
A Metagenomic and Gene Expression Analysis in Wheat (T. durum) and Maize (Z. mays) Biofertilized with PGPM and Biochar

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Char Has a High Cargo Capacity for Microbes
2.2. Effects of PGPM and Char on Plant Growth
2.2.1. Wheat
2.2.2. Maize
2.3. Analysis of Rhizosphere Microbiota and Mycobiota
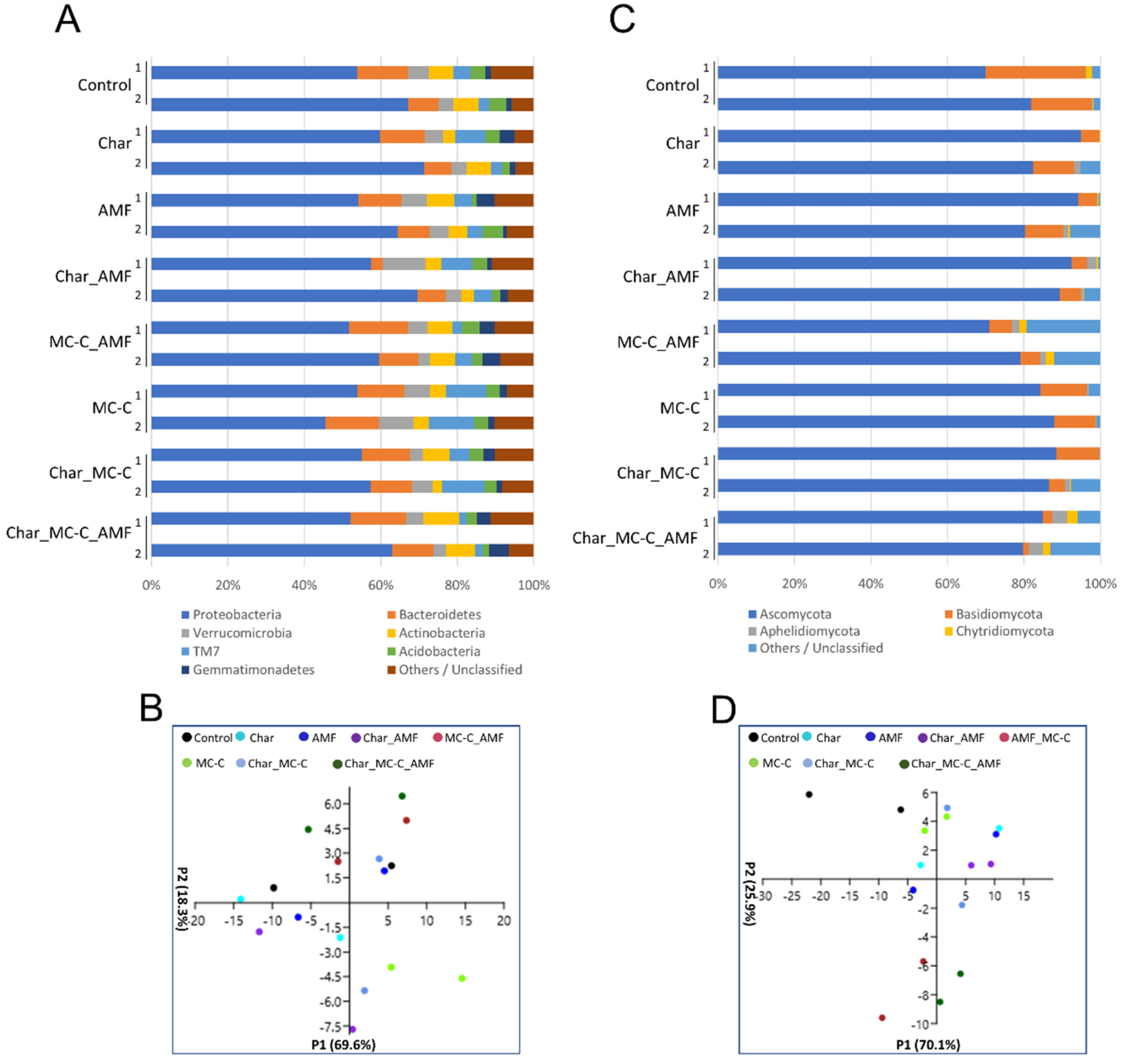
2.3.1. Bacteria and Fungi in the Rhizosphere of Wheat
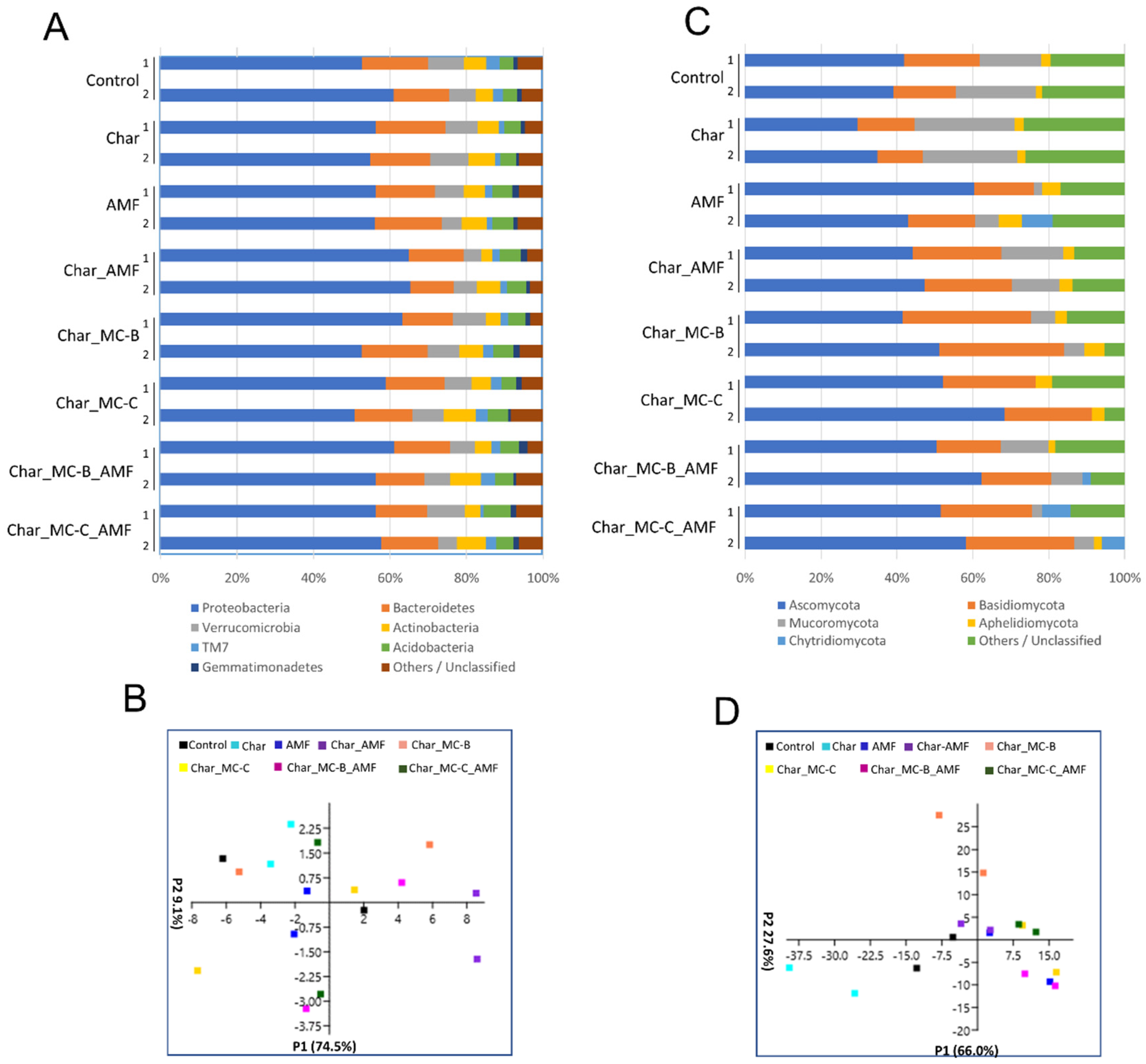
2.3.2. Bacteria and Fungi in the Rhizosphere of Maize
2.4. Gene Expression in Wheat and Maize Leaves (at Two Different Growth Stages)
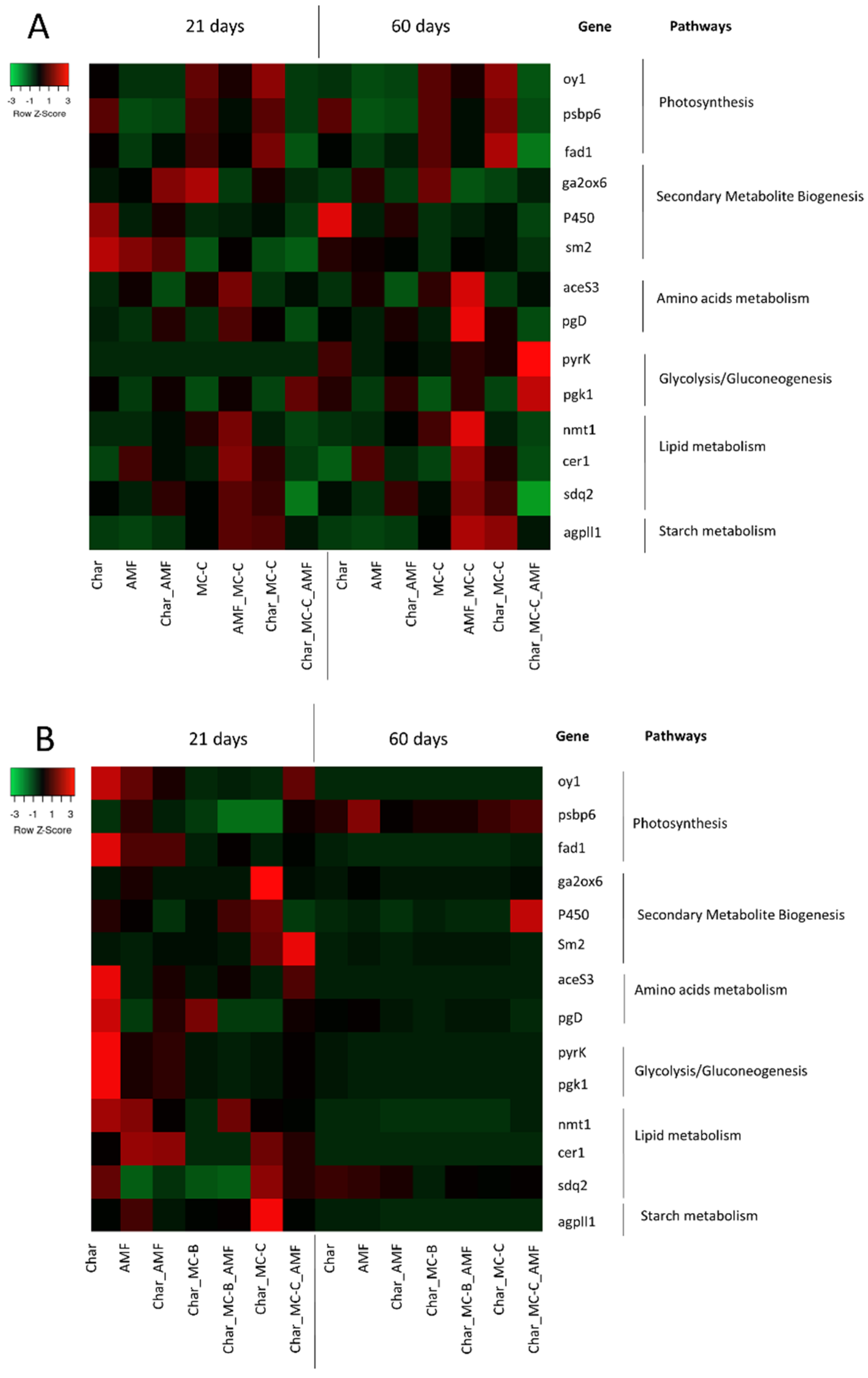
2.4.1. Wheat
2.4.2. Maize
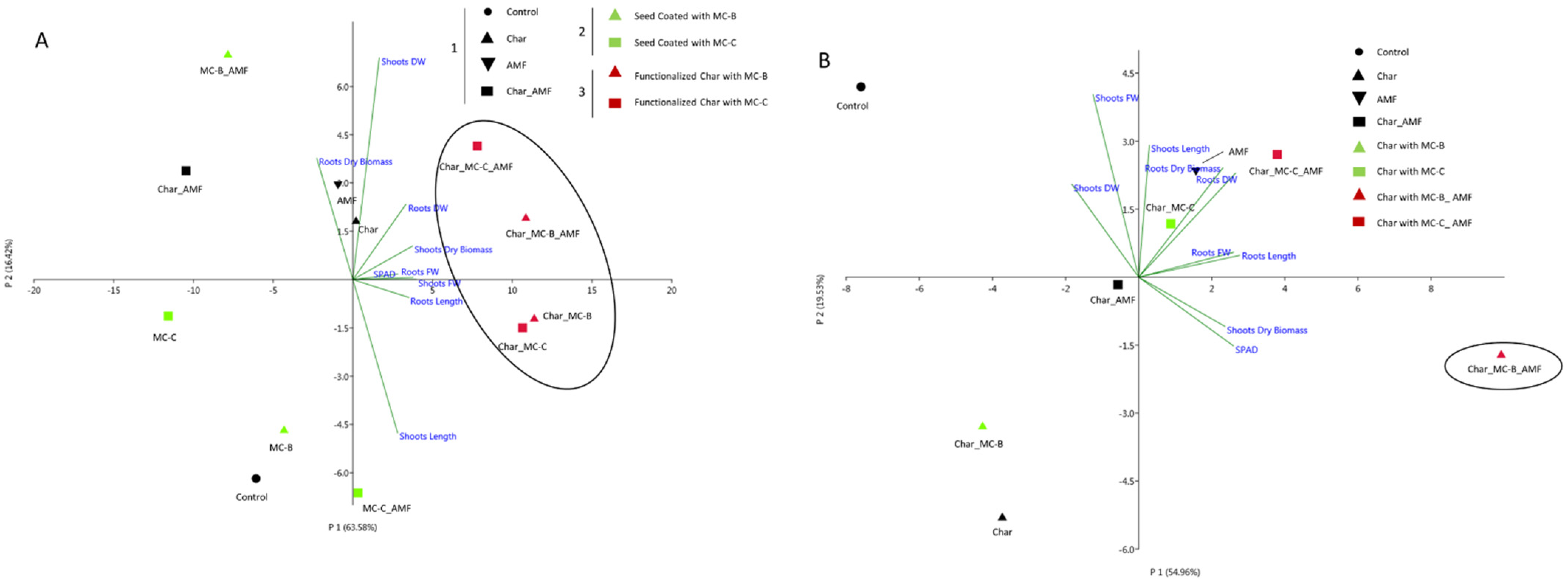
2.5. Correlation between Physiological and Molecular Data
3. Materials and Methods
3.1. Strains and Growth Conditions
3.2. Seed Coating and Liquid Delivery
3.3. Biochar Functionalization
3.4. Experimental Setup in Greenhouse
3.5. Plant Growth Parameters
3.6. Metabarcoding Analysis of Rhizosphere Soil
3.7. Target Genes Selection and Primers Design
3.8. Expression Analysis by RT-qtPCR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant Growth Promoting Rhizobacteria Dietzia Natronolimnaea Modulates the Expression of Stress Responsive Genes Providing Protection of Wheat from Salinity Stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef] [PubMed]
- Nath Yadav, A. Plant Growth Promoting Bacteria: Biodiversity and Multifunctional Attributes for Sustainable Agriculture. Adv. Biotechnol. Microbiol. 2017, 5, 555671. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- Valenzuela-Aragon, B.; Parra-Cota, F.I.; Santoyo, G.; Arellano-Wattenbarger, G.L.; de los Santos-Villalobos, S. Plant-Assisted Selection: A Promising Alternative for in Vivo Identification of Wheat (Triticum turgidum L. Subsp. Durum) Growth Promoting Bacteria. Plant Soil 2019, 435, 367–384. [Google Scholar] [CrossRef]
- Timsina, J. Can Organic Sources of Nutrients Increase Crop Yields to Meet Global Food Demand? Agronomy 2018, 8, 214. [Google Scholar] [CrossRef]
- Ramakrishnan, B.; Maddela, N.R.; Venkateswarlu, K.; Megharaj, M. Organic Farming: Does It Contribute to Contaminant-Free Produce and Ensure Food Safety? Sci. Total Environ. 2021, 769, 145079. [Google Scholar] [CrossRef]
- FAO. Inorganic Fertilizers 1961–2018; FAO: Rome, Italy, 2020. [Google Scholar]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A Critical Evaluation of Nanopesticides and Nanofertilizers against Their Conventional Analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef]
- Kah, M.; Tufenkji, N.; White, J.C. Nano-Enabled Strategies to Enhance Crop Nutrition and Protection. Nat. Nanotechnol. 2019, 14, 532–540. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture—Trends and Challenges. Annu. Rep. 2017, 296, 1–180. [Google Scholar]
- Authors NUR OKUR, A.; Hüsnü Kayikçioğlu, H.; Bülent Okur, I.; Yağmur, B.; Teresa Sponza, D.; Serdar Kara, R.; Agric For, T.J.; Okur, N.; Hüsnü KAYIKÇIOĞLU, H.; Okur, B.; et al. A Study of Olive Mill Wastewaters Obtained from Different Treatment Processes Effects Onchemical and Microbial Properties of a Typic Xerofluvent Soil and Wheat Yield. Turk. J. Agric. For. 2020, 44, 140–155. [Google Scholar] [CrossRef]
- dos Lopes, M.J.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful Plant Growth-Promoting Microbes: Inoculation Methods and Abiotic Factors. Front. Sustain. Food Syst. 2021, 5, 48. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Thalooth, A.T.; Elewa, T.A.; Ahmed, A.G. Yield and Nutrient Status of Wheat Plants (Triticum aestivum) as Affected by Sludge, Compost, and Biofertilizers under Newly Reclaimed Soil. Bull. Natl. Res. Cent. 2019, 43, 31. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 871, 1473. [Google Scholar] [CrossRef] [PubMed]
- Beesley, L.; Marmiroli, M. The Immobilisation and Retention of Soluble Arsenic, Cadmium and Zinc by Biochar. Environ. Pollut. 2011, 159, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Marmiroli, M.; Bonas, U.; Imperiale, D.; Lencioni, G.; Mussi, F.; Marmiroli, N.; Maestri, E. Structural and Functional Features of Chars from Different Biomasses as Potential Plant Amendments. Front. Plant Sci. 2018, 9, 1119. [Google Scholar] [CrossRef]
- Chen, B.; Yuan, M.; Qian, L. Enhanced Bioremediation of PAH-Contaminated Soil by Immobilized Bacteria with Plant Residue and Biochar as Carriers. J. Soils Sediments 2012, 12, 1350–1359. [Google Scholar] [CrossRef]
- Muter, O.; Grantina-ievina, L.; Makarenkova, G.; Vecstaudza, D.; Strikauska, S.; Selga, T.; Kasparinskis, R.; Stelmahere, S. Effect of Biochar and Trichoderma Application on Fungal Diversity and Growth of Zea Mays in a Sandy Loam Soil. Environ. Exp. Biol. 2017, 15, 289–296. [Google Scholar] [CrossRef]
- Ribera, J.; Gandía, M.; Marcos, J.F.; Bas, M.D.C.; Fink, S.; Schwarze, F.W.M.R. Effect of Trichoderma-Enriched Organic Charcoal in the Integrated Wood Protection Strategy. PLoS ONE 2017, 12, e0183004. [Google Scholar] [CrossRef]
- Semida, W.M.; Beheiry, H.R.; Sétamou, M.; Simpson, C.R.; Abd El-Mageed, T.A.; Rady, M.M.; Nelson, S.D. Biochar Implications for Sustainable Agriculture and Environment: A Review. S. Afr. J. Bot. 2019, 127, 333–347. [Google Scholar] [CrossRef]
- Rawat, J.; Sanwal, P.; Saxena, J. Towards the Mechanisms of Nutrient Solubilization and Fixation in Soil System. In Role of Rhizospheric Microbes in Soil; Meena, V., Ed.; Springer: Singapore, 2018; pp. 229–257. [Google Scholar]
- Fantin, V.; Righi, S.; Rondini, I.; Masoni, P. Environmental Assessment of Wheat and Maize Production in an Italian Farmers’ Cooperative. J. Clean. Prod. 2017, 140, 631–643. [Google Scholar] [CrossRef]
- Pospisil, A.; Pospisil, M. Agronomic Traits of Einkorn and Emmer under Different Seeding Agronomic Traits of Einkorn and Emmer under Different Seeding Rates and Topdressingwith Organic Fertilizers Rates and Topdressingwith Organic Fertilizers. Turk. J. Agric. For. 2020, 44, 95–102. [Google Scholar] [CrossRef]
- Pellegrino, E.; Piazza, G.; Arduini, I.; Ercoli, L. Field Inoculation of Bread Wheat with Rhizophagus Irregularis under Organic Farming: Variability in Growth Response and Nutritional Uptake of Eleven Old Genotypes and a Modern Variety. Agronomy 2020, 10, 333. [Google Scholar] [CrossRef]
- Ganugi, P.; Masoni, A.; Pietramellara, G.; Benedettelli, S. A Review of Studies from the Last Twenty Years on Plant–Arbuscular Mycorrhizal Fungi Associations and Their Uses for Wheat Crops. Agronomy 2019, 9, 840. [Google Scholar] [CrossRef]
- De Leon, D.G.; Vahter, T.; Zobel, M.; Koppel, M.; Edesi, L.; Davison, J.; Al-Quraishy, S.; Hozzein, W.N.; Moora, M.; Oja, J.; et al. Different Wheat Cultivars Exhibit Variable Responses to Inoculation with Arbuscular Mycorrhizal Fungi from Organic and Conventional Farms. PLoS ONE 2020, 15, e0233878. [Google Scholar] [CrossRef]
- Zhang, Y.; Lou, H.; Guo, D.; Zhang, R.; Su, M.; Hou, Z.; Zhou, H.; Liang, R.; Xie, C.; You, M.; et al. Identifying Changes in the Wheat Kernel Proteome under Heat Stress Using ITRAQ. Crop J. 2018, 6, 600–610. [Google Scholar] [CrossRef]
- Saboor, A.; Ali, M.A.; Danish, S.; Ahmed, N.; Fahad, S.; Datta, R.; Ansari, M.J.; Nasif, O.; Rahman, M.H.; Glick, B.R. Effect of Arbuscular Mycorrhizal Fungi on the Physiological Functioning of Maize under Zinc-Deficient Soils. Sci. Rep. 2021, 11, 18468. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Gill, A.R.; Jewell, N.; Brien, C.J.; Berger, B.; Tran, B.T.T.; Mace, E.; Cruickshank, A.W.; Jordan, D.R.; Garnett, T.; et al. Enhancement of Sorghum Grain Yield and Nutrition: A Role for Arbuscular Mycorrhizal Fungi Regardless of Soil Phosphorus Availability. Plants People Planet 2022, 4, 143–156. [Google Scholar] [CrossRef]
- Tran, B.T.T.; Cavagnaro, T.R.; Watts-Williams, S.J. Arbuscular Mycorrhizal Fungal Inoculation and Soil Zinc Fertilisation Affect the Productivity and the Bioavailability of Zinc and Iron in Durum Wheat. Mycorrhiza 2019, 29, 445–457. [Google Scholar] [CrossRef]
- Li, X.; Quan, X.; Mang, M.; Neumann, G.; Melchinger, A.; Ludewig, U. Flint Maize Root Mycorrhization and Organic Acid Exudates under Phosphorus Deficiency: Trends in Breeding Lines and Doubled Haploid Lines from Landraces. J. Plant Nutr. Soil Sci. 2021, 184, 346–359. [Google Scholar] [CrossRef]
- Hassan, T.U.; Bano, A. Biofertilizer: A Novel Formulation for Improving Wheat Growth, Physiology and Yield. Pakistan J. Bot. 2016, 48, 2233–2241. [Google Scholar]
- Dal Cortivo, C.; Ferrari, M.; Visioli, G.; Lauro, M.; Fornasier, F.; Barion, G.; Panozzo, A.; Vamerali, T. Effects of Seed-Applied Biofertilizers on Rhizosphere Biodiversity and Growth of Common Wheat (Triticum aestivum L.) in the Field. Front. Plant Sci. 2020, 11, 72. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.; Liu, S.; Li, Z.; Tan, X.; Huang, X.; Zeng, G.; Zhou, L.; Zheng, B. Biochar to Improve Soil Fertility. A Review. Agron. Sustain. Dev. 2016, 36, 36. [Google Scholar] [CrossRef]
- Schmidt, H.P.; Kammann, C.; Hagemann, N.; Leifeld, J.; Bucheli, T.D.; Sánchez Monedero, M.A.; Cayuela, M.L. Biochar in Agriculture—A Systematic Review of 26 Global Meta-Analyses. GCB Bioenergy 2021, 13, 1708–1730. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, S.; Verma, S.; Sar, T.; Sarsaiya, S.; Ravindran, B.; Liu, T.; Sindhu, R.; Patel, A.K.; Binod, P.; et al. Production and Beneficial Impact of Biochar for Environmental Application: A Comprehensive Review. Bioresour. Technol. 2021, 337, 125451. [Google Scholar] [CrossRef]
- Schröder, P.; Beckers, B.; Daniels, S.; Gnädinger, F.; Maestri, E.; Marmiroli, N.; Mench, M.; Millan, R.; Obermeier, M.M.; Oustriere, N.; et al. Intensify Production, Transform Biomass to Energy and Novel Goods and Protect Soils in Europe—A Vision How to Mobilize Marginal Lands. Sci. Total Environ. 2018, 616–617, 1101–1123. [Google Scholar] [CrossRef] [PubMed]
- Quilliam, R.S.; Glanville, H.C.; Wade, S.C.; Jones, D.L. Life in the “charosphere”—Does Biochar in Agricultural Soil Provide a Significant Habitat for Microorganisms? Soil Biol. Biochem. 2013, 65, 287–293. [Google Scholar] [CrossRef]
- Yrjälä, K.; Lopez-Echartea, E. Structure and Function of Biochar in Remediation and as Carrier of Microbes. Adv. Chem. Pollut. Environ. Manag. Prot. 2021, 7, 263–294. [Google Scholar]
- Tabacchioni, S.; Passato, S.; Ambrosino, P.; Huang, L.; Caldara, M.; Cantale, C.; Hett, J.; Del Fiore, A.; Fiore, A.; Schlüter, A.; et al. Identification of Beneficial Microbial Consortia and Bioactive Compounds with Potential as Plant Biostimulants for a Sustainable Agriculture. Microorganisms 2021, 9, 426. [Google Scholar] [CrossRef]
- Hett, J.; Neuhoff, D.; Döring, T.F.; Masoero, G.; Ercole, E.; Bevivino, A. Effects of Multi-Species Microbial Inoculants on Early Wheat Growth and Litterbag Microbial Activity. Agronomy 2022, 12, 899. [Google Scholar] [CrossRef]
- Malusá, E.; Sas-Paszt, L.; Ciesielska, J. Technologies for Beneficial Microorganisms Inocula Used as Biofertilizers. Sci. World J. 2012, 2012, 491206. [Google Scholar] [CrossRef]
- Rocha, I.; Ma, Y.; Souza-Alonso, P.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Seed Coating: A Tool for Delivering Beneficial Microbes to Agricultural Crops. Front. Plant Sci. 2019, 10, 1357. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Shurigin, V.; Jabborova, D.; Dela Cruz, J.A.; Dela Cruz, T.E.; Wirth, S.; Bellingrath-Kimura, S.D.; Egamberdieva, D. The Integrated Effect of Microbial Inoculants and Biochar Types on Soil Biological Properties, and Plant Growth of Lettuce (Lactuca sativa L.). Plants 2022, 11, 423. [Google Scholar] [CrossRef] [PubMed]
- Deaker, R.; Hartley, E.; Gemell, G. Conditions Affecting Shelf-Life of Inoculated Legume Seed. Agronomy 2012, 2, 38–51. [Google Scholar] [CrossRef]
- Tu, L.; He, Y.; Shan, C.; Wu, Z. Preparation of Microencapsulated Bacillus Subtilis SL-13 Seed Coating Agents and Their Effects on the Growth of Cotton Seedlings. Biomed Res. Int. 2016, 2016, 3251357. [Google Scholar] [CrossRef]
- Bevivino, A.; Sarrocco, S.; Dalmastri, C.; Tabacchioni, S.; Cantale, C.; Chiarini, L. Characterization of a Free-Living Maize-Rhizosphere Population of Burkholderia Cepacia: Effect of Seed Treatment on Disease Suppression and Growth Promotion of Maize. FEMS Microbiol. Ecol. 1998, 27, 225–237. [Google Scholar] [CrossRef][Green Version]
- Mulas, D.; García-Fraile, P.; Carro, L.; Ramírez-Bahena, M.H.; Casquero, P.; Velázquez, E.; González-Andrés, F. Distribution and Efficiency of Rhizobium Leguminosarum Strains Nodulating Phaseolus Vulgaris in Northern Spanish Soils: Selection of Native Strains That Replace Conventional N Fertilization. Soil Biol. Biochem. 2011, 43, 2283–2293. [Google Scholar] [CrossRef]
- Amirkhani, M.; Mayton, H.S.; Netravali, A.N.; Taylor, A.G. A Seed Coating Delivery System for Bio-Based Biostimulants to Enhance Plant Growth. Sustainability 2019, 11, 5304. [Google Scholar] [CrossRef]
- Latini, A.; Fiorani, F.; Galeffi, P.; Cantale, C.; Bevivino, A.; Jablonowski, N.D. Phenotyping of Different Italian Durum Wheat Varieties in Early Growth Stage with the Addition of Pure or Digestate-Activated Biochars. Front. Plant Sci. 2021, 12, 782072. [Google Scholar] [CrossRef]
- Glodowska, M. Biochar as a Potential Inoculant Carrier for Plant-Beneficial Bacteria. Master’s Thesis, McGill University, Sainte-Anne-de-Bellevue, QC, Canada, 2014. [Google Scholar]
- Sani, M.N.H.; Hasan, M.; Uddain, J.; Subramaniam, S. Impact of Application of Trichoderma and Biochar on Growth, Productivity and Nutritional Quality of Tomato under Reduced N-P-K Fertilization. Ann. Agric. Sci. 2020, 65, 107–115. [Google Scholar] [CrossRef]
- Nguyen, M.L. Biostimulant Effects of Rhizobacteria on Wheat Growth and Nutrient Uptake under Contrasted N Supplies. Ph.D. Thesis, Université de Liège, Liège, Belgium, 2018. [Google Scholar]
- Khabbaz, S.E.; Ladhalakshmi, D.; Babu, M.; Kandan, A.; Ramamoorthy, V.; Saravanakumar, D.; Al-Mughrabi, T.; Kandasamy, S. Plant Growth Promoting Bacteria (PGPB)—A Versatile Tool for Plant Health Management. Can. J. Pestic. Pest Manag. 2019, 1, 1. [Google Scholar] [CrossRef]
- Sandini, I.E.; Pacentchuk, F.; Hungria, M.; Nogueira, M.A.; da Cruz, S.P.; Nakatani, A.S.; Araújo, R.S. Seed Inoculation with Pseudomonas Fluorescens Promotes Growth, Yield and Reduces Nitrogen Application in Maize. Int. J. Agric. Biol. 2019, 22, 1369–1375. [Google Scholar] [CrossRef]
- Aasfar, A.; Bargaz, A.; Yaakoubi, K.; Hilali, A.; Bennis, I.; Zeroual, Y.; Meftah Kadmiri, I. Nitrogen Fixing Azotobacter Species as Potential Soil Biological Enhancers for Crop Nutrition and Yield Stability. Front. Microbiol. 2021, 12, 628379. [Google Scholar] [CrossRef] [PubMed]
- Mowafy, A.M.; Fawzy, M.M.; Gebreil, A.; Elsayed, A. Endophytic Bacillus, Enterobacter, and Klebsiella Enhance the Growth and Yield of Maize. Acta Agric. Scand. Sect. B Soil Plant Sci. 2021, 71, 237–246. [Google Scholar] [CrossRef]
- Ciccillo, F.; Fiore, A.; Bevivino, A.; Dalmastri, C.; Tabacchioni, S.; Chiarini, L. Effects of Two Different Application Methods of Burkholderia Ambifaria MCI 7 on Plant Growth and Rhizospheric Bacterial Diversity. Environ. Microbiol. 2002, 4, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Berg, G. Plant-Microbe Interactions Promoting Plant Growth and Health: Perspectives for Controlled Use of Microorganisms in Agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, E.; Roberts, I.N.; Montecchia, M.S.; Gutierrez-Boem, F.H.; Gomez, F.M.; Ruiz, J.A. A Novel Burkholderia Ambifaria Strain Able to Degrade the Mycotoxin Fusaric Acid and to Inhibit Fusarium Spp. Growth. Microbiol. Res. 2018, 206, 50–59. [Google Scholar] [CrossRef]
- Luo, C.; Xie, S.; Sun, W.; Li, X.; Cupples, A.M. Identification of a Novel Toluene-Degrading Bacterium from the Candidate Phylum TM7, as Determined by DNA Stable Isotope Probing. Appl. Environ. Microbiol. 2009, 75, 4644–4647. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Blagodatsky, S.; Selivanovskaya, S. Response of Bacterial and Fungal Communities to High Petroleum Pollution in Different Soils. Sci. Rep. 2021, 11, 164. [Google Scholar] [CrossRef]
- Wang, X.; Song, D.; Liang, G.; Zhang, Q.; Ai, C.; Zhou, W. Maize Biochar Addition Rate Influences Soil Enzyme Activity and Microbial Community Composition in a Fluvo-Aquic Soil. Appl. Soil Ecol. 2015, 96, 265–272. [Google Scholar] [CrossRef]
- Latini, A.; Bacci, G.; Teodoro, M.; Gattia, D.M.; Bevivino, A.; Trakal, L. The Impact of Soil-Applied Biochars From Different Vegetal Feedstocks on Durum Wheat Plant Performance and Rhizospheric Bacterial Microbiota in Low Metal-Contaminated Soil. Front. Microbiol. 2019, 10, 2694. [Google Scholar] [CrossRef]
- Brtnicky, M.; Datta, R.; Holatko, J.; Bielska, L.; Gusiatin, Z.M.; Kucerik, J.; Hammerschmiedt, T.; Danish, S.; Radziemska, M.; Mravcova, L.; et al. A Critical Review of the Possible Adverse Effects of Biochar in the Soil Environment. Sci. Total Environ. 2021, 796, 148756. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhu, M.; Wang, J.; Liu, X.; Guo, W.; Zheng, J.; Bian, R.; Wang, G.; Zhang, X.; Cheng, K.; et al. The Responses of Soil Organic Carbon Mineralization and Microbial Communities to Fresh and Aged Biochar Soil Amendments. GCB Bioenergy 2019, 11, 1408–1420. [Google Scholar] [CrossRef]
- White, R.A.; Bottos, E.M.; Roy Chowdhury, T.; Zucker, J.D.; Brislawn, C.J.; Nicora, C.D.; Fansler, S.J.; Glaesemann, K.R.; Glass, K.; Jansson, J.K. Moleculo Long-Read Sequencing Facilitates Assembly and Genomic Binning from Complex Soil Metagenomes. mSystems 2016, 1, e00045-16. [Google Scholar] [CrossRef]
- Silva, V.P.; Moreira-Santos, M.; Mateus, C.; Teixeira, T.; Ribeiro, R.; Viegas, C.A. Evaluation of Arthrobacter Aurescens Strain Tc1 as Bioaugmentation Bacterium in Soils Contaminated with the Herbicidal Substance Terbuthylazine. PLoS ONE 2015, 10, e0144978. [Google Scholar] [CrossRef]
- Singh, S.; Singh, N.; Kumar, V.; Datta, S.; Wani, A.B.; Singh, D.; Singh, K.; Singh, J. Toxicity, Monitoring and Biodegradation of the Fungicide Carbendazim. Environ. Chem. Lett. 2016, 14, 317–329. [Google Scholar] [CrossRef]
- Alvarez, A.; Saez, J.M.; Davila Costa, J.S.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current Research and Perspectives for Bioremediation of Pesticides and Heavy Metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef]
- Mawang, C.I.; Azman, A.S.; Fuad, A.S.M.; Ahamad, M. Actinobacteria: An Eco-Friendly and Promising Technology for the Bioaugmentation of Contaminants. Biotechnol. Rep. 2021, 32, e00679. [Google Scholar] [CrossRef]
- Bonfante, P.; Venice, F. Mucoromycota: Going to the Roots of Plant-Interacting Fungi. Fungal Biol. Rev. 2020, 34, 100–113. [Google Scholar] [CrossRef]
- Bell, N.L.; Adam, K.H.; Jones, R.J.; Johnson, R.D.; Mtandavari, Y.F.; Burch, G.; Cave, V.; Cameron, C.; Maclean, P.; Popay, A.J.; et al. Detection of Invertebrate Suppressive Soils, and Identification of a Possible Biological Control Agent for Meloidogyne Nematodes Using High Resolution Rhizosphere Microbial Community Analysis. Front. Plant Sci. 2016, 7, 1946. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Adachi, Y.; Sugiyama, S. Soil Productivity and Structure of Bacterial and Fungal Communities in Unfertilized Arable Soil. PLoS ONE 2018, 13, e0204085. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, T.; Zhao, J.; Wang, L.; Yang, D.; Li, G.; Xiu, W. Variation of Soil Bacterial and Fungal Communities from Fluvo-Aquic Soil Under Chemical Fertilizer Reduction Combined with Organic Materials in North China Plain. J. Soil Sci. Plant Nutr. 2021, 21, 349–363. [Google Scholar] [CrossRef]
- Bello, A.; Wang, B.; Zhao, Y.; Yang, W.; Ogundeji, A.; Deng, L.; Egbeagu, U.U.; Yu, S.; Zhao, L.; Li, D.; et al. Composted Biochar Affects Structural Dynamics, Function and Co-Occurrence Network Patterns of Fungi Community. Sci. Total Environ. 2021, 775, 145672. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, N.; Schmitz, J.; Polatajko, A.; Schlüter, U.; Fahnenstich, H.; Witt, S.; Fernie, A.R.; Uroic, K.; Scholz, U.; Sonnewald, U.; et al. An Integrated Functional Approach to Dissect Systemic Responses in Maize to Arbuscular Mycorrhizal Symbiosis. Plant Cell Environ. 2015, 38, 1591–1612. [Google Scholar] [CrossRef] [PubMed]
- Alharby, H.F.; Fahad, S. Melatonin Application Enhances Biochar Efficiency for Drought Tolerance in Maize Varieties: Modifications in Physio-Biochemical Machinery. Agron. J. 2020, 112, 2826–2847. [Google Scholar] [CrossRef]
- Santoyo, G.; Gamalero, E.; Glick, B.R. Mycorrhizal-Bacterial Amelioration of Plant Abiotic and Biotic Stress. Front. Sustain. Food Syst. 2021, 5, 672881. [Google Scholar] [CrossRef]
- Vannini, C.; Domingo, G.; Fiorilli, V.; Seco, D.G.; Novero, M.; Marsoni, M.; Wisniewski-Dye, F.; Bracale, M.; Moulin, L.; Bonfante, P. Proteomic Analysis Reveals How Pairing of a Mycorrhizal Fungus with Plant Growth-Promoting Bacteria Modulates Growth and Defense in Wheat. Plant Cell Environ. 2021, 44, 1946–1960. [Google Scholar] [CrossRef]
- Fracasso, A.; Telò, L.; Lanfranco, L.; Bonfante, P.; Amaducci, S. Physiological Beneficial Effect of Rhizophagus Intraradices Inoculation on Tomato Plant Yield under Water Deficit Conditions. Agronomy 2020, 10, 71. [Google Scholar] [CrossRef]
- Lehmann, J.; Cowie, A.; Masiello, C.A.; Kammann, C.; Woolf, D.; Amonette, J.E.; Cayuela, M.L.; Camps-Arbestain, M.; Whitman, T. Biochar in Climate Change Mitigation. Nat. Geosci. 2021, 14, 883–892. [Google Scholar] [CrossRef]
- Caldara, M.; Marmiroli, N. Tricyclic Antidepressants Inhibit Candida Albicans Growth and Biofilm Formation. Int. J. Antimicrob. Agents 2018, 52, 500–505. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a Prokaryotic Universal Primer for Simultaneous Analysis of Bacteria and Archaea Using Next-Generation Sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Basciani, S.; Camajani, E.; Contini, S.; Persichetti, A.; Risi, R.; Bertoldi, L.; Strigari, L.; Prossomariti, G.; Watanabe, M.; Mariani, S.; et al. Very-Low-Calorie Ketogenic Diets with Whey, Vegetable, or Animal Protein in Patients with Obesity: A Randomized Pilot Study. J. Clin. Endocrinol. Metab. 2020, 105, 2939–2949. [Google Scholar] [CrossRef] [PubMed]
- Juhmani, A.S.; Vezzi, A.; Wahsha, M.; Buosi, A.; De Pascale, F.; Schiavon, R.; Sfriso, A. Diversity and Dynamics of Seaweed Associated Microbial Communities Inhabiting the Lagoon of Venice. Microorganisms 2020, 8, 1657. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Monga, D.; Shetti, N.P.; Basu, S.; Raghava Reddy, K.; Badawi, M.; Bonilla-Petriciolet, A.; Aminabhavi, T.M. Engineered Biochar: A Way Forward to Environmental Remediation. Fuel 2022, 311, 122510. [Google Scholar] [CrossRef]
- Abadi, V.A.J.M.; Sepehri, M.; Rahmani, H.A.; Zarei, M.; Ronaghi, A.; Taghavi, S.M.; Shamshiripour, M. Correction to: Role of Dominant Phyllosphere Bacteria with Plant Growth–Promoting Characteristics on Growth and Nutrition of Maize (Zea mays L.). J. Soil Sci. Plant Nutr. 2021, 21, 2740. [Google Scholar] [CrossRef]
- El-Fattah, D.A.A.; Eweda, W.; Zayed, M.; Hassanein, M. Effect of carrier materials, sterilization method, and storage temperature on survival and biological activities of Azotobacter chroococcum inoculant. Ann. Agric. Sci. 2013, 58, 111–118. [Google Scholar] [CrossRef]
- Boari, A.; Zuccari, D.; Vurro, M. ‘Microbigation’: Delivery of biological control agents through drip irrigation systems. Irrig. Sci. 2007, 26, 101–107. [Google Scholar] [CrossRef]
- Daza, A.; Santamaría, C.; Rodríguez-Navarro, D.; Camacho, M.; Orive, R.; Temprano, F. Perlite as a carrier for bacterial inoculants. Soil Biol. Biochem. 2000, 32, 567–572. [Google Scholar] [CrossRef]
- Duo, L.A.; Liu, C.X.; Zhao, S.L. Alleviation of Drought Stress in Turfgrass by the Combined Application of Nano-compost and Microbes from Compost. Russ. J. Plant Physiol. 2018, 65, 419–426. [Google Scholar] [CrossRef]
- Esitken, A.; Pirlak, L.; Turan, M.; Sahin, F. Effects of floral and foliar application of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrition of sweet cherry. Sci. Hortic. 2006, 110, 324–327. [Google Scholar] [CrossRef]
- Jambhulkar, P.P.; Sharma, P.; Yadav, R. Delivery Systems for Introduction of Microbial Inoculants in the Field. In Microbial Inoculants in Sustainable Agricultural Productivity: Vol. 2: Functional Applications; Springer: Berlin, Germany, 2016; pp. 199–218. [Google Scholar] [CrossRef]
- Ma, Y. Seed coating with beneficial microorganisms for precision agriculture. Biotechnol. Adv. 2019, 37, 107423. [Google Scholar] [CrossRef] [PubMed]
- Madden, T.; Grentzer, J.M.; Secura, G.M.; Allsworth, J.; Peipert, J.F. Risk of Bacterial Vaginosis in Users of the Intrauterine Device. Sex Transm Dis. 2012, 39, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Nuti, M.; Giovannetti, G. Borderline Products between Bio-fertilizers/ Bio-effectors and Plant Protectants: The Role of Microbial Consortia. J. Agric. Sci. Technol. A 2015, 5, 305–315. [Google Scholar] [CrossRef]
- Nehra, V.; Choudhary, M. A review on plant growth promoting rhizobacteria acting as bioinoculants and their biological approach towards the production of sustainable agriculture. J. Appl. Nat. Sci. 2015, 7, 540–556. [Google Scholar] [CrossRef]
- Preininger, C.; Sauer, U.; Bejarano, A.; Berninger, T. Concepts and applications of foliar spray for microbial inoculants. Appl. Microbiol. Biotechnol. 2018, 102, 7265–7282. [Google Scholar] [CrossRef]
- Singh, D.P.; Singh, H.B.; Prabha, R. Microbial Inoculants in Sustainable Agricultural Productivity: Vol. 2: Functional Applications; Springer: Berlin, Germany, 2016. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
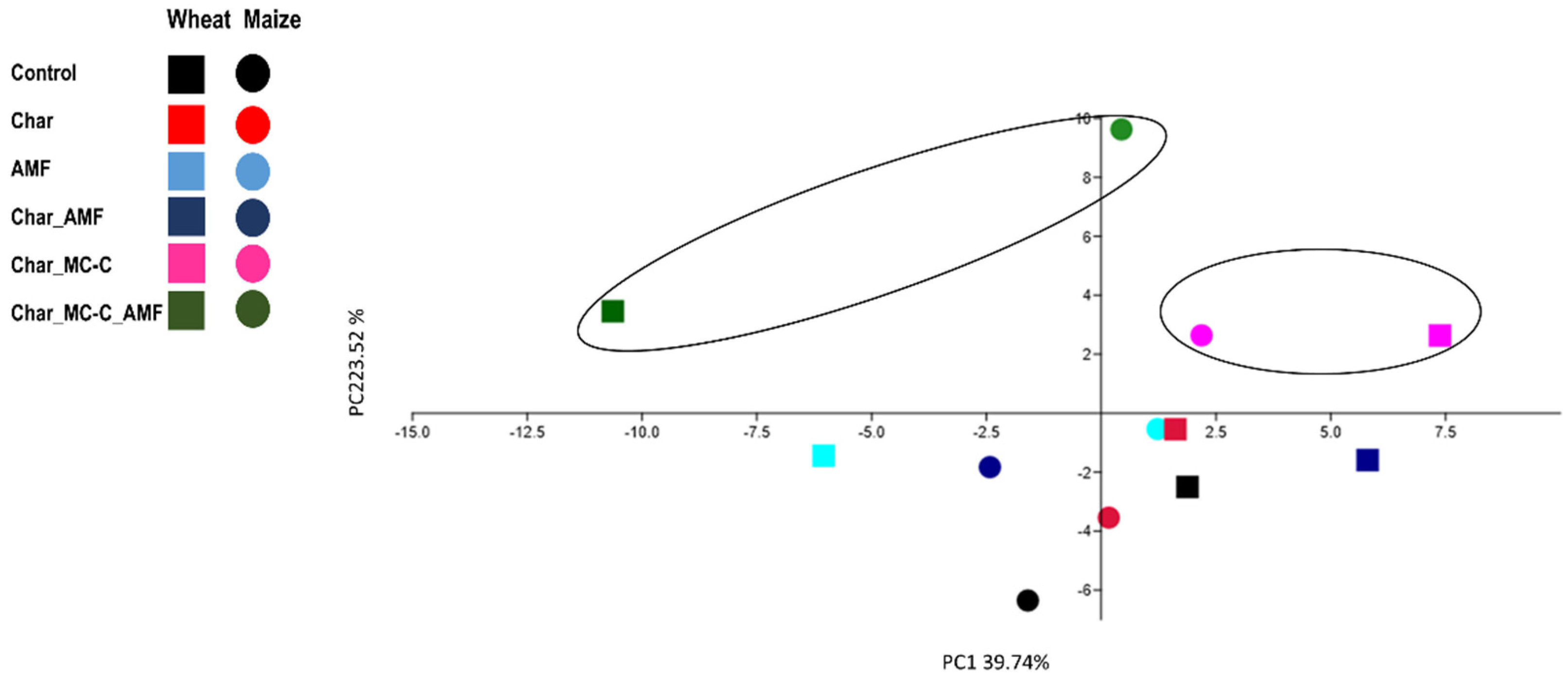{kind=link}
| Group | Length (cm) | Fresh Weight (FW) (g) | Dry Weight (DW) (g) | Dry Biomass % | Chlorophyll Content (SPAD) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments | Roots | Shoots | Roots | Shoots | Roots | Shoots | Roots | Shoots | Leaves |
| 1_Control | |||||||||
| Control | 61.2 ± 6.7 b | 33.3 ± 2.6 b | 0.7 ± 0.1 b | 0.4 ± 0.1 b | 0.1 ± 0.1 b | 0.12 ± 0.02 b | 11.4± 3.4 bc | 28.1 ± 2.7 ab | 25.8 ± 2.2 abc |
| Char | 26.2 ± 2.3 c | 32.0 ± 5.1 b | 0.7 ± 0.3 b | 0.6 ± 0.1 b | 0.1 ± 0.0 b | 0.16 ± 0.03 b | 15.5± 6.1 bc | 28.1 ± 6.2 ab | 27.2 ± 2.3 abc |
| AMF | 64.6 ± 9.0 ab | 31.8 ± 4.3 b | 0.7 ± 0.1 b | 0.5 ± 0.1 b | 0.1 ± 0.0 b | 0.15 ± 0.04 b | 15.7± 3.2 bc | 26.7 ± 13.3 ab | 27.7 ± 1.6 ab |
| Char_AMF | 44.4 ± 3.4 c | 30.0 ± 3.7 b | 0.5 ± 0.1 b | 0.4 ± 0.1 b | 0.1 ± 0.0 b | 0.31 ± 0.03 b | 14.6± 3.2 bc | 37.9 ± 10.8 a | 24.1 ± 3.5 bc |
| 2_Seed Coating | |||||||||
| MC-B | 53.6 ± 24.8 b | 35.4 ± 4.7 b | 0.7 ± 0.2 b | 0.5 ± 0.1 b | 0.1 ± 0.0 b | 0.15 ± 0.01 b | 12.5± 1.0 bc | 27.2 ± 4.3 ab | 23.6 ± 1.6 c |
| MC-C | 35.6 ± 17.2 bc | 27.8 ± 1.3 b | 0.6 ± 0.1 b | 0.4 ± 0.1 b | 0.1 ± 0.0 b | 0.14 ± 0.01 b | 11.4± 5.1 bc | 30.8 ± 7.6 ab | 24.2 ± 2.1 bc |
| MC-B_ AMF | 52.6 ± 8.1 ab | 28.4 ± 5.7 b | 0.4 ± 0.2 b | 0.4 ± 0.1 b | 0.1 ± 0.0 b | 0.15 ± 0.01 b | 18.5± 6.8 ab | 31.4 ± 4.6 ab | 22.2 ± 5.2 bc |
| MC-C_AMF | 39.4 ± 3.3 c | 31.6 ± 5.3 b | 1.5 ± 0.5 a | 0.6 ± 0.1 b | 0.1 ± 0.1 b | 0.16 ± 0.04 b | 9.9± 10.6 c | 27.2 ± 8.2 ab | 26.2 ± 3.3 abc |
| 3_Functionalized Char | |||||||||
| Char_MC-B | 49.2 ± 9.4 b | 45.0 ± 7.3 a | 1.6 ± 0.5 a | 1.9 ± 0.1 a | 0.2 ± 0.1 a | 0.47 ± 0.07 a | 15.2± 7.8 bc | 24.1 ± 2.2 b | 29.0 ± 4.1 a |
| Char_MC-C | 48.6 ± 10.0 b | 45.6 ± 3.3 a | 2.0 ± 0.7 a | 1.4 ± 0.4 a | 0.3 ± 0.1 a | 0.42 ± 0.02 a | 14.7± 3.7 bc | 30.6 ± 8.4 b | 33.3 ± 8.7 a |
| Char_MC-B_AMF | 54.8 ± 3.3 b | 45.4 ± 11.5 a | 1.6 ± 0.3 a | 1.7 ± 0.1 a | 0.3 ± 0.1 a | 0.44 ± 0.02 a | 17.3± 3.0 ab | 28.4 ± 3.0 b | 29.5 ± 1.5 a |
| Char_MC-C_AMF | 74.8 ± 7.0 a | 48.2 ± 3.3 a | 0.6 ± 0.1 b | 1.5 ± 0.3 a | 0.2 ± 0.0 b | 0.22 ± 0.06 a | 26.4 ± 6.7 a | 16.0 ± 6.1 c | 27.8 ± 3.0 ab |
| Treatment | Length (cm) | Fresh Weight (FW) (g) | Dry Weight (DW) (g) | Dry Biomass % | Chlorophyll Content (SPAD) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Roots | Shoots | Roots | Shoots | Roots | Shoots | Roots | Shoots | Leaves | |
| Control | 51.7 ± 10.3 b | 56.0 ± 8.0 c | 3.1 ± 0.7 | 4.3 ± 1.7 c | 0.3 ± 0.1 | 0.6 ± 0.2 c | 10.3 ± 1.2 | 14.5 ± 1.4 ab | 9.4 ± 2.4 d |
| Char | 59.2 ± 16.6 b | 68.0 ± 10.9 a | 2.6 ± 0.5 | 5.0 ± 3.3 c | 0.3 ± 0.6 | 0.6 ± 0.3 c | 9.0 ± 1.0 | 12.0 ± 0.9 c | 13.0 ± 2.6 c |
| AMF | 70.8 ± 9.3 a | 62.8 ± 13.8 b | 3.1 ± 1.1 | 6.6 ± 2.9 abc | 0.2 ± 0.1 | 1.1 ± 0.7 abc | 8.7 ± 3.0 | 15.4 ± 2.7 ab | 12.2 ± 1.8 c |
| Char_AMF | 61.2 ± 10.5 b | 68.3 ± 20.0 a | 3.2 ± 1.7 | 5.3 ± 2.4 bc | 0.2 ± 0.1 | 0.7 ± 0.3 bc | 8.4 ± 1.9 | 13.0 ± 1.3 c | 13.4 ± 3.4 bc |
| Char_MC-B | 61.7 ± 13.0 b | 64.5 ± 14.5 a | 2.6 ± 0.4 | 4.8 ± 2.0 c | 0.2 ± 0.1 | 0.6 ± 0.3 c | 9.2 ± 0.7 | 14.3 ± 0.1 abc | 12.5 ± 2.3 c |
| Char_MC-B_AMF | 88.2 ± 21.5 a | 62.2 ± 9.6 b | 3.0 ± 0.9 | 12.5 ±6.4 a | 0.2 ± 0.1 | 2.7 ± 0.1 a | 8.1 ± 1.2 | 22.3 ± 8.7 a | 17.5 ± 3.8 a |
| Char_MC-C | 65.2 ± 7.0 b | 65.2 ± 9.1 abc | 3.0 ± 0.4 | 4.7 ± 1.8 c | 0.3 ± 0.1 | 0.7 ± 0.3 c | 9.1 ± 1.3 | 14.6 ± 2.2 ab | 15.0 ± 3.7 ab |
| Char_MC-C_AMF | 81.0 ± 10.3 a | 59.7 ± 11.8 b | 2.7 ± 1.0 | 8.9 ± 3.1 b | 0.3 ± 0.1 | 1.5 ± 0.6 a | 9.5 ± 1.1 | 16.1 ± 1.9 ab | 14.5 ± 3.4 ab |
| WHEAT | |||||||||||||||
| Physiological Parameters * | Metagenomic § | Gene Expression # | |||||||||||||
| Length (cm) | FW (g) | DW (g) | Chlorophyll Content (SPAD) | Shannon Index | 21d | 60d | |||||||||
| Treatments | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Bacteria | Fungi | Up | Up | Total | Mean | |
| Control | 10 | 7 | 8 | 4 | 8 | 1 | 3 | 7 | 5 | 2 | 4 | 1 | 1 | 61 | 4.7 |
| Char | 1 | 6 | 8 | 8 | 8 | 7 | 8 | 7 | 7 | 1 | 1 | 7 | 7 | 76 | 5.8 |
| AMF | 11 | 5 | 8 | 6 | 8 | 5 | 9 | 3 | 8 | 6 | 4 | 3 | 2 | 78 | 6 |
| Char_AMF | 4 | 3 | 2 | 4 | 8 | 9 | 5 | 12 | 3 | 6 | 2 | 5 | 4 | 67 | 5.2 |
| MC-B | 8 | 8 | 8 | 6 | 8 | 5 | 4 | 5 | 2 | n.t. | n.t. | n.t. | n.t. | 54 | 6 |
| MC-C | 2 | 1 | 4 | 4 | 8 | 2 | 3 | 10 | 4 | 6 | 7 | 7 | 5 | 63 | 4.8 |
| MC-B_AMF | 7 | 2 | 1 | 4 | 8 | 5 | 11 | 11 | 1 | n.t. | n.t. | n.t. | n.t. | 50 | 5.6 |
| MC-C_AMF | 3 | 5 | 9 | 8 | 8 | 7 | 1 | 5 | 6 | 6 | 7 | 8 | 8 | 81 | 6.2 |
| Char_MC-B | 6 | 9 | 11 | 12 | 10 | 12 | 7 | 2 | 10 | n.t. | n.t. | n.t. | n.t. | 79 | 8.8 |
| Char_MC-C | 5 | 11 | 12 | 9 | 12 | 10 | 6 | 9 | 12 | 8 | 5 | 5 | 7 | 111 | 8.5 |
| Char_MC-B_AMF | 9 | 10 | 11 | 11 | 12 | 11 | 10 | 8 | 11 | n.t. | n.t. | n.t. | n.t. | 93 | 10.3 |
| Char_MC-C_AMF | 12 | 12 | 4 | 10 | 10 | 8 | 12 | 1 | 9 | 7 | 8 | 3 | 4 | 100 | 7.7 |
| MAIZE | |||||||||||||||
| Physiological Parameters * | Metagenomic § | Gene Expression # | |||||||||||||
| Length (cm) | FW (g) | DW (g) | Chlorophyll Content (SPAD) | Shannon Index | 21d | 60d | |||||||||
| Treatments | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Bacteria | Fungi | Up | Up | Total | Mean | |
| Control | 1 | 1 | 7 | 1 | 8 | 3 | 8 | 4 | 1 | 5 | 8 | 1 | 2 | 50 | 3.8 |
| Char | 2 | 7 | 2 | 4 | 8 | 3 | 4 | 1 | 4 | 2 | 7 | 7 | 8 | 59 | 4.5 |
| AMF | 6 | 4 | 7 | 6 | 4 | 6 | 3 | 6 | 2 | 7 | 5 | 6 | 7 | 69 | 5.3 |
| Char_AMF | 3 | 8 | 8 | 5 | 4 | 5 | 2 | 2 | 5 | 2 | 6 | 6 | 2 | 58 | 4.5 |
| Char_MC-B | 4 | 5 | 2 | 3 | 4 | 3 | 6 | 3 | 3 | 6 | 4 | 6 | 5 | 54 | 4.2 |
| Char_MC-C | 5 | 6 | 5 | 2 | 8 | 5 | 5 | 5 | 7 | 8 | 2 | 8 | 4 | 70 | 5.4 |
| Char_MC-B_AMF | 8 | 3 | 5 | 8 | 4 | 8 | 1 | 8 | 8 | 5 | 3 | 2 | 4 | 67 | 5.2 |
| Char_MC-C_AMF | 7 | 2 | 3 | 7 | 8 | 7 | 7 | 7 | 6 | 5 | 1 | 6 | 7 | 73 | 5.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graziano, S.; Caldara, M.; Gullì, M.; Bevivino, A.; Maestri, E.; Marmiroli, N. A Metagenomic and Gene Expression Analysis in Wheat (T. durum) and Maize (Z. mays) Biofertilized with PGPM and Biochar. Int. J. Mol. Sci. 2022, 23, 10376. https://doi.org/10.3390/ijms231810376
Graziano S, Caldara M, Gullì M, Bevivino A, Maestri E, Marmiroli N. A Metagenomic and Gene Expression Analysis in Wheat (T. durum) and Maize (Z. mays) Biofertilized with PGPM and Biochar. International Journal of Molecular Sciences. 2022; 23(18):10376. https://doi.org/10.3390/ijms231810376
Chicago/Turabian StyleGraziano, Sara, Marina Caldara, Mariolina Gullì, Annamaria Bevivino, Elena Maestri, and Nelson Marmiroli. 2022. "A Metagenomic and Gene Expression Analysis in Wheat (T. durum) and Maize (Z. mays) Biofertilized with PGPM and Biochar" International Journal of Molecular Sciences 23, no. 18: 10376. https://doi.org/10.3390/ijms231810376
APA StyleGraziano, S., Caldara, M., Gullì, M., Bevivino, A., Maestri, E., & Marmiroli, N. (2022). A Metagenomic and Gene Expression Analysis in Wheat (T. durum) and Maize (Z. mays) Biofertilized with PGPM and Biochar. International Journal of Molecular Sciences, 23(18), 10376. https://doi.org/10.3390/ijms231810376