

Sympathetic Nerve Activity and Blood Pressure Response to Exercise in Peripheral Artery Disease: From Molecular Mechanisms, Human Studies, to Intervention Strategy Development

{kind=link}
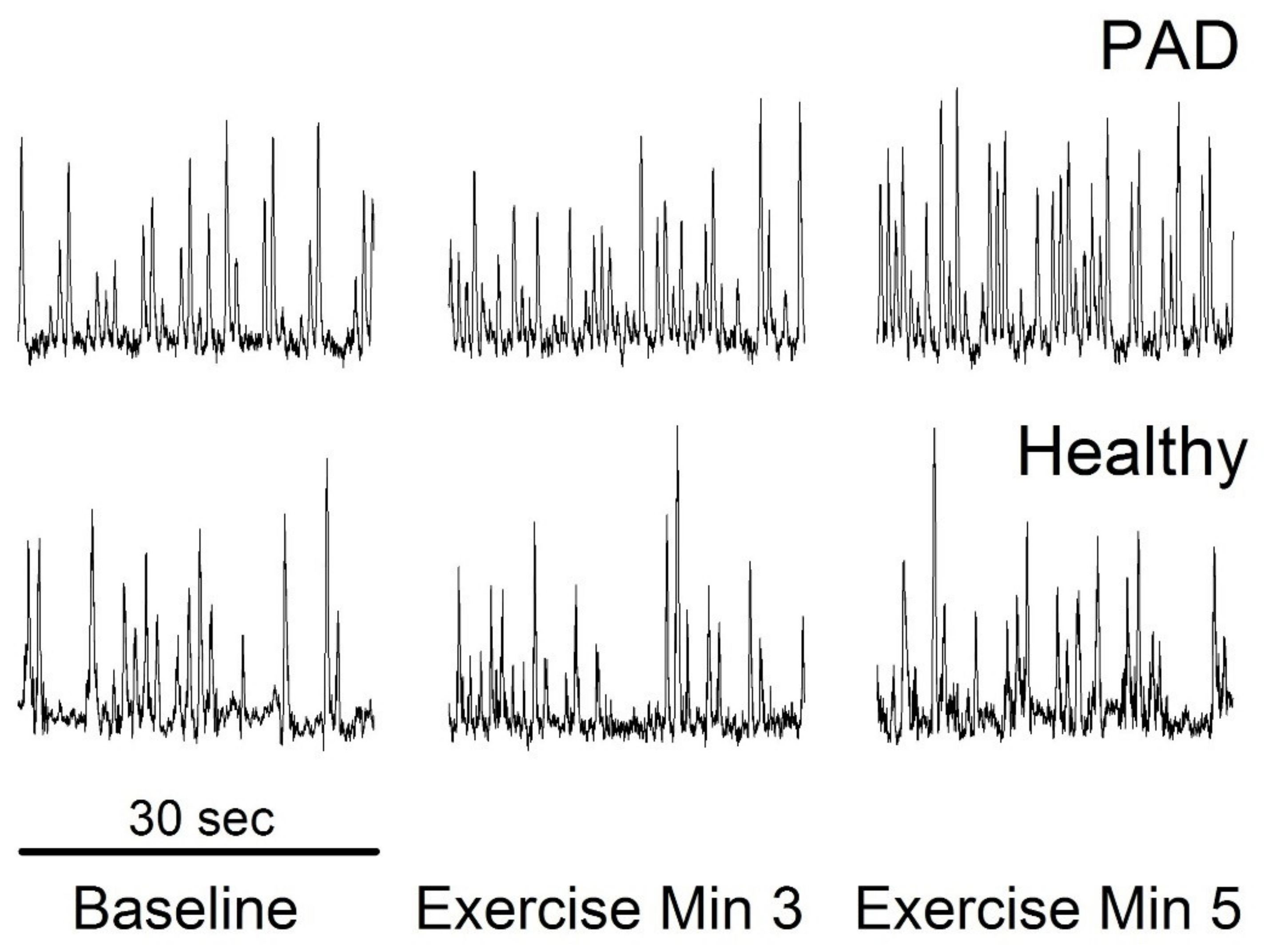{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Sympathetic Nerves and BP Regulation during Exercise in PAD
2.1. Exercise Pressor Reflex (EPR)
2.2. EPR in PAD Patients
3. Experimental Models to Study the Pathological Status in Human PAD
3.1. Blood Flow Restriction (BFR) and Ischemia-Reperfusion (IR) in Healthy Subject
3.2. Animal Models of Studying Human PAD
3.2.1. Femoral Artery Occlusion/Ligation
3.2.2. Hindlimb Ischemia-Reperfusion
4. Molecular Mechanisms Leading to Exaggerated SNA and BP Responses in PAD
4.1. Effects of Muscle Metabolic Products and Their Responsive Receptors (Figure 5 and Figure 6)


4.1.1. ASIC3 KO Suppresses the Exercise Pressor Response under Ischemic Situation
4.1.2. ASIC3 KO Attenuates the Exercise Pressor Response and the Activities of P2X3 under Ischemic Situation
4.2. Other Ischemia-Induced Products
4.2.1. HIF-1α
4.2.2. Reactive Oxidative Species
4.2.3. Endothelin-1 (ET-1)
4.3. Pro-Inflammation Cytokines and Ion Channels in Muscle Sensory Neurons (Figure 7 and Figure 8)
4.3.1. TNF-α and Activities of Nav Channels in Muscle DRG Neurons


4.3.2. IL-6 and Activities of Kv4 Channels in Muscle DRG Neurons
5. Heat Treatment and Nutrition Intervention on Improving Exercise-Induced Exaggerated SNA and BP Responses in PAD
5.1. Heat Treatment (Figure 7 and Figure 8)
5.2. Effects of Supplemental Nutrients
5.2.1. Vitamin B6
5.2.2. Vitamin C
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Criqui, M.H.; Aboyans, V. Epidemiology of peripheral artery disease. Circ. Res. 2015, 116, 1509–1526. [Google Scholar] [CrossRef] [PubMed]
- Fowkes, F.G.; Aboyans, V.; Fowkes, F.J.; McDermott, M.M.; Sampson, U.K.; Criqui, M.H. Peripheral artery disease: Epidemiology and global perspectives. Nat. Rev. Cardiol. 2017, 14, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Criqui, M.H.; Langer, R.D.; Fronek, A.; Feigelson, H.S.; Klauber, M.R.; McCann, T.J.; Browner, D. Mortality over a period of 10 years in patients with peripheral arterial disease. N. Engl. J. Med. 1992, 326, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; de Ferranti, S.; Despres, J.P.; Fullerton, H.J.; et al. Executive Summary: Heart Disease and Stroke Statistics--2016 Update: A Report From the American Heart Association. Circulation 2016, 133, 447–454. [Google Scholar] [CrossRef]
- Ouriel, K. Peripheral arterial disease. Lancet 2001, 358, 1257–1264. [Google Scholar] [CrossRef]
- Anand, S.S.; Caron, F.; Eikelboom, J.W.; Bosch, J.; Dyal, L.; Aboyans, V.; Abola, M.T.; Branch, K.R.H.; Keltai, K.; Bhatt, D.L.; et al. Major Adverse Limb Events and Mortality in Patients With Peripheral Artery Disease: The COMPASS Trial. J. Am. Coll. Cardiol. 2018, 71, 2306–2315. [Google Scholar] [CrossRef]
- Bauersachs, R.; Zeymer, U.; Brière, J.B.; Marre, C.; Bowrin, K.; Huelsebeck, M. Burden of Coronary Artery Disease and Peripheral Artery Disease: A Literature Review. Cardiovasc. Ther. 2019, 2019, 8295054. [Google Scholar] [CrossRef]
- Hirsch, A.T.; Haskal, Z.J.; Hertzer, N.R.; Bakal, C.W.; Creager, M.A.; Halperin, J.L.; Hiratzka, L.F.; Murphy, W.R.; Olin, J.W.; Puschett, J.B.; et al. ACC/AHA Guidelines for the Management of Patients with Peripheral Arterial Disease (lower extremity, renal, mesenteric, and abdominal aortic): A collaborative report from the American Associations for Vascular Surgery/Society for Vascular Surgery, Society for Cardiovascular Angiography and Interventions, Society for Vascular Medicine and Biology, Society of Interventional Radiology, and the ACC/AHA Task Force on Practice Guidelines (writing committee to develop guidelines for the management of patients with peripheral arterial disease)—Summary of recommendations. J. Vasc. Interv. Radiol. 2006, 17, 1383–1397, quiz 1398. [Google Scholar] [CrossRef]
- Clagett, G.P.; Sobel, M.; Jackson, M.R.; Lip, G.Y.H.; Tangelder, M.; Verhaeghe, R. Antithrombotic Therapy in Peripheral Arterial Occlusive Disease. The Seventh ACCP Conference on Antithrombotic and Thrombolytic Therapy. Chest 2004, 126, 609S–626S. [Google Scholar] [CrossRef]
- Kitrou, P.; Katsanos, K.; Karnabatidis, D.; Reppas, L.; Brountzos, E.; Spiliopoulos, S. Current Evidence and Future Perspectives on Anti-platelet and Statin Pharmacotherapy for Patients with Symptomatic Peripheral Arterial Disease. Curr. Vasc. Pharmacol. 2017, 15, 430–445. [Google Scholar] [CrossRef]
- Brendle, D.C.; Joseph, L.J.; Corretti, M.C.; Gardner, A.W.; Katzel, L.I. Effects of exercise rehabilitation on endothelial reactivity in older patients with peripheral arterial disease. Am. J. Cardiol. 2001, 87, 324–329. [Google Scholar] [CrossRef]
- Hamburg, N.M.; Balady, G.J. Exercise rehabilitation in peripheral artery disease: Functional impact and mechanisms of benefits. Circulation 2011, 123, 87–97. [Google Scholar] [CrossRef]
- McDermott, M.M. Exercise Rehabilitation for Peripheral Artery Disease: A REVIEW. J. Cardiopulm. Rehabil. Prev. 2018, 38, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Baccelli, G.; Reggiani, P.; Mattioli, A.; Corbellini, E.; Garducci, S.; Catalano, M. The exercise pressor reflex and changes in radial arterial pressure and heart rate during walking in patients with arteriosclerosis obliterans. Angiology 1999, 50, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Bakke, E.F.; Hisdal, J.; Jorgensen, J.J.; Kroese, A.; Stranden, E. Blood pressure in patients with intermittent claudication increases continuously during walking. Eur. J. Vasc. Endovasc. Surg. 2007, 33, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Lorentsen, E. Systemic arterial blood pressure during exercise in patients with atherosclerosis obliterans of the lower limbs. Circulation 1972, 46, 257–263. [Google Scholar] [CrossRef]
- Muller, M.D.; Drew, R.C.; Blaha, C.A.; Mast, J.L.; Cui, J.; Reed, A.B.; Sinoway, L.I. Oxidative stress contributes to the augmented exercise pressor reflex in peripheral arterial disease patients. J. Physiol. 2012, 590, 6237–6246. [Google Scholar] [CrossRef]
- Lewis, G.D.; Gona, P.; Larson, M.G.; Plehn, J.F.; Benjamin, E.J.; O’Donnell, C.J.; Levy, D.; Vasan, R.S.; Wang, T.J. Exercise blood pressure and the risk of incident cardiovascular disease (from the Framingham Heart Study). Am. J. Cardiol. 2008, 101, 1614–1620. [Google Scholar] [CrossRef]
- Thanassoulis, G.; Lyass, A.; Benjamin, E.J.; Larson, M.G.; Vita, J.A.; Levy, D.; Hamburg, N.M.; Widlansky, M.E.; O’Donnell, C.J.; Mitchell, G.F.; et al. Relations of exercise blood pressure response to cardiovascular risk factors and vascular function in the Framingham Heart Study. Circulation 2012, 125, 2836–2843. [Google Scholar] [CrossRef]
- Sinoway, L.; Prophet, S.; Gorman, I.; Mosher, T.; Shenberger, J.; Dolecki, M.; Briggs, R.; Zelis, R. Muscle Acidosis during Static Exercise Is Associated with Calf Vasoconstriction. J. Appl. Physiol. 1989, 66, 429–436. [Google Scholar] [CrossRef]
- Victor, R.G.; Bertocci, L.; Pryor, S.; Nunnally, R. Sympathetic nerve discharge is coupled to muscle cell pH during exercise in humans. J. Clin. Investig. 1988, 82, 1301–1305. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.M.; McCloskey, D.I.; Mitchell, J.H. Cardiovascular and respiratory responses to changes in central command during isometric exercise at constant muscle tension. J. Physiol. 1972, 226, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, T.G.; Mullins, D.C.; Millhorn, D.E. Control of respiration by the hypothalamus and by feedback from contracting muscles in cats. Respir. Physiol. 1986, 64, 317–328. [Google Scholar] [CrossRef]
- Victor, R.G.; Mark, A.L. Interaction of cardiopulmonary and carotid baroreflex control of vascular resistance in humans. J. Clin. Investig. 1985, 76, 1592–1598. [Google Scholar] [CrossRef]
- Freyschuss, U. Cardiovascular adjustment to somatomotor activation. The elicitation of increments in heart rate, aortic pressure and venomotor tone with the initiation of muscle contraction. Acta Physiol. Scand. 1970, 342, 1–63. [Google Scholar]
- Coote, J.H.; Hilton, S.M.; Pérez-González, J.F. The reflex nature of the pressor response to muscular exercise. J. Physiol. 1971, 215, 789–804. [Google Scholar] [CrossRef]
- Waldrop, T.G.; Eldridge, F.L.; Iwamoto, G.A.; Mitchell, J.H. Central neural control of respiration and circulation during exercise. Chapter 9. In Handbook of Physiology—Section 12, Exercise: Regulation and Integration of Multiple Systems; Rowell, L.B., Shepherd, J.T., Eds.; Oxford University Press: New York, NY, USA, 1996; pp. 333–380. [Google Scholar]
- Mitchell, J.H.; Kaufman, M.P.; Iwamoto, G.A. The exercise pressor reflex: Its cardiovascular effects, afferent mechanisms, and central pathways. Annu. Rev. Physiol. 1983, 45, 229–242. [Google Scholar] [CrossRef]
- Kaufman, M.P.; Forster, H.V. Reflexes controlling circulatory, ventilatory and airway responses to exercise. Chapter 10. In Handbook of Physiology—Section 12, Exercise: Regulation and Integration of Multiple Systems; Rowell, L.B., Shepherd, J.T., Eds.; Oxford University Press: New York, NY, USA, 1996; pp. 381–447. [Google Scholar]
- Drew, R.C.; Muller, M.D.; Blaha, C.A.; Mast, J.L.; Heffernan, M.J.; Estep, L.E.; Cui, J.; Reed, A.B.; Sinoway, L.I. Renal vasoconstriction is augmented during exercise in patients with peripheral arterial disease. Physiol. Rep. 2013, 1, e00154. [Google Scholar] [CrossRef]
- Kim, D.J.; Kuroki, M.; Cui, J.; Gao, Z.; Luck, J.C.; Pai, S.; Miller, A.; Sinoway, L. Systemic and regional hemodynamic response to activation of the exercise pressor reflex in patients with peripheral artery disease. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H916–H924. [Google Scholar] [CrossRef]
- Ritti-Dias, R.M.; Meneses, A.L.; Parker, D.E.; Montgomery, P.S.; Khurana, A.; Gardner, A.W. Cardiovascular responses to walking in patients with peripheral artery disease. Med. Sci. Sports Exerc. 2011, 43, 2017–2023. [Google Scholar] [CrossRef]
- Cui, J.; Leuenberger, U.; Kim, D.; Luck, J.; Pai, S.; Blaha, C.; Cauffman, A.; Sinoway, L. Muscle Sympathetic Nerve Activity Responses to Exercise in Patients with Peripheral Artery Disease. FASEB J. 2021, 35, 35. [Google Scholar] [CrossRef]
- Garcia, L.A. Epidemiology and pathophysiology of lower extremity peripheral arterial disease. J. Endovasc. Ther. 2006, 13, II3–II9. [Google Scholar] [CrossRef] [PubMed]
- Hiatt, W.R.; Brass, E.P. Pathophysiology of Peripheral Artery Disease, Intermittent Claudication, and Critical Limb Ischemia. Chapter 17. In Vascular Medicine: A Companion to Braunwald’s Heart Disease; Creager, M., Beckman, J.A., Loscalzo, J., Eds.; Saunders: Philadelphia, PA, USA, 2013; pp. 223–230. [Google Scholar]
- Hirsch, A.T.; Criqui, M.H.; Treat-Jacobson, D.; Regensteiner, J.G.; Creager, M.A.; Olin, J.W.; Krook, S.H.; Hunninghake, D.B.; Comerota, A.J.; Walsh, M.E.; et al. Peripheral arterial disease detection, awareness, and treatment in primary care. JAMA 2001, 286, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Isbell, D.C.; Berr, S.S.; Toledano, A.Y.; Epstein, F.H.; Meyer, C.H.; Rogers, W.J.; Harthun, N.L.; Hagspiel, K.D.; Weltman, A.; Kramer, C.M. Delayed calf muscle phosphocreatine recovery after exercise identifies peripheral arterial disease. J. Am. Coll. Cardiol. 2006, 47, 2289–2295. [Google Scholar] [CrossRef]
- Anderson, J.D.; Epstein, F.H.; Meyer, C.H.; Hagspiel, K.D.; Wang, H.; Berr, S.S.; Harthun, N.L.; Weltman, A.; Dimaria, J.M.; West, A.M.; et al. Multifactorial determinants of functional capacity in peripheral arterial disease: Uncoupling of calf muscle perfusion and metabolism. J. Am. Coll. Cardiol. 2009, 54, 628–635. [Google Scholar] [CrossRef]
- Sheriff, D.; Wyss, C.; Rowell, L.; Scher, A. Does inadequate oxygen delivery trigger pressor response to muscle hypoperfusion during exercise? Am. J. Physiol. Heart Circ. Physiol. 1987, 253, H1199–H1207. [Google Scholar] [CrossRef]
- O’Leary, D.S. Point: The muscle metaboreflex does restore blood flow to contracting muscles. J. Appl. Physiol. 2006, 100, 357–358, discussion 360–361. [Google Scholar] [CrossRef]
- Crisafulli, A.; de Farias, R.R.; Farinatti, P.; Lopes, K.G.; Milia, R.; Sainas, G.; Pinna, V.; Palazzolo, G.; Doneddu, A.; Magnani, S.; et al. Blood Flow Restriction Training Reduces Blood Pressure During Exercise Without Affecting Metaboreflex Activity. Front. Physiol. 2018, 9, 1736. [Google Scholar] [CrossRef]
- Jessee, M.B.; Buckner, S.L.; Mouser, J.G.; Mattocks, K.T.; Dankel, S.J.; Abe, T.; Bell, Z.W.; Bentley, J.P.; Loenneke, J.P. Muscle Adaptations to High-Load Training and Very Low-Load Training With and Without Blood Flow Restriction. Front. Physiol. 2018, 9, 1448. [Google Scholar] [CrossRef]
- Neto, G.R.; Novaes, J.S.; Dias, I.; Brown, A.; Vianna, J.; Cirilo-Sousa, M.S. Effects of resistance training with blood flow restriction on haemodynamics: A systematic review. Clin. Physiol. Funct. Imaging 2017, 37, 567–574. [Google Scholar] [CrossRef]
- Patterson, S.D.; Hughes, L.; Warmington, S.; Burr, J.; Scott, B.R.; Owens, J.; Abe, T.; Nielsen, J.L.; Libardi, C.A.; Laurentino, G.; et al. Blood Flow Restriction Exercise: Considerations of Methodology, Application, and Safety. Front. Physiol. 2019, 10, 533. [Google Scholar] [CrossRef] [PubMed]
- Libardi, C.A.; Chacon-Mikahil, M.P.; Cavaglieri, C.R.; Tricoli, V.; Roschel, H.; Vechin, F.C.; Conceicao, M.S.; Ugrinowitsch, C. Effect of concurrent training with blood flow restriction in the elderly. Int. J. Sports Med. 2015, 36, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Stavres, J.; Singer, T.J.; Brochetti, A.; Kilbane, M.J.; Brose, S.W.; McDaniel, J. The Feasibility of Blood Flow Restriction Exercise in Patients With Incomplete Spinal Cord Injury. PM R 2018, 10, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Singer, T.J.; Stavres, J.; Elmer, S.J.; Kilgas, M.A.; Pollock, B.S.; Kearney, S.G.; McDaniel, J. Knee extension with blood flow restriction: Impact of cuff pressure on hemodynamics. Eur. J. Appl. Physiol. 2020, 120, 79–90. [Google Scholar] [CrossRef]
- Cryer, H.G. Therapeutic approaches for clinical ischemia and reperfusion injury. Shock 1997, 8, 26–32. [Google Scholar] [CrossRef]
- Maxwell, S.R.; Lip, G.Y. Reperfusion injury: A review of the pathophysiology, clinical manifestations and therapeutic options. Int. J. Cardiol. 1997, 58, 95–117. [Google Scholar] [CrossRef]
- Haimovici, H. Arterial embolism with acute massive ischemic myopathy and myoglobinuria: Evaluation of a hitherto unreported syndrome with report of two cases. Surgery 1960, 47, 739–747. [Google Scholar]
- Lau, C.S.; Scott, N.; Shaw, J.W.; Belch, J.J. Increased activity of oxygen free radicals during reperfusion in patients with peripheral arterial disease undergoing percutaneous peripheral artery balloon angioplasty. Int. Angiol. 1991, 10, 244–246. [Google Scholar]
- Edwards, A.T.; Blann, A.D.; Suarez-Mendez, V.J.; Lardi, A.M.; McCollum, C.N. Systemic responses in patients with intermittent claudication after treadmill exercise. Br. J. Surg. 1994, 81, 1738–1741. [Google Scholar] [CrossRef]
- Loukogeorgakis, S.P.; van den Berg, M.J.; Sofat, R.; Nitsch, D.; Charakida, M.; Haiyee, B.; de Groot, E.; MacAllister, R.J.; Kuijpers, T.W.; Deanfield, J.E. Role of NADPH oxidase in endothelial ischemia/reperfusion injury in humans. Circulation 2010, 121, 2310–2316. [Google Scholar] [CrossRef]
- Seeger, J.P.; Lenting, C.J.; Schreuder, T.H.; Landman, T.R.; Cable, N.T.; Hopman, M.T.; Thijssen, D.H. Interval exercise, but not endurance exercise, prevents endothelial ischemia-reperfusion injury in healthy subjects. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H351–H357. [Google Scholar] [CrossRef] [PubMed]
- Loukogeorgakis, S.P.; Panagiotidou, A.T.; Broadhead, M.W.; Donald, A.; Deanfield, J.E.; MacAllister, R.J. Remote ischemic preconditioning provides early and late protection against endothelial ischemia-reperfusion injury in humans: Role of the autonomic nervous system. J. Am. Coll. Cardiol. 2005, 46, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Brunt, V.E.; Jeckell, A.T.; Ely, B.R.; Howard, M.J.; Thijssen, D.H.; Minson, C.T. Acute hot water immersion is protective against impaired vascular function following forearm ischemia-reperfusion in young healthy humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R1060–R1067. [Google Scholar] [CrossRef] [PubMed]
- Ely, B.R.; Francisco, M.A.; Halliwill, J.R.; Bryan, S.D.; Comrada, L.N.; Larson, E.A.; Brunt, V.E.; Minson, C.T. Heat therapy reduces sympathetic activity and improves cardiovascular risk profile in women who are obese with polycystic ovary syndrome. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R630–R640. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; McQuillan, P.; Momen, A.; Blaha, C.; Moradkhan, R.; Mascarenhas, V.; Hogeman, C.S.; Krishnan, A.; Sinoway, L.I. The role of the cyclooxygenase products in evoking sympathetic activation in exercise. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1861–H1868. [Google Scholar] [CrossRef]
- Qin, L.; Li, J. Sympathetic Nerve Control of Blood Pressure Response during Exercise in Peripheral Artery Disease and Current Application of Experimental Disease Models. Am. J. Biomed Sci. Res. 2020, 9, 204–209. [Google Scholar] [CrossRef]
- Limbourg, A.; Korff, T.; Napp, L.C.; Schaper, W.; Drexler, H.; Limbourg, F.P. Evaluation of postnatal arteriogenesis and angiogenesis in a mouse model of hind-limb ischemia. Nat. Protoc. 2009, 4, 1737–1746. [Google Scholar] [CrossRef]
- Yu, J.; de Muinck, E.D.; Zhuang, Z.; Drinane, M.; Kauser, K.; Rubanyi, G.M.; Qian, H.S.; Murata, T.; Escalante, B.; Sessa, W.C. Endothelial nitric oxide synthase is critical for ischemic remodeling, mural cell recruitment, and blood flow reserve. Proc. Natl. Acad. Sci. USA 2005, 102, 10999–11004. [Google Scholar] [CrossRef]
- Westvik, T.S.; Fitzgerald, T.N.; Muto, A.; Maloney, S.P.; Pimiento, J.M.; Fancher, T.T.; Magri, D.; Westvik, H.H.; Nishibe, T.; Velazquez, O.C.; et al. Limb ischemia after iliac ligation in aged mice stimulates angiogenesis without arteriogenesis. J. Vasc. Surg. 2009, 49, 464–473. [Google Scholar] [CrossRef]
- Yu, J.; Dardik, A. A Murine Model of Hind Limb Ischemia to Study Angiogenesis and Arteriogenesis. Methods Mol. Biol. 2018, 1717, 135–143. [Google Scholar] [CrossRef]
- Schuler, D.; Sansone, R.; Nicolaus, C.; Kelm, M.; Heiss, C. Repetitive remote occlusion (RRO) stimulates eNOS-dependent blood flow and collateral expansion in hindlimb ischemia. Free. Radic. Biol. Med. 2018, 129, 520–531. [Google Scholar] [CrossRef]
- Goggi, J.L.; Haslop, A.; Boominathan, R.; Chan, K.; Soh, V.; Cheng, P.; Robins, E.G.; Bhakoo, K.K. Imaging the Proangiogenic Effects of Cardiovascular Drugs in a Diabetic Model of Limb Ischemia. Contrast Media Mol. Imaging 2019, 2019, 2538909. [Google Scholar] [CrossRef] [PubMed]
- Liddell, R.P.; Patel, T.H.; Weiss, C.R.; Lee, D.S.; Matsuhashi, T.; Brown, P.R.; Gabrielson, K.L.; Rodriguez, E.R.; Eng, J.; Kimura, H.; et al. Endovascular model of rabbit hindlimb ischemia: A platform to evaluate therapeutic angiogenesis. J. Vasc. Interv. Radiol. JVIR 2005, 16, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Long, C.A.; Timmins, L.H.; Koutakis, P.; Goodchild, T.T.; Lefer, D.J.; Pipinos, I.I.; Casale, G.P.; Brewster, L.P. An endovascular model of ischemic myopathy from peripheral arterial disease. J. Vasc. Surg. 2017, 66, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.S.; Kim, H.W.; Roh, D.H.; Yoon, S.Y.; Kwon, Y.B.; Han, H.J.; Chung, J.M.; Beitz, A.J.; Lee, J.H. A new rat model for thrombus-induced ischemic pain (TIIP); development of bilateral mechanical allodynia. Pain 2008, 139, 520–532. [Google Scholar] [CrossRef]
- Waters, R.E.; Terjung, R.L.; Peters, K.G.; Annex, B.H. Preclinical models of human peripheral arterial occlusive disease: Implications for investigation of therapeutic agents. J. Appl. Physiol. 2004, 97, 773–780. [Google Scholar] [CrossRef]
- Li, J.; Xing, J. Muscle afferent receptors engaged in augmented sympathetic responsiveness in peripheral artery disease. Front. Physiol. 2012, 3, 247. [Google Scholar] [CrossRef]
- Tsuchimochi, H.; McCord, J.L.; Hayes, S.G.; Koba, S.; Kaufman, M.P. Chronic femoral artery occlusion augments exercise pressor reflex in decerebrated rats. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H106–H113. [Google Scholar] [CrossRef]
- Kuczmarski, J.M.; Unrath, K.; Thomas, G.D. Exaggerated cardiovascular responses to treadmill running in rats with peripheral arterial insufficiency. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H114–H121. [Google Scholar] [CrossRef]
- Signorelli, S.S.; Katsiki, N. Oxidative Stress and Inflammation: Their Role in the Pathogenesis of Peripheral Artery Disease with or Without Type 2 Diabetes Mellitus. Curr. Vasc. Pharmacol. 2018, 16, 547–554. [Google Scholar] [CrossRef]
- Harms, J.E.; Kuczmarski, J.M.; Kim, J.S.; Thomas, G.D.; Kaufman, M.P. The role played by oxidative stress in evoking the exercise pressor reflex in health and simulated peripheral artery disease. J. Physiol. 2017, 595, 4365–4378. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Lu, J.; Liu, J.; Li, J. Local Injections of Superoxide Dismutase Attenuate the Exercise Pressor Reflex in Rats with Femoral Artery Occlusion. Front. Physiol. 2018, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Lu, J.; Li, J. Role of TNF-α in Regulating the Exercise Pressor Reflex in Rats With Femoral Artery Occlusion. Front. Physiol. 2018, 9, 1461. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.L.; Queme, L.F.; Shank, A.T.; Hudgins, R.C.; Jankowski, M.P. Sensitization of group III and IV muscle afferents in the mouse after ischemia and reperfusion injury. J. Pain 2014, 15, 1257–1270. [Google Scholar] [CrossRef]
- Qin, L.; Li, Q.; Li, J. Exaggerated Activities of P2X3 and ASIC3 Signaling Pathway in Muscle Afferent Following Himblimb Muscle Ischemia-Reperfusion. FASEB J. 2022, 36, 1257–1270. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.D.; Lu, J.; Xing, J.; Li, J. Contribution of nerve growth factor to upregulation of P2X(3) expression in DRG neurons of rats with femoral artery occlusion. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1070–H1079. [Google Scholar] [CrossRef]
- Xing, J.; Lu, J.; Li, J. TRPA1 mediates amplified sympathetic responsiveness to activation of metabolically sensitive muscle afferents in rats with femoral artery occlusion. Front. Physiol. 2015, 6, 249. [Google Scholar] [CrossRef]
- Liu, J.; Gao, Z.; Li, J. Femoral artery occlusion increases expression of ASIC3 in dorsal root ganglion neurons. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1357–H1364. [Google Scholar] [CrossRef]
- Xing, J.; Lu, J.; Li, J. Acid-sensing ion channel subtype 3 function and immunolabelling increases in skeletal muscle sensory neurons following femoral artery occlusion. J. Physiol. 2012, 590, 1261–1272. [Google Scholar] [CrossRef]
- Xing, J.; Lu, J.; Li, J. Augmented P2X response and immunolabeling in dorsal root ganglion neurons innervating skeletal muscle following femoral artery occlusion. J. Neurophysiol. 2013, 109, 2161–2168. [Google Scholar] [CrossRef]
- Leal, A.K.; Yamauchi, K.; Kim, J.; Ruiz-Velasco, V.; Kaufman, M.P. Peripheral delta-opioid receptors attenuate the exercise pressor reflex. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1246–H1255. [Google Scholar] [CrossRef] [PubMed]
- Leal, A.K.; McCord, J.L.; Tsuchimochi, H.; Kaufman, M.P. Blockade of the TP receptor attenuates the exercise pressor reflex in decerebrated rats with chronic femoral artery occlusion. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2140–H2146. [Google Scholar] [CrossRef] [PubMed]
- Tsuchimochi, H.; McCord, J.L.; Kaufman, M.P. Peripheral mu-opioid receptors attenuate the augmented exercise pressor reflex in rats with chronic femoral artery occlusion. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H557–H565. [Google Scholar] [CrossRef]
- Yamauchi, K.; Kim, J.S.; Stone, A.J.; Ruiz-Velasco, V.; Kaufman, M.P. Endoperoxide 4 receptors play a role in evoking the exercise pressor reflex in rats with simulated peripheral artery disease. J. Physiol. 2013, 591, 2949–2962. [Google Scholar] [CrossRef]
- Puntambekar, P.; Van Buren, J.; Raisinghani, M.; Premkumar, L.S.; Ramkumar, V. Direct interaction of adenosine with the TRPV1 channel protein. J. Neurosci. 2004, 24, 3663–3671. [Google Scholar] [CrossRef] [PubMed]
- Tsuzuki, K.; Ase, A.; Seguela, P.; Nakatsuka, T.; Wang, C.Y.; She, J.X.; Gu, J.G. TNPATP-resistant P2X ionic current on the central terminals and somata of rat primary sensory neurons. J. Neurophysiol. 2003, 89, 3235–3242. [Google Scholar] [CrossRef] [PubMed]
- Waldmann, R.; Bassilana, F.; de Weille, J.; Champigny, G.; Heurteaux, C.; Lazdunski, M. Molecular cloning of a non-inactivating proton-gated Na+ channel specific for sensory neurons. J. Biol. Chem. 1997, 272, 20975–20978. [Google Scholar] [CrossRef]
- Waldmann, R.; Champigny, G.; Bassilana, F.; Heurteaux, C.; Lazdunski, M. A proton-gated cation channel involved in acid-sensing. Nature 1997, 386, 173–177. [Google Scholar] [CrossRef]
- Waldmann, R.; Champigny, G.; Lingueglia, E.; De Weille, J.R.; Heurteaux, C.; Lazdunski, M. H(+)-gated cation channels. Ann. N. Y. Acad. Sci. 1999, 868, 67–76. [Google Scholar] [CrossRef]
- Light, A.R.; Hughen, R.W.; Zhang, J.; Rainier, J.; Liu, Z.; Lee, J. Dorsal root ganglion neurons innervating skeletal muscle respond to physiological combinations of protons, ATP, and lactate mediated by ASIC, P2X, and TRPV1. J. Neurophysiol. 2008, 100, 1184–1201. [Google Scholar] [CrossRef]
- Deval, E.; Noël, J.; Gasull, X.; Delaunay, A.; Alloui, A.; Friend, V.; Eschalier, A.; Lazdunski, M.; Lingueglia, E. Acid-sensing ion channels in postoperative pain. J. Neurosci. 2011, 31, 6059–6066. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Noël, J.; Lay, N.; Alloui, A.; Diochot, S.; Friend, V.; Jodar, M.; Lazdunski, M.; Lingueglia, E. ASIC3, a sensor of acidic and primary inflammatory pain. EMBO J. 2008, 19, 3047–3055. [Google Scholar] [CrossRef] [PubMed]
- MacLean, D.A.; LaNoue, K.F.; Gray, K.S.; Sinoway, L.I. Effects of hindlimb contraction on pressor and muscle interstitial metabolite responses in the cat. J. Appl. Physiol. 1998, 85, 1583–1592. [Google Scholar] [CrossRef]
- Rotto, D.M.; Stebbins, C.L.; Kaufman, M.P. Reflex cardiovascular and ventilatory responses to increasing H+ activity in cat hindlimb muscle. J. Appl. Physiol. 1989, 67, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Yagi, J.; Wenk, H.N.; Naves, L.A.; McCleskey, E.W. Sustained currents through ASIC3 ion channels at the modest pH changes that occur during myocardial ischemia. Circ. Res. 2006, 99, 501–509. [Google Scholar] [CrossRef]
- MacLean, D.A.; Imadojemu, V.A.; Sinoway, L.I. Interstitial pH, K+, lactate and phosphate determined with MSNA during exercise in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R563–R571. [Google Scholar] [CrossRef] [PubMed]
- Tsuchimochi, H.; Yamauchi, K.; McCord, J.L.; Kaufman, M.P. Blockade of acid sensing ion channels attenuates the augmented exercise pressor reflex in rats with chronic femoral artery occlusion (Abstract). FASEB J. 2011, 589, 6173–6189. [Google Scholar]
- Stephan, G.; Huang, L.; Tang, Y.; Vilotti, S.; Fabbretti, E.; Yu, Y.; Nörenberg, W.; Franke, H.; Gölöncsér, F.; Sperlágh, B.; et al. The ASIC3/P2X3 cognate receptor is a pain-relevant and ligand-gated cationic channel. Nat. Commun. 2018, 9, 1354. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Li, J.D.; Sinoway, L.I.; Li, J. Effect of muscle interstitial pH on P2X and TRPV1 receptor-mediated pressor response. J. Appl. Physiol. 2007, 102, 2288–2293. [Google Scholar] [CrossRef]
- Lu, J.; Xing, J.; Li, J. Bradykinin B2 receptor contributes to the exaggerated muscle mechanoreflex in rats with femoral artery occlusion. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1166–H1174. [Google Scholar] [CrossRef]
- Qin, L.; Li, J. Nerve growth factor in muscle afferent neurons of peripheral artery disease and autonomic function. Neural. Regen. Res. 2021, 16, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Jiang, B.H.; Rue, E.A.; Semenza, G.L. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. USA 1995, 92, 5510–5514. [Google Scholar] [CrossRef] [PubMed]
- Ceradini, D.J.; Kulkarni, A.R.; Callaghan, M.J.; Tepper, O.M.; Bastidas, N.; Kleinman, M.E.; Capla, J.M.; Galiano, R.D.; Levine, J.P.; Gurtner, G.C. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nat. Med. 2004, 10, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Iyer, N.V.; Kotch, L.E.; Agani, F.; Leung, S.W.; Laughner, E.; Wenger, R.H.; Gassmann, M.; Gearhart, J.D.; Lawler, A.M.; Yu, A.Y.; et al. Cellular and developmental control of O2 homeostasis by hypoxia-inducible factor 1 alpha. Genes. Dev. 1998, 12, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Kelly, B.D.; Hackett, S.F.; Hirota, K.; Oshima, Y.; Cai, Z.; Berg-Dixon, S.; Rowan, A.; Yan, Z.; Campochiaro, P.A.; Semenza, G.L. Cell type-specific regulation of angiogenic growth factor gene expression and induction of angiogenesis in nonischemic tissue by a constitutively active form of hypoxia-inducible factor 1. Circ. Res. 2003, 93, 1074–1081. [Google Scholar] [CrossRef]
- Manalo, D.J.; Rowan, A.; Lavoie, T.; Natarajan, L.; Kelly, B.D.; Ye, S.Q.; Garcia, J.G.; Semenza, G.L. Transcriptional regulation of vascular endothelial cell responses to hypoxia by HIF-1. Blood 2005, 105, 659–669. [Google Scholar] [CrossRef]
- Gao, W.; Li, J. Femoral artery occlusion increases muscle pressor reflex and expression of hypoxia-inducible factor-1α in sensory neurons. J. Cardiovasc. Dis. 2013, 1, 34–40. [Google Scholar]
- Jaakkola, P.; Mole, D.R.; Tian, Y.M.; Wilson, M.I.; Gielbert, J.; Gaskell, S.J.; Kriegsheim, A.; Hebestreit, H.F.; Mukherji, M.; Schofield, C.J.; et al. Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 2001, 292, 468–472. [Google Scholar] [CrossRef]
- Milkiewicz, M.; Pugh, C.W.; Egginton, S. Inhibition of endogenous HIF inactivation induces angiogenesis in ischaemic skeletal muscles of mice. J. Physiol. 2004, 560, 21–26. [Google Scholar] [CrossRef]
- Xing, J.; Gao, Z.; Lu, J.; Sinoway, L.I.; Li, J. Femoral artery occlusion augments TRPV1-mediated sympathetic responsiveness. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1262–H1269. [Google Scholar] [CrossRef]
- Xing, J.; Lu, J.; Li, J. Contribution of nerve growth factor to augmented TRPV1 responses of muscle sensory neurons by femoral artery occlusion. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1380–H1387. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Lu, J.; Li, J. ASIC3 Function and Immunolabeling Increases in Skeletal Muscle Sensory Neurons Following Femoral Artery Occlusion. J. Physiol. 2012, 590, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Farinelli, S.E.; Greene, L.A. Cell cycle blockers mimosine, ciclopirox, and deferoxamine prevent the death of PC12 cells and postmitotic sympathetic neurons after removal of trophic support. J. Neurosci. 1996, 16, 1150–1162. [Google Scholar] [CrossRef]
- Lomb, D.J.; Straub, J.A.; Freeman, R.S. Prolyl hydroxylase inhibitors delay neuronal cell death caused by trophic factor deprivation. J. Neurochem. 2007, 103, 1897–1906. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Johnson, R.S.; Freeman, R.S. Inhibition of NGF deprivation-induced death by low oxygen involves suppression of BIMEL and activation of HIF-1. J. Cell Biol. 2005, 168, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Ruan, T.; Lin, Y.S.; Lin, K.S.; Kou, Y.R. Sensory transduction of pulmonary reactive oxygen species by capsaicin-sensitive vagal lung afferent fibres in rats. J. Physiol. 2005, 565, 563–578. [Google Scholar] [CrossRef] [PubMed]
- Ruan, T.; Lin, Y.S.; Lin, K.S.; Kou, Y.R. Mediator mechanisms involved in TRPV1 and P2X receptor-mediated, ROS-evoked bradypneic reflex in anesthetized rats. J. Appl. Physiol. 2006, 101, 644–654. [Google Scholar] [CrossRef]
- Wang, H.J.; Pan, Y.X.; Wang, W.Z.; Zucker, I.H.; Wang, W. NADPH oxidase-derived reactive oxygen species in skeletal muscle modulates the exercise pressor reflex. J. Appl. Physiol. 2009, 107, 450–459. [Google Scholar] [CrossRef]
- McCord, J.L.; Tsuchimochi, H.; Yamauchi, K.; Leal, A.; Kaufman, M.P. Tempol attenuates the exercise pressor reflex independently of neutralizing reactive oxygen species in femoral artery ligated rats. J. Appl. Physiol. 2011, 111, 971–979. [Google Scholar] [CrossRef]
- Yamauchi, K.; Stone, A.J.; Stocker, S.D.; Kaufman, M.P. Blockade of ATP-sensitive potassium channels prevents the attenuation of the exercise pressor reflex by tempol in rats with ligated femoral arteries. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H332–H340. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Bessac, B.F.; Sivula, M.; von Hehn, C.A.; Escalera, J.; Cohn, L.; Jordt, S.-E. TRPA1 is a major oxidant sensor in murine airway sensory neurons. J. Clin. Investig. 2008, 118, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Trevisani, M.; Siemens, J.; Materazzi, S.; Bautista, D.M.; Nassini, R.; Campi, B.; Imamachi, N.; Andre, E.; Patacchini, R.; Cottrell, G.S.; et al. 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc. Natl. Acad. Sci. USA 2007, 104, 13519–13524. [Google Scholar] [CrossRef]
- Bautista, D.M.; Jordt, S.-E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef]
- Katsura, H.; Obata, K.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Sakagami, M.; Noguchi, K. Antisense knock down of TRPA1, but not TRPM8, alleviates cold hyperalgesia after spinal nerve ligation in rats. Exp. Neurol. 2006, 200, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.-S.; Woolf, C.J.; Corey, D.P. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. [Erratum appears in J. Clin. Investig. 2010 Jan;120(1):394]. J. Clin. Investig. 2005, 115, 2393–2401. [Google Scholar] [CrossRef]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef]
- Koba, S.; Hayes, S.G.; Sinoway, L.I. Transient receptor potential A1 channel contributes to activation of the muscle reflex. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H201–H213. [Google Scholar] [CrossRef]
- Böhm, F.; Pernow, J. The importance of endothelin-1 for vascular dysfunction in cardiovascular disease. Cardiovasc. Res. 2007, 76, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Sokolovsky, M. Endothelin receptor subtypes and their role in transmembrane signaling mechanisms. Pharmacol. Ther. 1995, 68, 435–471. [Google Scholar] [CrossRef]
- Yeager, M.E.; Belchenko, D.D.; Nguyen, C.M.; Colvin, K.L.; Ivy, D.D.; Stenmark, K.R. Endothelin-1, the unfolded protein response, and persistent inflammation: Role of pulmonary artery smooth muscle cells. Am. J. Respir Cell Mol. Biol. 2012, 46, 14–22. [Google Scholar] [CrossRef]
- Chen, H.C.; Guh, J.Y.; Shin, S.J.; Tsai, J.H.; Lai, Y.H. Reactive oxygen species enhances endothelin-1 production of diabetic rat glomeruli in vitro and in vivo. J. Lab. Clin. Med. 2000, 135, 309–315. [Google Scholar] [CrossRef]
- Ruef, J.; Moser, M.; Kubler, W.; Bode, C. Induction of endothelin-1 expression by oxidative stress in vascular smooth muscle cells. Cardiovasc. Pathol. 2001, 10, 311–315. [Google Scholar] [CrossRef]
- De Haro Miralles, J.; Gonzalez, A.F.; Varela Casariego, C.; Garcia, F.A. Onset of peripheral arterial disease: Role of endothelin in endothelial dysfunction. Interact. Cardiovasc. Thorac. Surg. 2010, 10, 760–765. [Google Scholar] [CrossRef]
- Mangiafico, R.A.; Malatino, L.S.; Santonocito, M.; Sarnataro, F.; Dell’Arte, S.; Messina, R.; Santangelo, B. Plasma endothelin-1 levels in patients with peripheral arterial occlusive disease at different Fontaine’s stages. Panminerva Med. 1999, 41, 22–26. [Google Scholar]
- Qin, L.; Li, J. Heat treatment attenuates activities of ET-1 signaling pathway and enhances the protective HSP72-SOD2 in ischemic skeletal muscle of rats with peripheral artery disease. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Holwerda, S.W.; Restaino, R.M.; Fadel, P.J. Adrenergic and non-adrenergic control of active skeletal muscle blood flow: Implications for blood pressure regulation during exercise. Auton. Neurosci. Basic Clin. 2015, 188, 24–31. [Google Scholar] [CrossRef]
- Pomonis, J.D.; Rogers, S.D.; Peters, C.M.; Ghilardi, J.R.; Mantyh, P.W. Expression and localization of endothelin receptors: Implications for the involvement of peripheral glia in nociception. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 999–1006. [Google Scholar] [CrossRef]
- Motta, E.M.; Calixto, J.B.; Rae, G.A. Mechanical hyperalgesia induced by endothelin-1 in rats is mediated via phospholipase C, protein kinase C, and MAP kinases. Exp. Biol. Med. 2006, 231, 1141–1145. [Google Scholar]
- Motta, E.M.; Chichorro, J.G.; D’Orléans-Juste, P.; Rae, G.A. Roles of endothelin ETA and ETB receptors in nociception and chemical, thermal and mechanical hyperalgesia induced by endothelin-1 in the rat hindpaw. Peptides 2009, 30, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Hasue, F.; Kuwaki, T.; Kisanuki, Y.Y.; Yanagisawa, M.; Moriya, H.; Fukuda, Y.; Shimoyama, M. Increased sensitivity to acute and persistent pain in neuron-specific endothelin-1 knockout mice. Neuroscience 2005, 130, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Barr, T.P.; Kam, S.; Khodorova, A.; Montmayeur, J.P.; Strichartz, G.R. New perspectives on the endothelin axis in pain. Pharmacol. Res. 2011, 63, 532–540. [Google Scholar] [CrossRef]
- Feng, B.; Strichartz, G. Endothelin-1 raises excitability and reduces potassium currents in sensory neurons. Brain Res. Bull. 2009, 79, 345–350. [Google Scholar] [CrossRef]
- Zhou, Z.; Davar, G.; Strichartz, G. Endothelin-1 (ET-1) selectively enhances the activation gating of slowly inactivating tetrodotoxin-resistant sodium currents in rat sensory neurons: A mechanism for the pain-inducing actions of ET-1. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 6325–6330. [Google Scholar] [CrossRef] [Green Version]
- Mule, N.K.; Singh, J.N.; Shah, K.U.; Gulati, A.; Sharma, S.S. Endothelin-1 Decreases Excitability of the Dorsal Root Ganglion Neurons via ET(B) Receptor. Mol. Neurobiol. 2018, 55, 4297–4310. [Google Scholar] [CrossRef]
- Werner, M.F.; Trevisani, M.; Campi, B.; André, E.; Geppetti, P.; Rae, G.A. Contribution of peripheral endothelin ETA and ETB receptors in neuropathic pain induced by spinal nerve ligation in rats. Eur. J. Pain 2010, 14, 911–917. [Google Scholar] [CrossRef]
- Miller, R.J.; Jung, H.; Bhangoo, S.; White, F.A. Cytokine and chemokine regulation of sensory neuron function. In Handbook of Experimental Pharmacology; Canning, B.J., Spina, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 194, pp. 417–449. [Google Scholar]
- Oppenheim, J.J. Cytokines: Past, present, and future. Int. J. Hematol. 2001, 74, 3–8. [Google Scholar] [CrossRef]
- Jonsdottir, I.H.; Schjerling, P.; Ostrowski, K.; Asp, S.; Richter, E.A.; Pedersen, B.K. Muscle contractions induce interleukin-6 mRNA production in rat skeletal muscles. J. Physiol. 2000, 528, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; van Hall, G.; Osada, T.; Sacchetti, M.; Saltin, B.; Pedersen, B.K. Production of interleukin-6 in contracting human skeletal muscles can account for the exercise-induced increase in plasma interleukin-6. J. Physiol. 2000, 529, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.L. Inflammatory mediators and the failing heart: Past, present, and the foreseeable future. Circ. Res. 2002, 91, 988–998. [Google Scholar] [CrossRef] [PubMed]
- El-Menyar, A.A. Cytokines and myocardial dysfunction: State of the art. J. Card Fail. 2008, 14, 61–74. [Google Scholar] [CrossRef]
- Yndestad, A.; Damas, J.K.; Oie, E.; Ueland, T.; Gullestad, L.; Aukrust, P. Systemic inflammation in heart failure–the whys and wherefores. Heart Fail. Rev. 2006, 11, 83–92. [Google Scholar] [CrossRef]
- Lai, J.; Porreca, F.; Hunter, J.C.; Gold, M.S. Voltage-gated sodium channels and hyperalgesia. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 371–397. [Google Scholar] [CrossRef]
- Stone, A.J.; Kim, J.S.; Yamauchi, K.; Ruiz-Velasco, V.; Kaufman, M.P. Attenuation of autonomic reflexes by A803467 may not be solely caused by blockade of NaV 1.8 channels. Neurosci. Lett. 2013, 543, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Pang, R.P.; Shen, K.F.; Zimmermann, M.; Xin, W.J.; Li, Y.Y.; Liu, X.G. TNF-alpha enhances the currents of voltage gated sodium channels in uninjured dorsal root ganglion neurons following motor nerve injury. Exp. Neurol. 2011, 227, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Garbers, C.; Rose-John, S. Interleukin-6: From basic biology to selective blockade of pro-inflammatory activities. Semin. Immunol. 2014, 26, 2–12. [Google Scholar] [CrossRef]
- Chaparala, R.P.; Orsi, N.M.; Lindsey, N.J.; Girn, R.S.; Homer-Vanniasinkam, S. Inflammatory profiling of peripheral arterial disease. Ann. Vasc. Surg. 2009, 23, 172–178. [Google Scholar] [CrossRef]
- Girn, H.R.; Orsi, N.M.; Homer-Vanniasinkam, S. An overview of cytokine interactions in atherosclerosis and implications for peripheral arterial disease. Vasc. Med. 2007, 12, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Palmer-Kazen, U.; Religa, P.; Wahlberg, E. Exercise in patients with intermittent claudication elicits signs of inflammation and angiogenesis. Eur. J. Vasc. Endovasc. Surg. 2009, 38, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.S.; Mazzarino, M.C.; Di Pino, L.; Malaponte, G.; Porto, C.; Pennisi, G.; Marchese, G.; Costa, M.P.; Digrandi, D.; Celotta, G.; et al. High circulating levels of cytokines (IL-6 and TNFalpha), adhesion molecules (VCAM-1 and ICAM-1) and selectins in patients with peripheral arterial disease at rest and after a treadmill test. Vasc. Med. 2003, 8, 15–19. [Google Scholar] [CrossRef]
- Shill, D.D.; Polley, K.R.; Willingham, T.B.; Call, J.A.; Murrow, J.R.; McCully, K.K.; Jenkins, N.T. Experimental intermittent ischemia augments exercise-induced inflammatory cytokine production. J. Appl. Physiol. (1985) 2017, 123, 434–441. [Google Scholar] [CrossRef]
- Copp, S.W.; Stone, A.J.; Li, J.; Kaufman, M.P. Role played by interleukin-6 in evoking the exercise pressor reflex in decerebrate rats: Effect of femoral artery ligation. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H166–H173. [Google Scholar] [CrossRef]
- Jostock, T.; Müllberg, J.; Ozbek, S.; Atreya, R.; Blinn, G.; Voltz, N.; Fischer, M.; Neurath, M.F.; Rose-John, S. Soluble gp130 is the natural inhibitor of soluble interleukin-6 receptor transsignaling responses. Eur. J. Biochem. 2001, 268, 160–167. [Google Scholar] [CrossRef]
- Wolf, J.; Rose-John, S.; Garbers, C. Interleukin-6 and its receptors: A highly regulated and dynamic system. Cytokine 2014, 70, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Fakhry, F.; Spronk, S.; van der Laan, L.; Wever, J.J.; Teijink, J.A.; Hoffmann, W.H.; Smits, T.M.; van Brussel, J.P.; Stultiens, G.N.; Derom, A.; et al. Endovascular Revascularization and Supervised Exercise for Peripheral Artery Disease and Intermittent Claudication: A Randomized Clinical Trial. JAMA 2015, 314, 1936–1944. [Google Scholar] [CrossRef]
- Monroe, J.C.; Song, Q.; Emery, M.S.; Hirai, D.M.; Motaganahalli, R.L.; Roseguini, B.T. Acute effects of leg heat therapy on walking performance and cardiovascular and inflammatory responses to exercise in patients with peripheral artery disease. Physiol. Rep. 2021, 8, e14650. [Google Scholar] [CrossRef]
- Neff, D.; Kuhlenhoelter, A.M.; Lin, C.; Wong, B.J.; Motaganahalli, R.L.; Roseguini, B.T. Thermotherapy reduces blood pressure and circulating endothelin-1 concentration and enhances leg blood flow in patients with symptomatic peripheral artery disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R392–R400. [Google Scholar] [CrossRef]
- Kim, K.; Reid, B.A.; Ro, B.; Casey, C.A.; Song, Q.; Kuang, S.; Roseguini, B.T. Heat therapy improves soleus muscle force in a model of ischemia-induced muscle damage. J. Appl. Physiol. (1985) 2019, 127, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Ro, B.; Damen, F.W.; Gramling, D.P.; Lehr, T.D.; Song, Q.F.; Goergen, C.J.; Roseguini, B.T. Heat therapy improves body composition and muscle function but does not affect capillary or collateral growth in a model of obesity and hindlimb ischemia. J. Appl. Physiol. 2021, 130, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.; Proppe, D.W. Cardiovascular adjustments to heat stress. In Handbook of Physiology—Environmental Physiology; Fregly, M.J., Blatteis, C.M., Eds.; Oxford University Press: New York, NY, USA, 1996; pp. 215–243. [Google Scholar]
- Brothers, R.M.; Bhella, P.S.; Shibata, S.; Wingo, J.E.; Levine, B.D.; Crandall, C.G. Cardiac systolic and diastolic function during whole body heat stress. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1150–H1156. [Google Scholar] [CrossRef]
- Crandall, C.G.; Wilson, T.E.; Marving, J.; Vogelsang, T.W.; Kjaer, A.; Hesse, B.; Secher, N.H. Effects of passive heating on central blood volume and ventricular dimensions in humans. J. Physiol. 2008, 586, 293–301. [Google Scholar] [CrossRef]
- Tei, C.; Horikiri, Y.; Park, J.C.; Jeong, J.W.; Chang, K.S.; Toyama, Y.; Tanaka, N. Acute hemodynamic improvement by thermal vasodilation in congestive heart failure. Circulation 1995, 91, 2582–2590. [Google Scholar] [CrossRef]
- Kisanuki, A.; Daitoku, S.; Kihara, T.; Otsuji, Y.; Tei, C. Thermal therapy improves left ventricular diastolic function in patients with congestive heart failure: A tissue doppler echocardiographic study. J. Cardiol. 2007, 49, 187–191. [Google Scholar] [PubMed]
- Michalsen, A.; Ludtke, R.; Buhring, M.; Spahn, G.; Langhorst, J.; Dobos, G.J. Thermal hydrotherapy improves quality of life and hemodynamic function in patients with chronic heart failure. Am. Heart J. 2003, 146, E11. [Google Scholar] [CrossRef]
- Biro, S.; Masuda, A.; Kihara, T.; Tei, C. Clinical implications of thermal therapy in lifestyle-related diseases. Exp. Biol. Med. 2003, 228, 1245–1249. [Google Scholar] [CrossRef]
- Weber, A.A.; Silver, M.A. Heat therapy in the management of heart failure. Congest. Heart Fail. 2007, 13, 81–83. [Google Scholar] [CrossRef]
- Ikeda, Y.; Biro, S.; Kamogawa, Y.; Yoshifuku, S.; Eto, H.; Orihara, K.; Yu, B.; Kihara, T.; Miyata, M.; Hamasaki, S.; et al. Repeated sauna therapy increases arterial endothelial nitric oxide synthase expression and nitric oxide production in cardiomyopathic hamsters. Circ. J. 2005, 69, 722–729. [Google Scholar] [CrossRef]
- Imamura, M.; Biro, S.; Kihara, T.; Yoshifuku, S.; Takasaki, K.; Otsuji, Y.; Minagoe, S.; Toyama, Y.; Tei, C. Repeated thermal therapy improves impaired vascular endothelial function in patients with coronary risk factors. J. Am. Coll. Cardiol. 2001, 38, 1083–1088. [Google Scholar] [CrossRef]
- Kihara, T.; Biro, S.; Imamura, M.; Yoshifuku, S.; Takasaki, K.; Ikeda, Y.; Otuji, Y.; Minagoe, S.; Toyama, Y.; Tei, C. Repeated sauna treatment improves vascular endothelial and cardiac function in patients with chronic heart failure. J. Am. Coll. Cardiol. 2002, 39, 754–759. [Google Scholar] [CrossRef]
- Thomas, K.N.; van Rij, A.M.; Lucas, S.J.; Cotter, J.D. Lower-limb hot-water immersion acutely induces beneficial hemodynamic and cardiovascular responses in peripheral arterial disease and healthy, elderly controls. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R281–R291. [Google Scholar] [CrossRef] [PubMed]
- Tei, C.; Shinsato, T.; Miyata, M.; Kihara, T.; Hamasaki, S. Waon therapy improves peripheral arterial disease. J. Am. Coll. Cardiol. 2007, 50, 2169–2171. [Google Scholar] [CrossRef]
- Shinsato, T.; Miyata, M.; Kubozono, T.; Ikeda, Y.; Fujita, S.; Kuwahata, S.; Akasaki, Y.; Hamasaki, S.; Fujiwara, H.; Tei, C. Waon therapy mobilizes CD34+ cells and improves peripheral arterial disease. J. Cardiol. 2010, 56, 361–366. [Google Scholar] [CrossRef]
- Cui, J.; Mascarenhas, V.; Moradkhan, R.; Blaha, C.; Sinoway, L.I. Effects of muscle metabolites on responses of muscle sympathetic nerve activity to mechanoreceptor(s) stimulation in healthy humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R458–R466. [Google Scholar] [CrossRef]
- Cui, J.; Blaha, C.; Sinoway, L.I. Whole body heat stress attenuates the pressure response to muscle metaboreceptor stimulation in humans. J. Appl. Physiol. (1985) 2016, 121, 1178–1186. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Shibasaki, M.; Low, D.A.; Keller, D.M.; Davis, S.L.; Crandall, C.G. Heat stress attenuates the increase in arterial blood pressure during the cold pressor test. J. Appl. Physiol. 2010, 109, 1354–1359. [Google Scholar] [CrossRef]
- Sterns, D.A.; Ettinger, S.M.; Gray, K.S.; Whisler, S.K.; Mosher, T.J.; Smith, M.B.; Sinoway, L.I. Skeletal muscle metaboreceptor exercise responses are attenuated in heart failure. Circulation 1991, 84, 2034–2039. [Google Scholar] [CrossRef]
- Ray, C.A.; Gracey, K.H. Augmentation of exercise-induced muscle sympathetic nerve activity during muscle heating. J. Appl. Physiol. 1997, 82, 1719–1725. [Google Scholar] [CrossRef]
- Kuipers, N.T.; Sauder, C.L.; Kearney, M.L.; Ray, C.A. Interactive effect of aging and local muscle heating on renal vasoconstriction during isometric handgrip. Am. J. Physiol. Renal. Physiol. 2009, 297, F327–F332. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, N.T.; Sauder, C.L.; Kearney, M.L.; Ray, C.A. Changes in forearm muscle temperature alter renal vascular responses to isometric handgrip. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3432–H3439. [Google Scholar] [CrossRef] [PubMed]
- Hertel, H.C.; Howaldt, B.; Mense, S. Responses of group IV and group III muscle afferents to thermal stimuli. Brain Res. 1976, 113, 201–205. [Google Scholar] [CrossRef]
- Kumazawa, T.; Mizumura, K. Thin-fibre receptors responding to mechanical, chemical, and thermal stimulation in the skeletal muscle of the dog. J. Physiol. 1977, 273, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Kehoe, V.; Xing, J.; Sinoway, L.; Li, J. Temperature modulates P2X receptor-mediated cardiovascular responses to muscle afferent activation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1255–H1261. [Google Scholar] [CrossRef]
- Bombor, I.; Wissgott, C.; Andresen, R. Lumbar sympathicolysis in patients with severe peripheral artery disease: Hemodynamics of the lower limbs determined by near-infrared spectroscopy, color coded duplex sonography, and temperature measurement. Clin. Med. Insights Cardiol. 2014, 8, 29–36. [Google Scholar] [CrossRef]
- Gray, S.R.; De Vito, G.; Nimmo, M.A.; Farina, D.; Ferguson, R.A. Skeletal muscle ATP turnover and muscle fiber conduction velocity are elevated at higher muscle temperatures during maximal power output development in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R376–R382. [Google Scholar] [CrossRef] [Green Version]
- Dakshinamurti, K.; Dakshinamurti, S. Blood pressure regulation and micronutrients. Nutr. Res. Rev. 2001, 14, 3–44. [Google Scholar] [CrossRef]
- Wilmink, A.B.; Welch, A.A.; Quick, C.R.; Burns, P.J.; Hubbard, C.S.; Bradbury, A.W.; Day, N.E. Dietary folate and vitamin B6 are independent predictors of peripheral arterial occlusive disease. J. Vasc. Surg. 2004, 39, 513–516. [Google Scholar] [CrossRef]
- Khakh, B.S.; Humphrey, P.P.; Surprenant, A. Electrophysiological properties of P2X-purinoceptors in rat superior cervical, nodose and guinea-pig coeliac neurones. J. Physiol. 1995, 484, 385–395. [Google Scholar] [CrossRef]
- He, D.D.; Gao, Y.; Wang, S.; Xie, Z.; Song, X.J. Systematic Administration of B Vitamins Alleviates Diabetic Pain and Inhibits Associated Expression of P2X3 and TRPV1 in Dorsal Root Ganglion Neurons and Proinflammatory Cytokines in Spinal Cord in Rats. Pain. Res. Manag. 2020, 2020, 3740162. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Leuenberger, U.A.; Blaha, C.; King, N.C.; Sinoway, L.I. Effect of P2 receptor blockade with pyridoxine on sympathetic response to exercise pressor reflex in humans. J. Physiol. 2011, 589, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.B.; Gambardella, J.; Castellanos, V.; Trimarco, V.; Santulli, G. Vitamin C and Cardiovascular Disease: An Update. Antioxidants 2020, 9, 1227. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, L.; Cui, J.; Li, J. Sympathetic Nerve Activity and Blood Pressure Response to Exercise in Peripheral Artery Disease: From Molecular Mechanisms, Human Studies, to Intervention Strategy Development. Int. J. Mol. Sci. 2022, 23, 10622. https://doi.org/10.3390/ijms231810622
Qin L, Cui J, Li J. Sympathetic Nerve Activity and Blood Pressure Response to Exercise in Peripheral Artery Disease: From Molecular Mechanisms, Human Studies, to Intervention Strategy Development. International Journal of Molecular Sciences. 2022; 23(18):10622. https://doi.org/10.3390/ijms231810622
Chicago/Turabian StyleQin, Lu, Jian Cui, and Jianhua Li. 2022. "Sympathetic Nerve Activity and Blood Pressure Response to Exercise in Peripheral Artery Disease: From Molecular Mechanisms, Human Studies, to Intervention Strategy Development" International Journal of Molecular Sciences 23, no. 18: 10622. https://doi.org/10.3390/ijms231810622
APA StyleQin, L., Cui, J., & Li, J. (2022). Sympathetic Nerve Activity and Blood Pressure Response to Exercise in Peripheral Artery Disease: From Molecular Mechanisms, Human Studies, to Intervention Strategy Development. International Journal of Molecular Sciences, 23(18), 10622. https://doi.org/10.3390/ijms231810622