

Screening of Induced Mutants Led to the Identification of Starch Biosynthetic Genes Associated with Improved Resistant Starch in Wheat

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Mutant Population Screening for Resistant Starch
2.2. High and Low Resistant Starch Mutant Lines
2.3. Study of Yield Realted Traits in the High and Low Resistant Starch Mutant Lines
2.4. Influences on the Morphology of Starch Granules in High and Low Resistant Starch Mutants
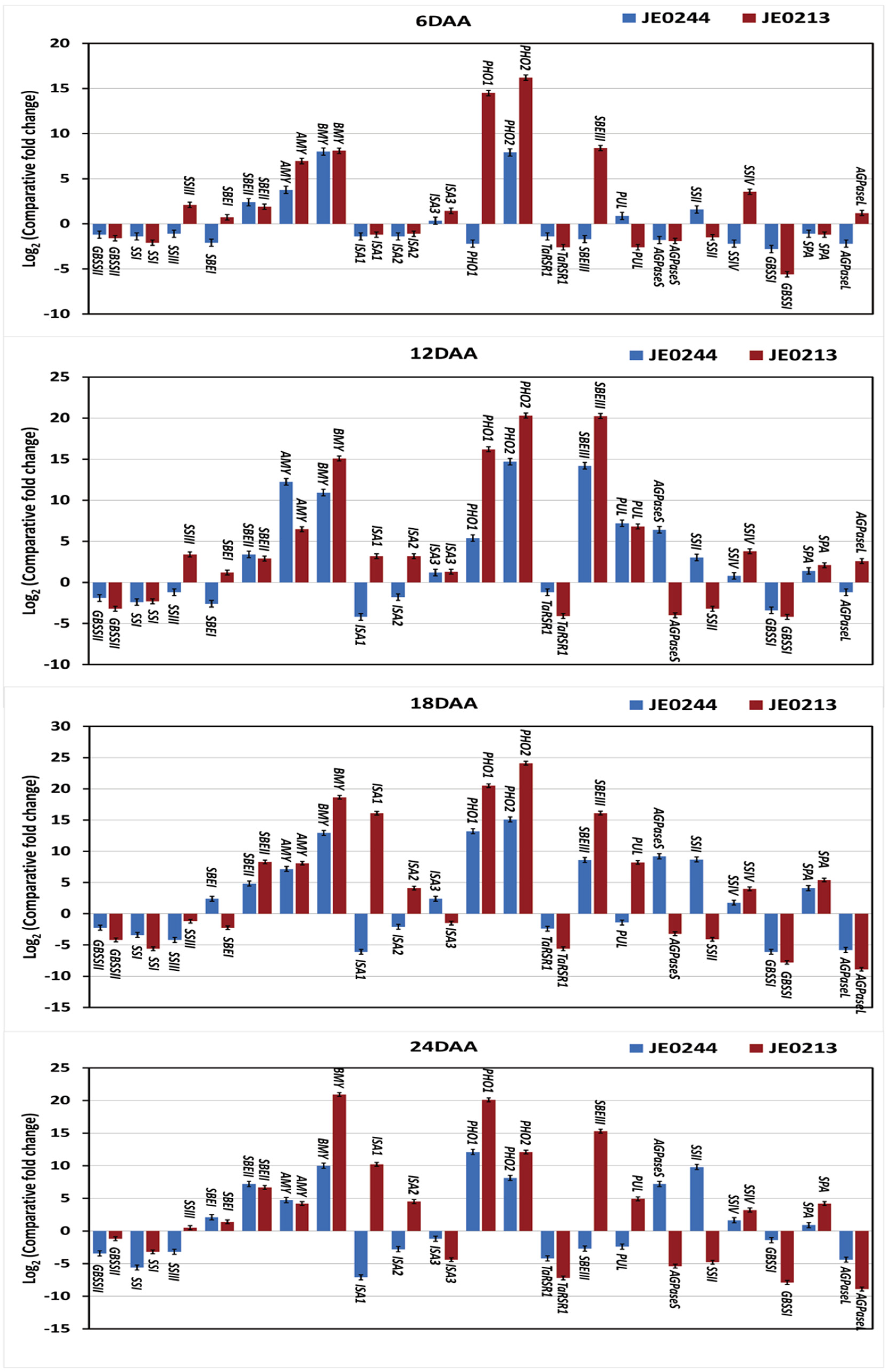
2.5. Expression Analysis for Resitant Starch Mutant Lines
2.6. High Resistant Starch Mutant Lines Expression Analysis of Starch Metabolic Genes
2.7. Low Resistant Starch Mutant Lines Expression Analysis of Starch Metabolic Genes
2.8. Identification of SNPs and InDels in High and Low Resistant Starch Mutant Lines
3. Discussion
4. Material and Method
4.1. Plant Material
4.2. Measurement of Resistant Starch and Digestible Starch
= ΔA × F × EV/W × 0.90
= ΔA × F × EV/W × 0.90
4.3. Determination of Total Starch
= ΔA × F × EV × D/W × 0.90
4.4. Protein, Gluten Content, and Seed Hardness
4.5. Measurement of Grain Weight, Width, and Length
4.6. Starch Purification and Grain Morphology
4.7. Exome Sequencing of Mutant Lines
4.8. Quantitative Gene Expression Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pfister, B.; Zeeman, S.C.; Rugen, M.D.; Field, R.A.; Ebenhöh, O.; Raguin, A. Theoretical and experimental approaches to understand the biosynthesis of starch granules in a physiological context. Photosynth. Res. 2020, 145, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Irshad, A.; Guo, H.; Rehman, S.U.; Wang, X.; Wang, C.; Raza, A.; Zhou, C.; Li, Y.; Liu, L. Soluble Starch Synthase Enzymes in Cereals: An Updated Review. Agronomy 2021, 11, 1983. [Google Scholar] [CrossRef]
- Guo, H.; Xie, Y.; Zhao, L.; Xiong, H.; Gu, J.; Zhao, S.; Liu, L. Progress of Mutant Resource Development and Tilling on Starch Biosynthesis in Wheat. In Mutation Breeding, Genetic Diversity and Crop Adaptation to Climate Change; Sivasankar, S., Ellis, N., Jankuloski, L., Ingelbrecht, I., Eds.; CABI: Wallingford, UK, 2021; pp. 280–284. ISBN 978-1-78924-909-5. [Google Scholar]
- Wang, Q.; Wang, P.; Xiao, Z. Resistant starch prevents tumorigenesis of dimethylhydrazine-induced colon tumors via regulation of an ER stress-mediated mitochondrial apoptosis pathway. Int. J. Mol. Med. 2018, 41, 1887–1898. [Google Scholar] [CrossRef] [PubMed]
- Meenu, M.; Xu, B. A critical review on anti-diabetic and anti-obesity effects of dietary resistant starch. Crit. Rev. Food Sci. Nutr. 2019, 59, 3019–3031. [Google Scholar] [CrossRef] [PubMed]
- Koh, G.Y.; Rowling, M.J. Resistant starch as a novel dietary strategy to maintain kidney health in diabetes mellitus. Nutr. Rev. 2017, 75, 350–360. [Google Scholar] [CrossRef] [PubMed]
- McRae, M.P. Dietary Fiber Is Beneficial for the Prevention of Cardiovascular Disease: An Umbrella Review of Meta-analyses. J. Chiropr. Med. 2017, 16, 289–299. [Google Scholar] [CrossRef]
- Irshad, A.; Guo, H.; Zhang, S.; Gu, J.; Zhao, L.; Xie, Y.; Xiong, H.; Ding, Y.; Ma, Y.; Liu, L. EcoTILLING Reveals Natural Allelic Variations in Starch Synthesis Key Gene TaSSIV and Its Haplotypes Associated with Higher Thousand Grain Weight. Genes 2019, 10, 307. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, L.; Liu, G.; Meng, X.; Jing, Y.; Shu, X.; Kong, X.; Sun, J.; Yu, H.; Smith, S.M.; et al. Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 12844–12849. [Google Scholar] [CrossRef]
- Miura, S.; Koyama, N.; Crofts, N.; Hosaka, Y.; Abe, M.; Fujita, N. Generation and Starch Characterization of Non-Transgenic BEI and BEIIb Double Mutant Rice (Oryza sativa) with Ultra-High Level of Resistant Starch. Rice 2021, 14, 3. [Google Scholar] [CrossRef]
- Zhao, X.; Jayarathna, S.; Turesson, H.; Fält, A.-S.; Nestor, G.; González, M.N.; Olsson, N.; Beganovic, M.; Hofvander, P.; Andersson, R.; et al. Amylose starch with no detectable branching developed through DNA-free CRISPR-Cas9 mediated mutagenesis of two starch branching enzymes in potato. Sci. Rep. 2021, 11, 4311. [Google Scholar] [CrossRef]
- Luo, S.; Ma, Q.; Zhong, Y.; Jing, J.; Wei, Z.; Zhou, W.; Lu, X.; Tian, Y.; Zhang, P. Editing of the starch branching enzyme gene SBE2 generates high-amylose storage roots in cassava. Plant Mol. Biol. 2022, 108, 429–442. [Google Scholar] [CrossRef]
- Tabassum, J.; Ahmad, S.; Hussain, B.; Mawia, A.; Zeb, A.; Ju, L. Applications and Potential of Genome-Editing Systems in Rice Improvement: Current and Future Perspectives. Agronomy 2021, 11, 1359. [Google Scholar] [CrossRef]
- Wang, H.-P.; Cai, H.-Y.; Zhu, J.-H.; Wei, X.; Zhang, S.; Liu, G.; He, Y.-G.; Li, B.; Xu, L.; Jiao, C.-H.; et al. Dynamic Resistant Starch Accumulation in Contrasting Wheat Genotypes Highlights the Lipid Metabolic Pathway Related to Resistant Starch Synthesis. Agriculture 2022, 12, 308. [Google Scholar] [CrossRef]
- Biselli, C.; Volante, A.; Desiderio, F.; Tondelli, A.; Gianinetti, A.; Finocchiaro, F.; Taddei, F.; Gazza, L.; Sgrulletta, D.; Cattivelli, L.; et al. GWAS for Starch-Related Parameters in Japonica Rice (Oryza sativa L.). Plants 2019, 8, 292. [Google Scholar] [CrossRef]
- Zeng, Y.; Sun, D.; Du, J.; Pu, X.; Yang, S.; Yang, X.; Yang, T.; Yang, J. Identification of QTLs for resistant starch and total alkaloid content in brown and polished rice. Genet. Mol. Res. 2016, 15, 15037268. [Google Scholar] [CrossRef]
- Johnson, N.; Boatwright, J.L.; Bridges, W.; Thavarajah, P.; Kumar, S.; Shipe, E.; Thavarajah, D. Genome-wide association mapping of lentil (Lens culinaris Medikus) prebiotic carbohydrates toward improved human health and crop stress tolerance. Sci. Rep. 2021, 11, 13926. [Google Scholar] [CrossRef]
- Shu, X.; Sun, J.; Wu, D. Effects of grain development on formation of resistant starch in rice. Food Chem. 2014, 164, 89–97. [Google Scholar] [CrossRef]
- Mishra, A.; Singh, A.; Sharma, M.; Kumar, P.; Roy, J. Development of EMS-induced mutation population for amylose and resistant starch variation in bread wheat (Triticum aestivum) and identification of candidate genes responsible for amylose variation. BMC Plant Biol. 2016, 16, 217. [Google Scholar] [CrossRef]
- Till, B.J.; Datta, S.; Jankowicz-Cieslak, J. TILLING: The Next Generation. In Plant Genetics and Molecular Biology; Varshney, R.K., Pandey, M.K., Chitikineni, A., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer International Publishing: Cham, Switzerland, 2018; Volume 164, pp. 139–160. ISBN 978-3-319-91312-4. [Google Scholar]
- Taheri, S.; Abdullah, T.L.; Jain, S.M.; Sahebi, M.; Azizi, P. TILLING, high-resolution melting (HRM), and next-generation sequencing (NGS) techniques in plant mutation breeding. Mol. Breed. 2017, 37, 40. [Google Scholar] [CrossRef]
- Moehs, C.P.; Austill, W.J.; Facciotti, D.; Holm, A.; Loeffler, D.; Lu, Z.; Mullenberg, J.C.; Slade, A.J.; Steine, M.N.; van Boxtel, J.; et al. Development of non-transgenic glyphosate tolerant wheat by TILLING. PLoS ONE 2021, 16, e0245802. [Google Scholar] [CrossRef]
- Hussain, M.; Iqbal, M.A.; Till, B.J.; Rahman, M.-U. Identification of induced mutations in hexaploid wheat genome using exome capture assay. PLoS ONE 2018, 13, e0201918. [Google Scholar] [CrossRef] [PubMed]
- González-Thuillier, I.; Pellny, T.K.; Tosi, P.; Mitchell, R.A.C.; Haslam, R.; Shewry, P.R. Accumulation and deposition of triacylglycerols in the starchy endosperm of wheat grain. J. Cereal Sci. 2021, 98, 103167. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhu, D.; Ma, C.; Cao, H.; Wang, Y.; Xu, Y.; Zhang, W.; Yan, Y. Transcriptome analysis reveals key differentially expressed genes involved in wheat grain development. Crop J. 2016, 4, 92–106. [Google Scholar] [CrossRef]
- Song, Y.; Luo, G.; Shen, L.; Yu, K.; Yang, W.; Li, X.; Sun, J.; Zhan, K.; Cui, D.; Liu, D.; et al. TubZIP28, a novel bZIP family transcription factor from Triticum urartu, and TabZIP28, its homologue from Triticum aestivum, enhance starch synthesis in wheat. New Phytol. 2020, 226, 1384–1398. [Google Scholar] [CrossRef]
- Parween, S.; Anonuevo, J.J.; Butardo, V.M.; Misra, G.; Anacleto, R.; Llorente, C.; Kosik, O.; Romero, M.V.; Bandonill, E.H.; Mendioro, M.S.; et al. Balancing the double-edged sword effect of increased resistant starch content and its impact on rice texture: Its genetics and molecular physiological mechanisms. Plant Biotechnol. J. 2020, 18, 1763–1777. [Google Scholar] [CrossRef]
- Xia, J.; Zhu, D.; Wang, R.; Cui, Y.; Yan, Y. Crop resistant starch and genetic improvement: A review of recent advances. Theor. Appl. Genet. 2018, 131, 2495–2511. [Google Scholar] [CrossRef]
- Zhang, X.; Karim, H.; Feng, X.; Lan, J.; Tang, H.; Guzmán, C.; Xu, Q.; Zhang, Y.; Qi, P.; Deng, M.; et al. A single base change at exon of Wx-A1 caused gene inactivation and starch properties modified in a wheat EMS mutant line. J. Sci. Food Agric. 2022, 102, 2012–2022. [Google Scholar] [CrossRef] [PubMed]
- Gillmor, C.S.; Lukowitz, W. EMS Mutagenesis of Arabidopsis Seeds. In Plant Embryogenesis; Bayer, M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2020; Volume 2122, pp. 15–23. ISBN 978-1-07-160341-3. [Google Scholar]
- Chen, L.; Hu, Y.-G. Using TILLING Lines for Functional Genomics in Wheat. In Accelerated Breeding of Cereal Crops; Bilichak, A., Laurie, J.D., Eds.; Springer Protocols Handbooks; Springer: New York, NY, USA, 2022; pp. 181–193. ISBN 978-1-07-161525-6. [Google Scholar]
- Bovina, R.; Brunazzi, A.; Gasparini, G.; Sestili, F.; Palombieri, S.; Botticella, E.; Lafiandra, D.; Mantovani, P.; Massi, A. Development of a TILLING resource in durum wheat for reverse- and forward-genetic analyses. Crop Pasture Sci. 2014, 65, 112–124. [Google Scholar] [CrossRef]
- Bird, A.R.; Regina, A. High amylose wheat: A platform for delivering human health benefits. J. Cereal Sci. 2018, 82, 99–105. [Google Scholar] [CrossRef]
- Wang, J.; Hu, P.; Chen, Z.; Liu, Q.; Wei, C. Progress in High-Amylose Cereal Crops through Inactivation of Starch Branching Enzymes. Front. Plant Sci. 2017, 8, 469. [Google Scholar] [CrossRef] [Green Version]
- Howard, T.P.; Fahy, B.; Leigh, F.; Howell, P.; Powell, W.; Greenland, A.; Trafford, K.; Smith, A.M. Use of advanced recombinant lines to study the impact and potential of mutations affecting starch synthesis in barley. J. Cereal Sci. 2014, 59, 196–202. [Google Scholar] [CrossRef]
- Zi, Y.; Ding, J.; Song, J.; Humphreys, G.; Peng, Y.; Li, C.; Zhu, X.; Guo, W. Grain Yield, Starch Content and Activities of Key Enzymes of Waxy and Non-waxy Wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4548. [Google Scholar] [CrossRef]
- Asai, H.; Abe, N.; Matsushima, R.; Crofts, N.; Oitome, N.F.; Nakamura, Y.; Fujita, N. Deficiencies in both starch synthase IIIa and branching enzyme IIb lead to a significant increase in amylose in SSIIa-inactive japonica rice seeds. J. Exp. Bot. 2014, 65, 5497–5507. [Google Scholar] [CrossRef]
- Yu, G.; Gaoyang, Y.; Liu, L.; Shoaib, N.; Deng, Y.; Zhang, N.; Li, Y.; Huang, Y. The Structure, Function, and Regulation of Starch Synthesis Enzymes SSIII with Emphasis on Maize. Agronomy 2022, 12, 1359. [Google Scholar] [CrossRef]
- Shufen, C.; Yicong, C.; Baobing, F.; Guiai, J.; Zhonghua, S.; Ju, L.; Shaoqing, T.; Jianlong, W.; Peisong, H.; Xiangjin, W. Editing of Rice Isoamylase Gene ISA1 Provides Insights into Its Function in Starch Formation. Rice Sci. 2019, 26, 77–87. [Google Scholar] [CrossRef]
- Zhao, X.; Andersson, M.; Andersson, R. Resistant starch and other dietary fiber components in tubers from a high-amylose potato. Food Chem. 2018, 251, 58–63. [Google Scholar] [CrossRef]
- Shoaib, N.; Liu, L.; Ali, A.; Mughal, N.; Yu, G.; Huang, Y. Molecular Functions and Pathways of Plastidial Starch Phosphorylase (PHO1) in Starch Metabolism: Current and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 10450. [Google Scholar] [CrossRef]
- Orawetz, T.; Malinova, I.; Orzechowski, S.; Fettke, J. Reduction of the plastidial phosphorylase in potato (Solanum tuberosum L.) reveals impact on storage starch structure during growth at low temperature. Plant Physiol. Biochem. 2016, 100, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Seung, D. Amylose in starch: Towards an understanding of biosynthesis, structure and function. New Phytol. 2020, 228, 1490–1504. [Google Scholar] [CrossRef]
- McMaugh, S.J.; Thistleton, J.L.; Anschaw, E.; Luo, J.; Konik-Rose, C.; Wang, H.; Huang, M.; Larroque, O.; Regina, A.; Jobling, S.A.; et al. Suppression of starch synthase I expression affects the granule morphology and granule size and fine structure of starch in wheat endosperm. J. Exp. Bot. 2014, 65, 2189–2201. [Google Scholar] [CrossRef] [Green Version]
- Brummell, D.A.; Watson, L.M.; Zhou, J.; McKenzie, M.J.; Hallett, I.C.; Simmons, L.; Carpenter, M.; Timmerman-Vaughan, G.M. Overexpression of STARCH BRANCHING ENZYME II increases short-chain branching of amylopectin and alters the physicochemical properties of starch from potato tuber. BMC Biotechnol. 2015, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Chen, Y.; Zhang, M.; Cai, R.; Cheng, B.; Ma, Q.; Zhao, Y. A novel GRAS transcription factor, ZmGRAS20, regulates starch biosynthesis in rice endosperm. Physiol. Mol. Biol. Plants 2017, 23, 143–154. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; Sloane, N.; Draga, A. Determination of total dietary fibre and available carbohydrates: A rapid integrated procedure that simulates in vivo digestion. Starch-Stärke 2015, 67, 860–883. [Google Scholar] [CrossRef]
- Torbica, A.; Horvat, D.; Živančev, D.; Belović, M.; Šimić, G.; Magdić, D.; Đukić, N.; Dvojković, K. Prediction of the genetic similarity of wheat and wheat quality by reversed-phase high-performance liquid chromatography and lab-on-chip methods. Acta Aliment. 2017, 46, 137–144. [Google Scholar] [CrossRef]
- Hawkins, E.; Chen, J.; Watson-Lazowski, A.; Ahn-Jarvis, J.; Barclay, J.E.; Fahy, B.; Hartley, M.; Warren, F.J.; Seung, D. STARCH SYNTHASE 4 is required for normal starch granule initiation in amyloplasts of wheat endosperm. New Phytol. 2021, 230, 2371–2386. [Google Scholar] [CrossRef] [PubMed]
- King, R.; Bird, N.; Ramirez-Gonzalez, R.; Coghill, J.A.; Patil, A.; Hassani-Pak, K.; Uauy, C.; Phillips, A.L. Mutation Scanning in Wheat by Exon Capture and Next-Generation Sequencing. PLoS ONE 2015, 10, e0137549. [Google Scholar] [CrossRef] [PubMed]
- Qidi, D.; Huijun, G.; Hong-Chun, X.; Yong-Dun, X.; Lin-Shu, Z.; Jia-Yu, G.; Shi-Rong, Z.; Yu-Ping, D.; Xi-Yun, S.; Lu-Xiang, L. Gene Mapping of Apical Spikelet Degeneration Mutant Asd1 in Wheat. Acta Agron. Sin. 2022, 48, 1905–1913. [Google Scholar] [CrossRef]
- Singh, A.; Mantri, S.; Sharma, M.; Chaudhury, A.; Tuli, R.; Roy, J. Genome-wide transcriptome study in wheat identified candidate genes related to processing quality, majority of them showing interaction (quality x development) and having temporal and spatial distributions. BMC Genom. 2014, 15, 29. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, P.; Sharma, M.; Tuli, R.; Dhaliwal, H.S.; Chaudhury, A.; Pal, D.; Roy, J. Expression Patterns of Genes Involved in Starch Biosynthesis during Seed Development in Bread Wheat (Triticum Aestivum). Mol. Breed. 2015, 35, 184. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irshad, A.; Guo, H.; Ur Rehman, S.; Gu, J.; Wang, C.; Xiong, H.; Xie, Y.; Zhao, S.; Liu, L. Screening of Induced Mutants Led to the Identification of Starch Biosynthetic Genes Associated with Improved Resistant Starch in Wheat. Int. J. Mol. Sci. 2022, 23, 10741. https://doi.org/10.3390/ijms231810741
Irshad A, Guo H, Ur Rehman S, Gu J, Wang C, Xiong H, Xie Y, Zhao S, Liu L. Screening of Induced Mutants Led to the Identification of Starch Biosynthetic Genes Associated with Improved Resistant Starch in Wheat. International Journal of Molecular Sciences. 2022; 23(18):10741. https://doi.org/10.3390/ijms231810741
Chicago/Turabian StyleIrshad, Ahsan, Huijun Guo, Shoaib Ur Rehman, Jiayu Gu, Chaojie Wang, Hongchun Xiong, Yongdun Xie, Shirong Zhao, and Luxiang Liu. 2022. "Screening of Induced Mutants Led to the Identification of Starch Biosynthetic Genes Associated with Improved Resistant Starch in Wheat" International Journal of Molecular Sciences 23, no. 18: 10741. https://doi.org/10.3390/ijms231810741
APA StyleIrshad, A., Guo, H., Ur Rehman, S., Gu, J., Wang, C., Xiong, H., Xie, Y., Zhao, S., & Liu, L. (2022). Screening of Induced Mutants Led to the Identification of Starch Biosynthetic Genes Associated with Improved Resistant Starch in Wheat. International Journal of Molecular Sciences, 23(18), 10741. https://doi.org/10.3390/ijms231810741