
Dynamic Changes in the Endocannabinoid System during the Aging Process: Focus on the Middle-Age Crisis



 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Levels of 2-Archidonoylglycerol and Anandamide Fluctuate in an Age- and Region-Specific Manner
2.2. Prominent Reduction in Hippocampal DAGLα during the Early Middle-Age
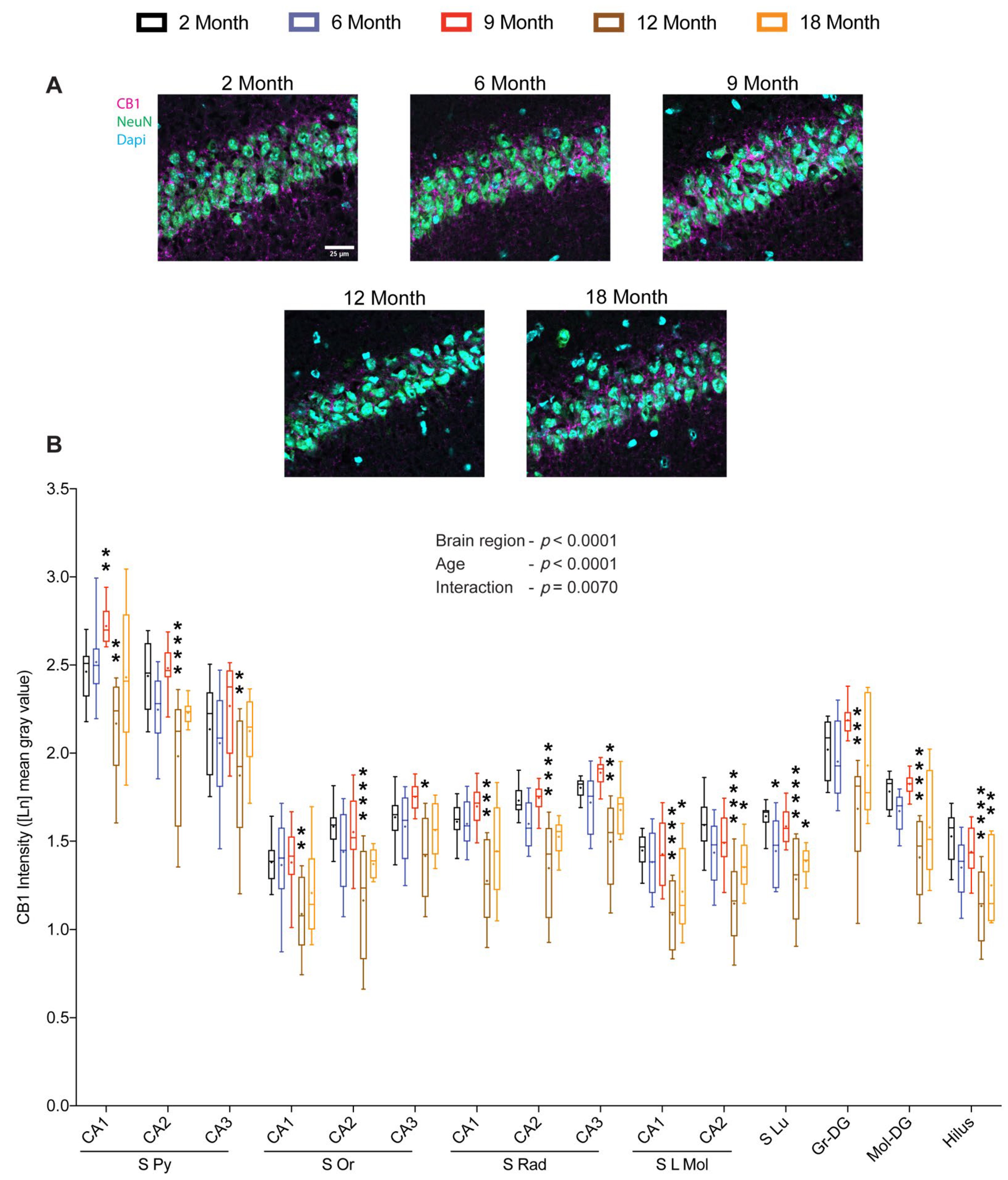
2.3. Density of Hippocampal CB1 Increases during the Early Middle-Age
2.4. DAGLα and CB1 Are Reduced with Age and in a Brain Region-Specific Manner in the Limbic Forebrain
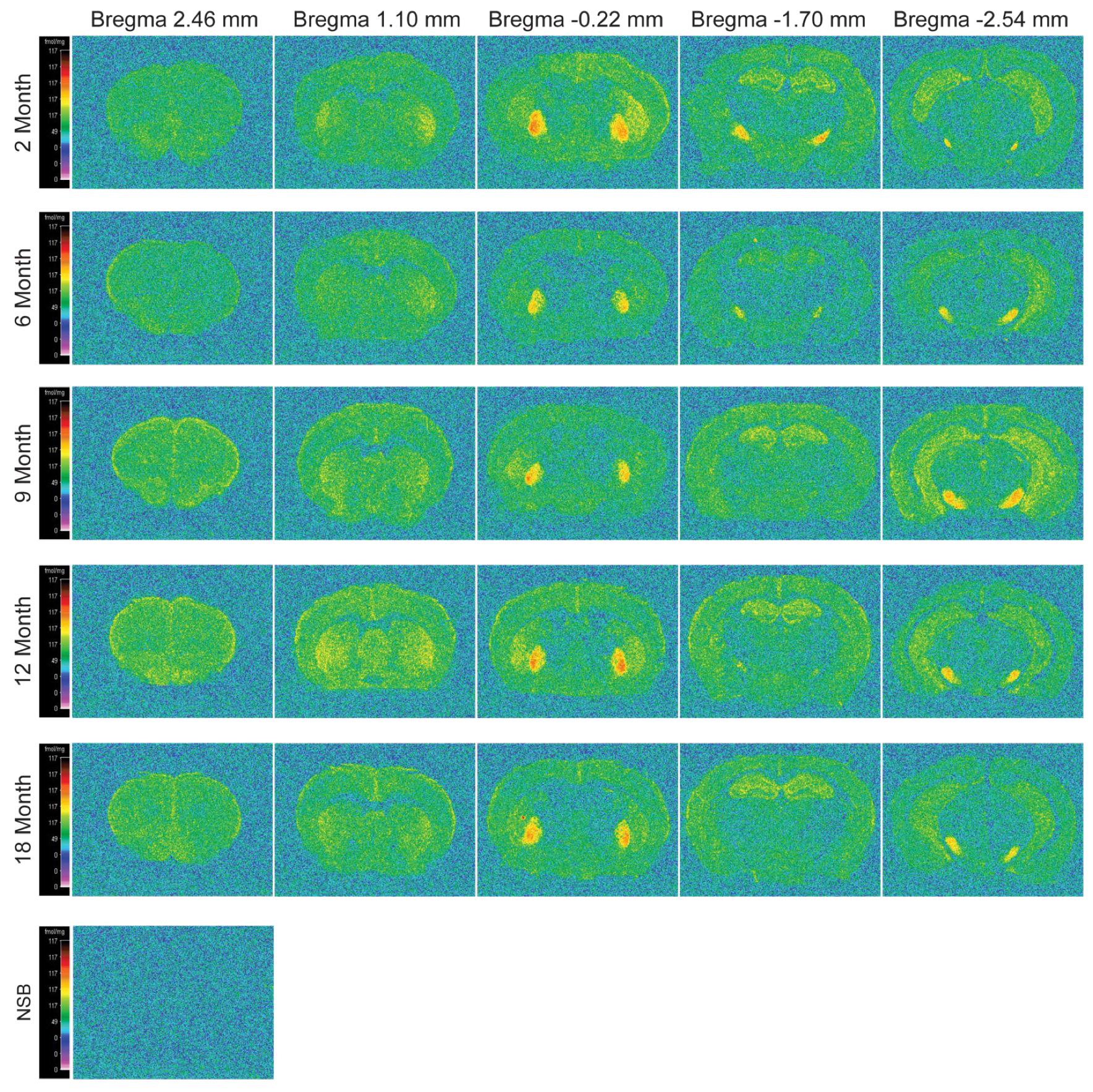
2.5. Alterations in CB1-Specific Binding of [3H] CP55940 across Brain Regions
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Endocannabinoids Extraction
4.3. LC/MS Quantitative Analyses
4.4. Immunofluorescence
4.5. Autoradiographic Binding of CB1 Receptor
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Marzo, V.; Stella, N.; Zimmer, A. Endocannabinoid Signalling and the Deteriorating Brain. Nat. Rev. Neurosci. 2015, 16, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M. Missing Pieces to the Endocannabinoid Puzzle. Trends Mol. Med. 2019, 26, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Jenniches, I.; Ternes, S.; Albayram, O.; Otte, D.M.; Bach, K.; Bindila, L.; Michel, K.; Lutz, B.; Bilkei-Gorzo, A.; Zimmer, A. Anxiety, Stress, and Fear Response in Mice With Reduced Endocannabinoid Levels. Biol. Psychiatry 2016, 79, 858–868. [Google Scholar] [CrossRef]
- Maccarrone, M.; Guzmán, M.; Mackie, K.; Doherty, P.; Harkany, T. Programming of Neural Cells by (Endo)Cannabinoids: From Physiological Rules to Emerging Therapies. Nat. Rev. Neurosci. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Meyer, H.C.; Lee, F.S.; Gee, D.G. The Role of the Endocannabinoid System and Genetic Variation in Adolescent Brain Development. Neuropsychopharmacology 2018, 43, 21–33. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A.; Racz, I.; Valverde, O.; Otto, M.; Michel, K.; Sastre, M.; Sarstre, M.; Zimmer, A. Early Age-Related Cognitive Impairment in Mice Lacking Cannabinoid CB1 Receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 15670–15675. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A.; Drews, E.; Albayram, Ö.; Piyanova, A.; Gaffal, E.; Tueting, T.; Michel, K.; Mauer, D.; Maier, W.; Zimmer, A. Early Onset of Aging-like Changes Is Restricted to Cognitive Abilities and Skin Structure in Cnr1−/− Mice. Neurobiol. Aging 2012, 33, e11–e200. [Google Scholar] [CrossRef] [PubMed]
- Albayram, O.; Alferink, J.; Pitsch, J.; Piyanova, A.; Neitzert, K.; Poppensieker, K.; Mauer, D.; Michel, K.; Legler, A.; Becker, A.; et al. Role of CB1 Cannabinoid Receptors on GABAergic Neurons in Brain Aging. Proc. Natl. Acad. Sci. USA 2011, 108, 11256–11261. [Google Scholar] [CrossRef]
- Piyanova, A.; Lomazzo, E.; Bindila, L.; Lerner, R.; Albayram, O.; Ruhl, T.; Lutz, B.; Zimmer, A.; Bilkei-Gorzo, A. Age-Related Changes in the Endocannabinoid System in the Mouse Hippocampus. Mech. Ageing Dev. 2015, 150, 55–64. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A.; Albayram, O.; Draffehn, A.; Michel, K.; Piyanova, A.; Oppenheimer, H.; Dvir-Ginzberg, M.; Rácz, I.; Ulas, T.; Imbeault, S.; et al. A Chronic Low Dose of Δ9-Tetrahydrocannabinol (THC) Restores Cognitive Function in Old Mice. Nat. Med. 2017, 23, 782–787. [Google Scholar] [CrossRef]
- Nidadavolu, P.; Bilkei-Gorzo, A.; Krämer, M.; Schürmann, B.; Palmisano, M.; Beins, E.C.; Madea, B.; Zimmer, A. Efficacy of Δ9-Tetrahydrocannabinol (THC) Alone or in Combination With a 1:1 Ratio of Cannabidiol (CBD) in Reversing the Spatial Learning Deficits in Old Mice. Front. Aging Neurosci. 2021, 13, 718850. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.; Mills, S.; Winstone, J.; Leishman, E.; Wager-Miller, J.; Bradshaw, H.; Mackie, K. Chronic Adolescent Δ9-Tetrahydrocannabinol Treatment of Male Mice Leads to Long-Term Cognitive and Behavioral Dysfunction, Which Are Prevented by Concurrent Cannabidiol Treatment. Cannabis Cannabinoid Res. 2017, 2, 235–246. [Google Scholar] [CrossRef]
- Solowij, N.; Stephens, R.S.; Roffman, R.A.; Babor, T.; Kadden, R.; Miller, M.; Christiansen, K.; McRee, B.; Vendetti, J. Marijuana Treatment Project Research Group Cognitive Functioning of Long-Term Heavy Cannabis Users Seeking Treatment. JAMA 2002, 287, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Harvey, M.A.; Sellman, J.D.; Porter, R.J.; Frampton, C.M. The Relationship between Non-Acute Adolescent Cannabis Use and Cognition. Drug Alcohol Rev. 2007, 26, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, D.M.; Mathias, C.W.; Dawes, M.A.; Furr, R.M.; Charles, N.E.; Liguori, A.; Shannon, E.E.; Acheson, A. Impulsivity, Attention, Memory, and Decision-Making among Adolescent Marijuana Users. Psychopharmacology 2013, 226, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Feliszek, M.; Bindila, L.; Lutz, B.; Zimmer, A.; Bilkei-Gorzo, A.; Schlicker, E. Lack of Hippocampal CB1 Receptor Desensitization by Δ9-Tetrahydrocannabinol in Aged Mice and by Low Doses of JZL 184. Naunyn. Schmiedebergs Arch. Pharmacol. 2016, 389, 603–612. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Harvey-White, J.; Zimmer, A.; Kunos, G. Endocannabinoid Signaling via Cannabinoid Receptor 1 Is Involved in Ethanol Preference and Its Age-Dependent Decline in Mice. Proc. Natl. Acad. Sci. USA 2003, 100, 1393–1398. [Google Scholar] [CrossRef]
- Jackson, S.J.; Andrews, N.; Ball, D.; Bellantuono, I.; Gray, J.; Hachoumi, L.; Holmes, A.; Latcham, J.; Petrie, A.; Potter, P.; et al. Does Age Matter? The Impact of Rodent Age on Study Outcomes. Lab. Anim. 2017, 51, 160–169. [Google Scholar] [CrossRef]
- Flurkey, K.; Mcurrer, J.; Harrison, D. Mouse Models in Aging Research. In The Mouse in Biomedical Research; Elsevier: Amsterdam, The Netherlands, 2007; Volume 3, pp. 637–672. ISBN 9780123694546. [Google Scholar]
- Lehallier, B.; Gate, D.; Schaum, N.; Nanasi, T.; Lee, S.E.; Yousef, H.; Moran Losada, P.; Berdnik, D.; Keller, A.; Verghese, J.; et al. Undulating Changes in Human Plasma Proteome Profiles across the Lifespan. Nat. Med. 2019, 25, 1843–1850. [Google Scholar] [CrossRef] [PubMed]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Age-Related Changes in Behavior in C57BL/6J Mice from Young Adulthood to Middle Age. Mol. Brain 2016, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Hendrickx, J.O.; De Moudt, S.; Calus, E.; De Deyn, P.P.; Van Dam, D.; De Meyer, G.R.Y. Age-Related Cognitive Decline in Spatial Learning and Memory of C57BL/6J Mice. Behav. Brain Res. 2022, 418, 113649. [Google Scholar] [CrossRef] [PubMed]
- Kaczorowski, C.C.; Disterhoft, J.F. Memory Deficits Are Associated with Impaired Ability to Modulate Neuronal Excitability in Middle-Aged Mice. Learn. Mem. 2009, 16, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Moretti, M.; de Souza, A.G.; de Chaves, G.; de Andrade, V.M.; Romao, P.R.T.; Gavioli, E.C.; Boeck, C.R. Emotional Behavior in Middle-Aged Rats: Implications for Geriatric Psychopathologies. Physiol. Behav. 2011, 102, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Bilkei-Gorzo, A. The Endocannabinoid System in Normal and Pathological Brain Ageing. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3326–3341. [Google Scholar] [CrossRef]
- Marcaggi, P. Cerebellar Endocannabinoids: Retrograde Signaling from Purkinje Cells. Cerebellum 2015, 14, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, S.K.; King, C.; Kortleven, C.; Huson, V.; Kroon, T.; Kevenaar, J.T.; Schut, D.; Saarloos, I.; Hoetjes, J.P.; de Wit, H.; et al. Presynaptic Inhibition upon CB1 or MGlu2/3 Receptor Activation Requires ERK/MAPK Phosphorylation of Munc18-1. EMBO J. 2016, 35, 1236–1250. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Jia, Y.; Pham, D.T.; Palmer, L.C.; Jung, K.-M.; Cox, C.D.; Rumbaugh, G.; Piomelli, D.; Gall, C.M.; Lynch, G. Atypical Endocannabinoid Signaling Initiates a New Form of Memory-Related Plasticity at a Cortical Input to Hippocampus. Cereb. Cortex 2018, 28, 2253–2266. [Google Scholar] [CrossRef]
- Wang, W.; Trieu, B.H.; Palmer, L.C.; Jia, Y.; Pham, D.T.; Jung, K.-M.; Karsten, C.A.; Merrill, C.B.; Mackie, K.; Gall, C.M.; et al. A Primary Cortical Input to Hippocampus Expresses a Pathway-Specific and Endocannabinoid-Dependent Form of Long-Term Potentiation. eNeuro 2016, 3, ENEURO.0160-16.2016. [Google Scholar] [CrossRef] [PubMed]
- Amani, M.; Lauterborn, J.C.; Le, A.A.; Cox, B.M.; Wang, W.; Quintanilla, J.; Cox, C.D.; Gall, C.M.; Lynch, G. Rapid Aging in the Perforant Path Projections to the Rodent Dentate Gyrus. J. Neurosci. 2021, 41, 2301–2312. [Google Scholar] [CrossRef] [PubMed]
- Shonesy, B.C.; Parrish, W.P.; Haddad, H.K.; Stephenson, J.R.; Báldi, R.; Bluett, R.J.; Marks, C.R.; Centanni, S.W.; Folkes, O.M.; Spiess, K.; et al. Role of Striatal Direct Pathway 2-Arachidonoylglycerol Signaling in Sociability and Repetitive Behavior. Biol. Psychiatry 2018, 84, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, J.; Berrendero, F.; Garcia-Gil, L.; de la Cruz, P.; Ramos, J.A.; Fernández-Ruiz, J.J. Loss of Cannabinoid Receptor Binding and Messenger RNA Levels and Cannabinoid Agonist-Stimulated [35S]Guanylyl-5’O-(Thio)-Triphosphate Binding in the Basal Ganglia of Aged Rats. Neuroscience 1998, 84, 1075–1083. [Google Scholar] [CrossRef]
- Berrendero, F.; Romero, J.; García-Gil, L.; Suarez, I.; De la Cruz, P.; Ramos, J.A.; Fernández-Ruiz, J.J. Changes in Cannabinoid Receptor Binding and MRNA Levels in Several Brain Regions of Aged Rats. Biochim. Biophys. Acta 1998, 1407, 205–214. [Google Scholar] [CrossRef]
- Liu, P.; Bilkey, D.K.; Darlington, C.L.; Smith, P.F. Cannabinoid CB1 Receptor Protein Expression in the Rat Hippocampus and Entorhinal, Perirhinal, Postrhinal and Temporal Cortices: Regional Variations and Age-Related Changes. Brain Res. 2003, 979, 235–239. [Google Scholar] [CrossRef]
- Dalle, S.; Koppo, K. Cannabinoid Receptor 1 Expression Is Higher in Muscle of Old vs. Young Males, and Increases upon Resistance Exercise in Older Adults. Sci. Rep. 2021, 11, 18349. [Google Scholar] [CrossRef]
- Hill, M.N.; Titterness, A.K.; Morrish, A.C.; Carrier, E.J.; Lee, T.T.-Y.; Gil-Mohapel, J.; Gorzalka, B.B.; Hillard, C.J.; Christie, B.R. Endogenous Cannabinoid Signaling Is Required for Voluntary Exercise-Induced Enhancement of Progenitor Cell Proliferation in the Hippocampus. Hippocampus 2010, 20, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Galdino, G.; Romero, T.R.L.; Silva, J.F.P.; Aguiar, D.C.; de Paula, A.M.; Cruz, J.S.; Parrella, C.; Piscitelli, F.; Duarte, I.D.; Di Marzo, V.; et al. The Endocannabinoid System Mediates Aerobic Exercise-Induced Antinociception in Rats. Neuropharmacology 2014, 77, 313–324. [Google Scholar] [CrossRef]
- Pamplona, F.A.; Ferreira, J.; Menezes de Lima, O.; Duarte, F.S.; Bento, A.F.; Forner, S.; Villarinho, J.G.; Bellocchio, L.; Bellochio, L.; Wotjak, C.T.; et al. Anti-Inflammatory Lipoxin A4 Is an Endogenous Allosteric Enhancer of CB1 Cannabinoid Receptor. Proc. Natl. Acad. Sci. USA 2012, 109, 21134–21139. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Lipoxin A4 Is an Allosteric Endocannabinoid That Strengthens Anandamide-Induced CB1 Receptor Activation. Proc. Natl. Acad. Sci. USA 2012, 109, 20781–20782. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, H.; McCaul, M.E.; Wand, G.S.; Earley, C.J.; Allen, R.P.; Weerts, E.M.; Dannals, R.F.; Wong, D.F. Dissociative Changes in the Bmax and KD of Dopamine D2/D3 Receptors with Aging Observed in Functional Subdivisions of the Striatum: A Revisit with an Improved Data Analysis Method. J. Nucl. Med. 2012, 53, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Steindel, F.; Lerner, R.; Häring, M.; Ruehle, S.; Marsicano, G.; Lutz, B.; Monory, K. Neuron-Type Specific Cannabinoid-Mediated G Protein Signalling in Mouse Hippocampus. J. Neurochem. 2013, 124, 795–807. [Google Scholar] [CrossRef]
- Lerner, R.; Post, J.M.; Ellis, S.R.; Vos, D.R.N.; Heeren, R.M.A.; Lutz, B.; Bindila, L. Simultaneous Lipidomic and Transcriptomic Profiling in Mouse Brain Punches of Acute Epileptic Seizure Model Compared to Controls. J. Lipid Res. 2018, 59, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Berrendero, F.; Mendizábal, V.; Murtra, P.; Kieffer, B.L.; Maldonado, R. Cannabinoid Receptor and WIN 55 212-2-Stimulated [35S]-GTPgammaS Binding in the Brain of Mu-, Delta- and Kappa-Opioid Receptor Knockout Mice. Eur. J. Neurosci. 2003, 18, 2197–2202. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, I.; Slowe, S.J.; Matthes, H.W.; Kieffer, B. Quantitative Autoradiographic Mapping of Mu-, Delta- and Kappa-Opioid Receptors in Knockout Mice Lacking the Mu-Opioid Receptor Gene. Brain Res. 1997, 778, 73–88. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Groups | ||||||
|---|---|---|---|---|---|---|
| Region | Bregma (mm) | 2 Month | 6 Month | 9 Month | 12 Month | 18 Month |
| M1 + M2 | +2.46 | 70.53 ± 4.47 | 65.68 ± 7.71 | 78.99 ± 8.09 | 74.03 ± 8.18 | 78.62 ± 8.57 |
| LO + VO | +2.46 | 65.16 ± 4.67 | 63.45 ± 6.71 | 71.82 ± 5.59 | 69.89 ± 6.49 | 71.89 ± 7.16 |
| AON | +2.46 | 82.67 ± 5.59 | 75.04 ± 9.33 | 89.84 ± 8.49 | 88.57 ± 6.93 | 88.50 ± 7.86 |
| PrL | +2.46 | 75.47 ± 4.8 | 68.89 ± 8.81 | 82.83 ± 8.26 | 78.84 ± 8.65 | 82.89 ± 8.51 |
| M1 + M2 | +1.10 | 72.79 ± 4.86 | 67.65 ± 5.77 | 83.97 ± 6.19 | 80.83 ± 6.93 | 82.04 ± 7.69 |
| S1 | +1.10 | 62.84 ± 4.84 | 58.31 ± 5.16 | 68.54 ± 4.51 | 64.85 ± 6.32 | 69.15 ± 6.78 |
| Cg1 + Cg2 | +1.10 | 76.20 ± 4.93 | 69.36 ± 7.00 | 85.32 ± 6.27 | 84.04 ± 7.76 | 87.87 ± 9.12 |
| Cpu | +1.10 | 92.17 ± 4.35 | 79.79 ± 6.52 | 91.20 ± 6.49 | 84.91 ± 9.06 | 84.97 ± 8.2 |
| AcbSh | +1.10 | 81.79 ± 9.01 | 65.55 ± 5.33 | 79.83 ± 6.83 | 83.74 ± 7.57 | 74.87 ± 7.40 |
| AcbC | +1.10 | 88.73 ± 10.18 | 73.92 ± 7.56 | 88.74 ± 10.29 | 87.69 ± 7.49 | 83.00 ± 9.32 |
| DEn | +1.10 | 69.02 ± 4.61 | 60.56 ± 3.75 | 73.67 ± 5.18 | 81.51 ± 6.95 | 78.72 ± 7.0 |
| LS | +1.10 | 74.66 ± 5.15 | 67.10 ± 6.47 | 83.24 ± 8.43 | 82.31 ± 8.78 | 81.99 ± 7.84 |
| MS | +1.10 | 73.20 ± 7.61 | 62.68 ± 4.72 | 73.88 ± 6.35 | 77.96 ± 9.95 | 70.53 ± 7.76 |
| Tu | +1.10 | 71.23 ± 5.42 | 58.49 ± 3.97 | 72.06 ± 5.9 | 71.30 ± 7.45 | 67.05 ± 7.18 |
| PGP | −0.22 | 150.12 ± 10.53 | 127.42 ± 10.82 ** | 144.26 ± 7.22 | 145.05 ± 11.2 | 135.72 ± 8.98 |
| CA1 | −1.70 | 117.27 ± 6.18 | 117.33 ± 7.28 | 138.41 ± 8.39 * | 126.16 ± 11.49 | 126.99 ± 10.38 |
| CA2 | −1.70 | 119.58 ± 5.8 | 123.30 ± 9.9 | 140.8 ± 6.9 * | 129.09 ± 11.56 | 135.00 ± 10.42 |
| CA3 | −1.70 | 129.31 ± 7.40 | 138.34 ± 12.30 | 155.03 ± 8.62 ** | 138.37 ± 12.21 | 138.82 ± 11.18 |
| DG | −1.70 | 129.00 ± 8.24 | 130.27 ± 10.93 | 146.28 ± 8.3 a | 134.75 ± 13.81 | 135.34 ± 11.4 |
| Hy | −1.70 | 58.29 ± 4.92 | 51.24 ± 3.84 | 54.89 ± 3.37 | 55.41 ± 6.92 | 53.83 ± 6.04 |
| Amy | −1.70 | 56.29 ± 3.1 | 53.74 ± 4.54 | 62.95 ± 4.30 | 65.14 ± 7.03 | 65.24 ± 6.75 |
| BLA | −1.70 | 60.31 ± 4.65 | 56.21 ± 4.56 | 65.32 ± 3.77 | 71.64 ± 8.21 | 66.34 ± 6.58 |
| BMA | −1.70 | 53.77 ± 3.37 | 50.74 ± 5.21 | 58.33 ± 5.55 | 58.65 ± 6.08 | 62.27 ± 7.21 |
| Pir | −1.70 | 53.43 ± 3.88 | 50.01 ± 4.07 | 58.84 ± 4.51 | 62.80 ± 5.8 | 63.92 ± 7.17 |
| S1 + S2 | −1.70 | 57.68 ± 4.28 | 56.02 ± 4.10 | 66.59 ± 3.43 | 67.02 ± 7.60 | 65.76 ± 7.12 |
| Th | −1.70 | 38.63 ± 4.30 | 34.26 ± 3.11 | 40.13 ± 3.56 | 37.04 ± 4.47 | 38.34 ± 5.49 |
| SNC + SNR | −2.54 | 130.56 ± 10.19 | 135.85 ± 8.34 | 132.91 ± 13.69 | 98.15 ± 17.05 | 105.87 ± 14.11 ** |
| PAG | −2.54 | 62.23 ± 4.82 | 61.40 ± 3.94 | 68.80 ± 6.15 | 54.64 ± 4.93 | 59.21 ± 7.65 |
| Hip | −2.54 | 80.25 ± 4.48 | 87.39 ± 4.99 | 102.23 ± 5.49 ** | 82.47 ± 8.14 | 93.38 ± 6.22 |
| LEnt | −2.54 | 55.31 ± 2.38 | 60.17 ± 5.14 | 72.36 ± 4.69 * | 74.75 ± 5.34 * | 73.27 ± 5.84 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nidadavolu, P.; Bilkei-Gorzo, A.; Effah, F.; Leidmaa, E.; Schürmann, B.; Berger, M.; Bindila, L.; Schmid, M.; Lutz, B.; Zimmer, A.; et al. Dynamic Changes in the Endocannabinoid System during the Aging Process: Focus on the Middle-Age Crisis. Int. J. Mol. Sci. 2022, 23, 10254. https://doi.org/10.3390/ijms231810254
Nidadavolu P, Bilkei-Gorzo A, Effah F, Leidmaa E, Schürmann B, Berger M, Bindila L, Schmid M, Lutz B, Zimmer A, et al. Dynamic Changes in the Endocannabinoid System during the Aging Process: Focus on the Middle-Age Crisis. International Journal of Molecular Sciences. 2022; 23(18):10254. https://doi.org/10.3390/ijms231810254
Chicago/Turabian StyleNidadavolu, Prakash, Andras Bilkei-Gorzo, Felix Effah, Este Leidmaa, Britta Schürmann, Moritz Berger, Laura Bindila, Matthias Schmid, Beat Lutz, Andreas Zimmer, and et al. 2022. "Dynamic Changes in the Endocannabinoid System during the Aging Process: Focus on the Middle-Age Crisis" International Journal of Molecular Sciences 23, no. 18: 10254. https://doi.org/10.3390/ijms231810254
APA StyleNidadavolu, P., Bilkei-Gorzo, A., Effah, F., Leidmaa, E., Schürmann, B., Berger, M., Bindila, L., Schmid, M., Lutz, B., Zimmer, A., & Bailey, A. (2022). Dynamic Changes in the Endocannabinoid System during the Aging Process: Focus on the Middle-Age Crisis. International Journal of Molecular Sciences, 23(18), 10254. https://doi.org/10.3390/ijms231810254