allergic lung inflammation. J. Immunol. 2012, 188, 3784–3790. [Google Scholar] [CrossRef]
- Fells, J.I.; Tsukahara, R.; Fujiwara, Y.; Liu, J.; Perygin, D.H.; Osborne, D.A.; Tigyi, G.; Parrill, A.L. Identification of non-lipid LPA3 antagonists by virtual screening. Bioorg. Med. Chem. 2008, 16, 6207–6217. [Google Scholar] [CrossRef]
- Knowlden, S.A.; Hillman, S.E.; Chapman, T.J.; Patil, R.; Miller, D.D.; Tigyi, G.; Georas, S.N. Novel inhibitory effect of a lysophosphatidic acid 2 agonist on allergen-driven airway inflammation. Am. J. Respir. Cell Mol. Biol. 2016, 54, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; He, D.; Zhao, J.; Wang, L.; Leff, A.R.; Spannhake, E.W.; Georas, S.; Natarajan, V. Lysophosphatidic acid induces interleukin-13 (IL-13) receptor α2 expression and inhibits IL-13 signaling in primary human bronchial epithelial cells. J. Biol. Chem. 2007, 282, 10172–10179. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Natarajan, V.; Stern, R.; Gorshkova, I.A.; Solway, J.; Spannhake, E.W.; Zhao, Y. Lysophosphatidic acid-induced transactivation of epidermal growth factor receptor regulates cyclo-oxygenase-2 expression and prostaglandin E2 release via C/EBPβ in human bronchial epithelial cells. Biochem. J. 2008, 412, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Cerutis, D.R.; Nogami, M.; Anderson, J.L.; Churchill, J.D.; Romberger, D.J.; Rennard, S.I.; Toews, M.L. Lysophosphatidic acid and EGF stimulate mitogenesis in human airway smooth muscle cells. Am. J. Physiol.-Lung Cell. Mol. Physiol. 1997, 273, L10–L15. [Google Scholar] [CrossRef] [PubMed]
- Toews, M.; Ustinova, E.; Schultz, H. Lysophosphatidic acid enhances contractility of isolated airway smooth muscle. J. Appl. Physiol. 1997, 83, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Im, D.S. Suppressive Effect of Carnosol on Ovalbumin-Induced Allergic Asthma. Biomol. Ther. 2021, 29, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Im, D.-S. 4-CMTB Ameliorates Ovalbumin-Induced Allergic Asthma through FFA2 Activation in Mice. Biomol. Ther. 2021, 29, 427. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-J.; Im, D.-S. Efficacy Comparison of LPA2 Antagonist H2L5186303 and Agonist GRI977143 on Ovalbumin-Induced Allergic Asthma in BALB/c Mice. Int. J. Mol. Sci. 2022, 23, 9745. https://doi.org/10.3390/ijms23179745
Lee Y-J, Im D-S. Efficacy Comparison of LPA2 Antagonist H2L5186303 and Agonist GRI977143 on Ovalbumin-Induced Allergic Asthma in BALB/c Mice. International Journal of Molecular Sciences. 2022; 23(17):9745. https://doi.org/10.3390/ijms23179745
Chicago/Turabian StyleLee, Ye-Ji, and Dong-Soon Im. 2022. "Efficacy Comparison of LPA2 Antagonist H2L5186303 and Agonist GRI977143 on Ovalbumin-Induced Allergic Asthma in BALB/c Mice" International Journal of Molecular Sciences 23, no. 17: 9745. https://doi.org/10.3390/ijms23179745
APA StyleLee, Y.-J., & Im, D.-S. (2022). Efficacy Comparison of LPA2 Antagonist H2L5186303 and Agonist GRI977143 on Ovalbumin-Induced Allergic Asthma in BALB/c Mice. International Journal of Molecular Sciences, 23(17), 9745. https://doi.org/10.3390/ijms23179745