
Healthy-like CD4+ Regulatory and CD4+ Conventional T-Cell Receptor Repertoires Predict Protection from GVHD Following Donor Lymphocyte Infusion

 , , ,
, , ,
Abstract
1. Introduction
2. Results
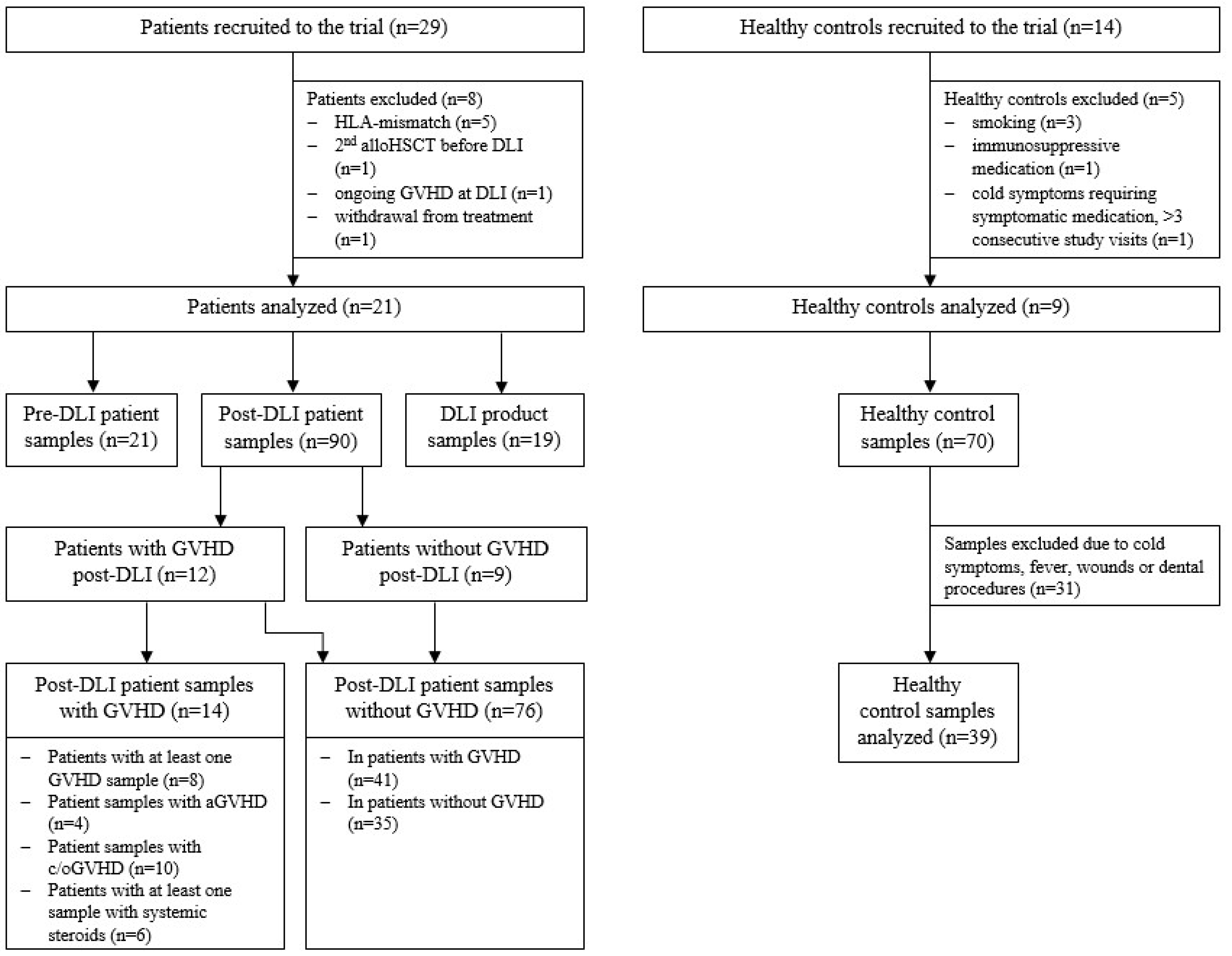
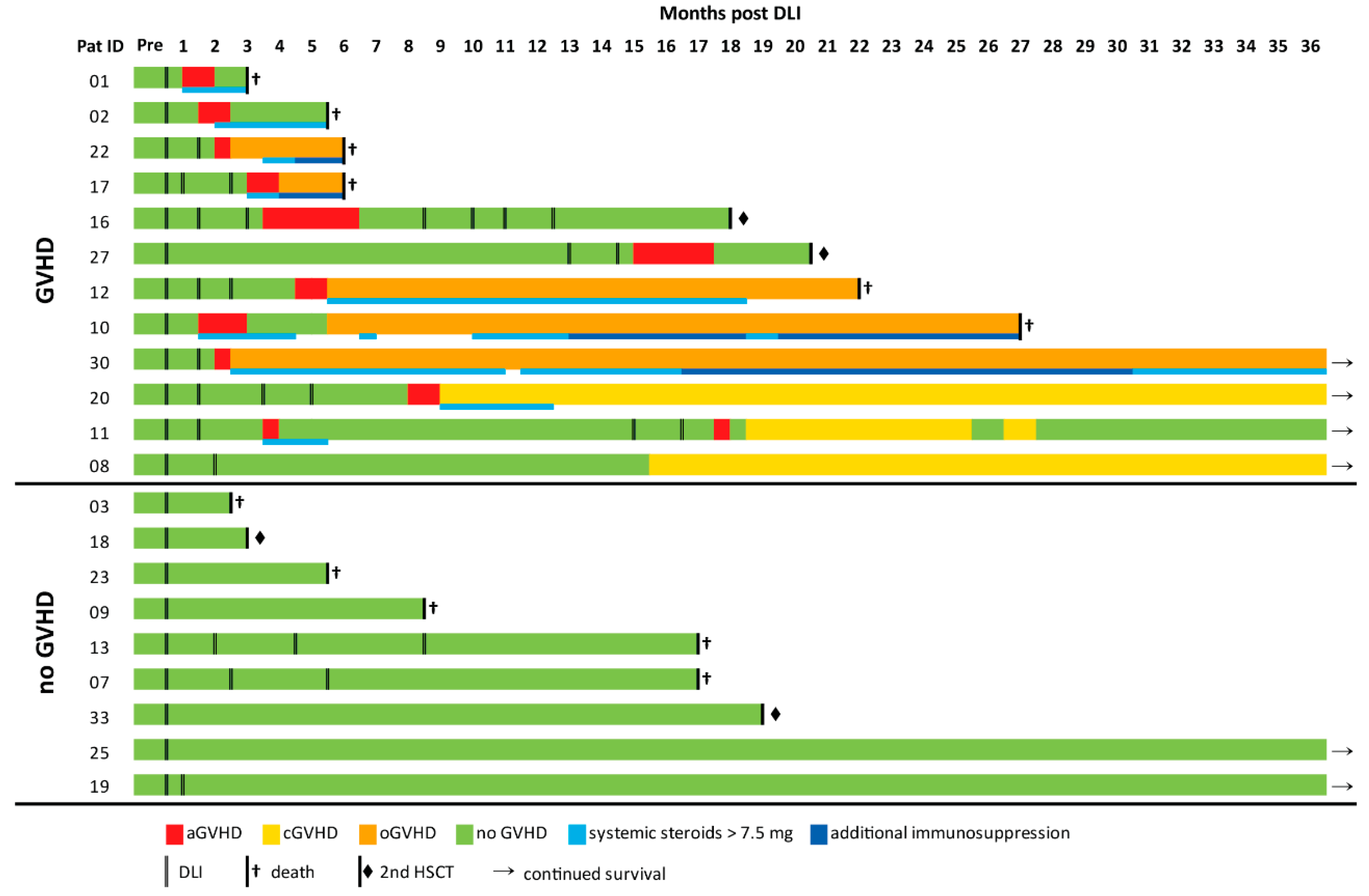
2.1. Cohort Analysis
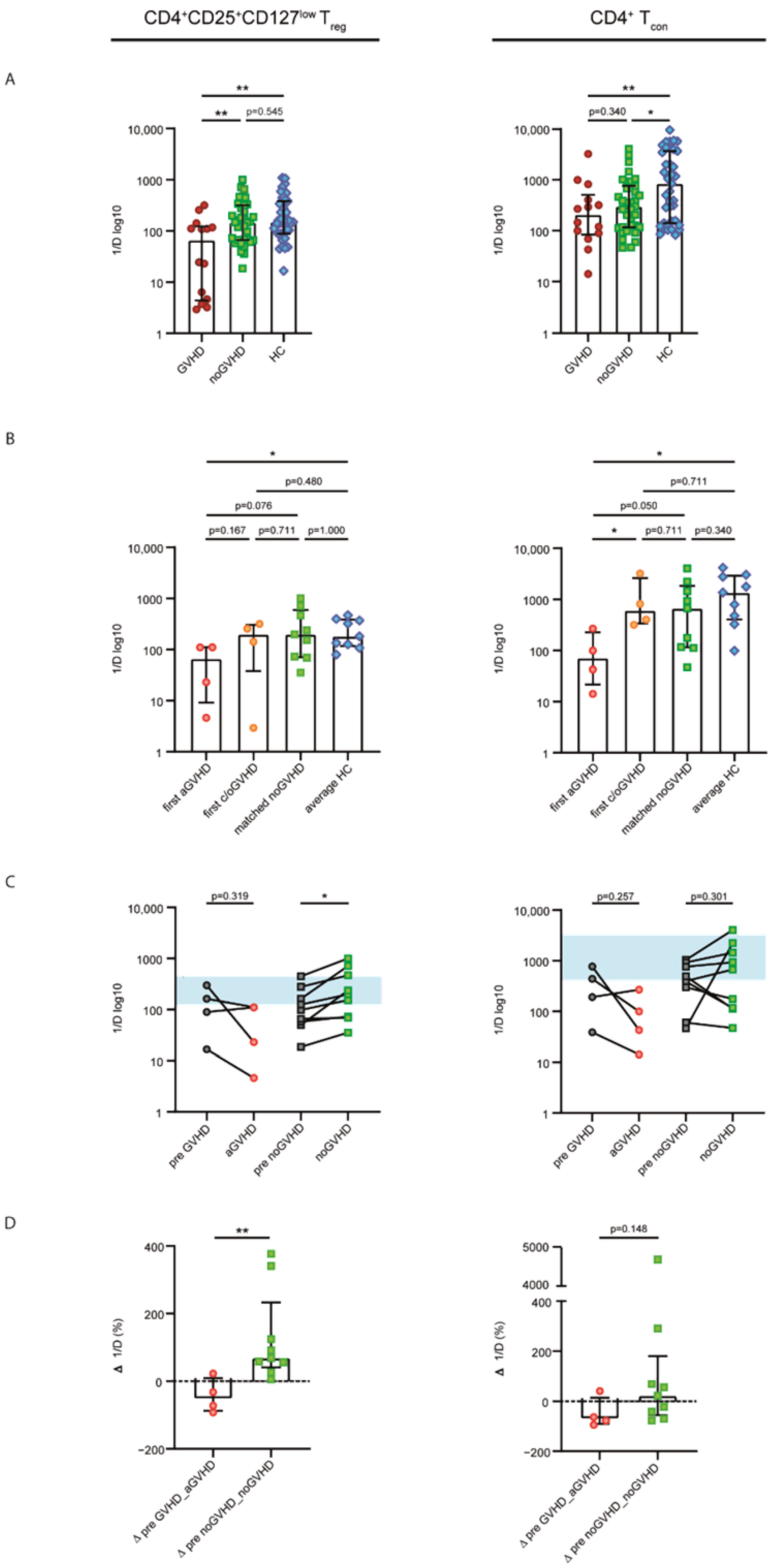
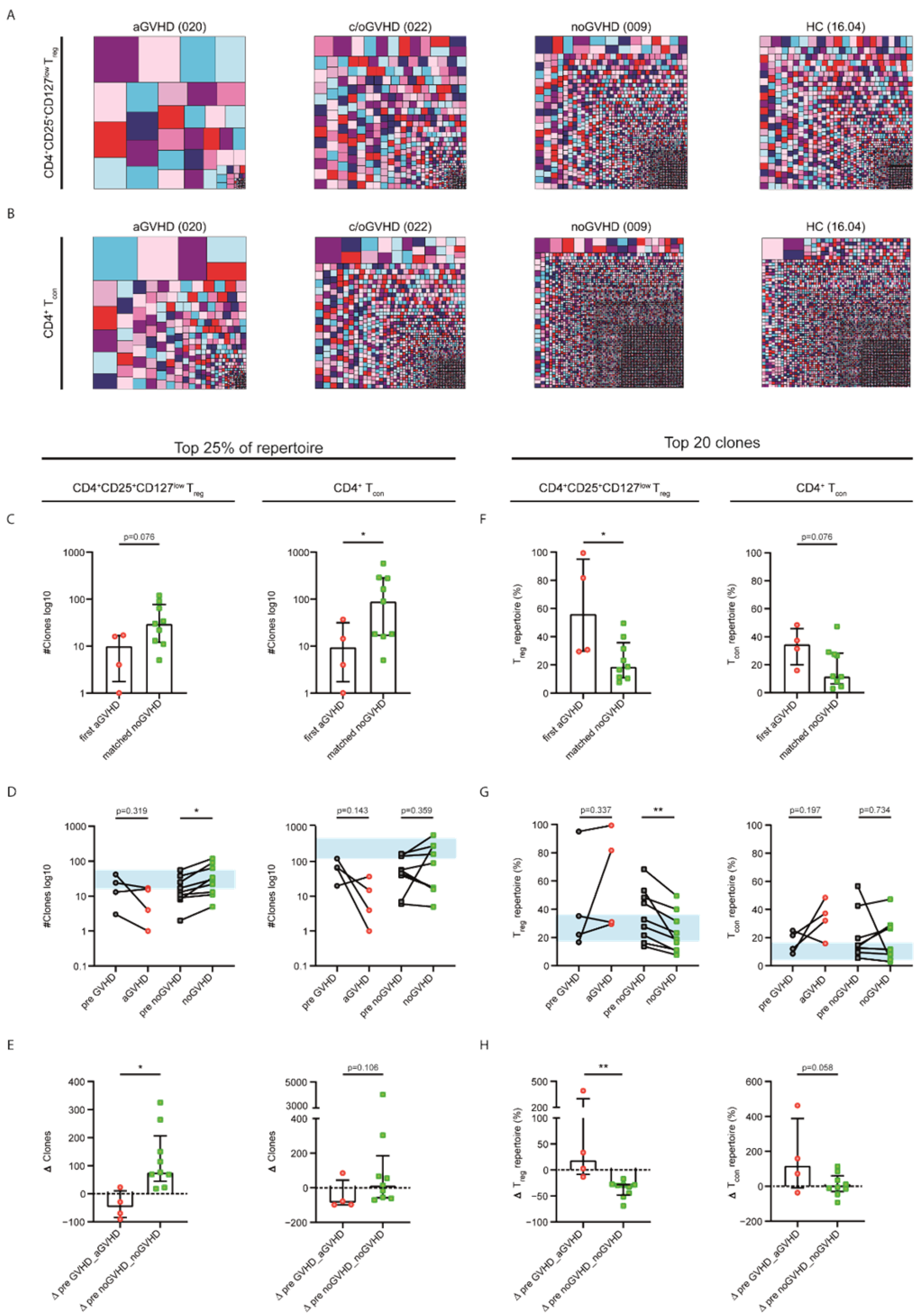
2.2. Patients without GVHD Exhibited a Healthy-LIKE CD4+CD25+CD127low Treg and CD4+ Tcon Repertoire Diversity
2.3. Acute GVHD Was Associated with Clonally Expanded CD4+CD25+CD127low Treg and CD4+ Tcon Repertoires
2.4. Steroid Sensitivity Was Linked to Diversification of the CD4+CD25+CD127low Treg and CD4+ Tcon Repertoires
2.5. Repertoire Properties of DLI Products and Patient Samples Prior to DLI Did Not Differ between GVHD and noGVHD
2.6. Resemblance to Healthy-like Repertoires Early after DLI Coincided with a Lower Incidence of GVHD
3. Discussion
4. Materials and Methods
4.1. Cohort and Study Design
4.2. Flow Cytometry and Cell Sorting
4.3. Next-Generation Sequencing and Sequence Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmid, C.; Kuball, J.; Bug, G. Defining the Role of Donor Lymphocyte Infusion in High-Risk Hematologic Malignancies. J. Clin. Oncol. 2021, 39, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H. Graft-versus-Leukemia Effects of Transplantation and Donor Lymphocytes Pathophysiology of GVHD. ASH 50th Anniv. Rev. 2018, 112, 4371–4384. [Google Scholar] [CrossRef]
- Schmid, C.; Labopin, M.; Nagler, A.; Bornhäuser, M.; Finke, J.; Fassas, A.; Volin, L.; Gürman, G.; Maertens, J.; Bordigoni, P.; et al. Donor Lymphocyte Infusion in the Treatment of First Hematological Relapse after Allogeneic Stem-Cell Transplantation in Adults with Acute Myeloid Leukemia: A Retrospective Risk Factors Analysis and Comparison with Other Strategies by the EBMT Acute Leukem. J. Clin. Oncol. 2007, 25, 4938–4945. [Google Scholar] [CrossRef] [PubMed]
- Schmid, C.; Labopin, M.; Schaap, N.; Veelken, H.; Brecht, A.; Stadler, M.; Finke, J.; Baron, F.; Collin, M.; Bug, G.; et al. Long-Term Results and GvHD after Prophylactic and Preemptive Donor Lymphocyte Infusion after Allogeneic Stem Cell Transplantation for Acute Leukemia. Bone Marrow Transplant. 2022, 57, 215–223. [Google Scholar] [CrossRef]
- Mackinnon, S.; Papadopoulos, E.B.; Carabasi, M.H.; Reich, L.; Collins, N.H.; Boulad, F.; Castro-Malaspina, H.; Childs, B.H.; Gillio, A.P.; Kernan, N.A. Adoptive Immunotherapy Evaluating Escalating Doses of Donor Leukocytes for Relapse of Chronic Myeloid Leukemia after Bone Marrow Transplantation: Separation of Graft-versus-Leukemia Responses from Graft-versus-Host Disease. Blood 1995, 86, 1261–1268. [Google Scholar] [CrossRef]
- Stadler, M.; Venturini, L.; Bünting, I.; Dammann, E.; Weissinger, E.M.; Schwarzer, A.; Schultze-Florey, C.; Ehrlich, S.; Markel, D.; Lueck, C.; et al. Navigating Preemptive and Therapeutic Donor Lymphocyte Infusions in Advanced Myeloid Malignancies by High-Sensitivity Chimerism Analysis. Front. Oncol. 2022, 12, 867356. [Google Scholar] [CrossRef] [PubMed]
- Simula, M.P.; Marktel, S.; Fozza, C.; Kaeda, J.; Szydlo, R.M.; Nadal, E.; Bua, M.; Rahemtulla, A.; Kanfer, E.; Marin, D.; et al. Response to Donor Lymphocyte Infusions for Chronic Myeloid Leukemia Is Dose-Dependent: The Importance of Escalating the Cell Dose to Maximize Therapeutic Efficacy. Leukemia 2007, 21, 943–948. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ortí, G.; Palacio-Garcia, C.; García-Cadenas, I.; Sánchez-Ortega, I.; Jimenez, M.J.; Azqueta, C.; Villacampa, G.; Ferrà, C.; Parody, R.; Martino, R.; et al. Analysis of Cell Subsets in Donor Lymphocyte Infusions from HLA Identical Sibling Donors after Allogeneic Hematopoietic Cell Transplant. Transplant. Cell. Ther. 2021, 27, 53.e1–53.e8. [Google Scholar] [CrossRef] [PubMed]
- Ohkura, N.; Kitagawa, Y.; Sakaguchi, S. Development and Maintenance of Regulatory T Cells. Immunity 2013, 38, 414–423. [Google Scholar] [CrossRef]
- Edinger, M.; Hoffmann, P.; Ermann, J.; Drago, K.; Fathman, C.G.; Strober, S.; Negrin, R.S. CD4+CD25+ Regulatory T Cells Preserve Graft-versus-Tumor Activity While Inhibiting Graft-versus-Host Disease after Bone Marrow Transplantation. Nat. Med. 2003, 9, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.A.; Lamikanra, A.; Dorée, C.; Gration, B.; Tsang, P.; Danby, R.D.; Roberts, D.J. Increased Regulatory T Cell Graft Content Is Associated with Improved Outcome in Haematopoietic Stem Cell Transplantation: A Systematic Review. Br. J. Haematol. 2017, 176, 448–463. [Google Scholar] [CrossRef] [PubMed]
- Maury, S.; Lemoine, F.M.; Hicheri, Y.; Rosenzwajg, M.; Badoual, C.; Cheraï, M.; Beaumont, J.-L.; Azar, N.; Dhedin, N.; Sirvent, A.; et al. CD4+CD25+ Regulatory T Cell Depletion Improves the Graft-Versus-Tumor Effect of Donor Lymphocytes After Allogeneic Hematopoietic Stem Cell Transplantation. Sci. Transl. Med. 2010, 2, 41ra52. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Shiobara, S.; Nakao, S. Identification of T-Cell Clones Showing Expansion Associated with Graft-vs-Leukemia Effect on Chronic Myelogenous Leukemia in Vivo and in Vitro. Exp. Hematol. 2001, 29, 471–476. [Google Scholar] [CrossRef]
- Verfuerth, S.; Peggs, K.; Vyas, P.; Barnett, L.; O’Reilly, R.J.; Mackinnon, S. Longitudinal Monitoring of Immune Reconstitution by CDR3 Size Spectratyping after T-Cell-Depleted Allogeneic Bone Marrow Transplant and the Effect of Donor Lymphocyte Infusions on T-Cell Repertoire. Blood 2000, 95, 3990–3995. [Google Scholar] [CrossRef]
- Claret, E.J.; Alyea, E.P.; Orsini, E.; Pickett, C.C.; Collins, H.; Wang, Y.; Neuberg, D.; Soiffer, R.J.; Ritz, J. Characterization of T Cell Repertoire in Patients with Graft-versus-Leukemia after Donor Lymphocyte Infusion. J. Clin. Investig. 1997, 100, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Ravens, S.; Schultze-Florey, C.; Raha, S.; Sandrock, I.; Drenker, M.; Oberdörfer, L.; Reinhardt, A.; Ravens, I.; Beck, M.; Geffers, R.; et al. Human Γδ T Cells Are Quickly Reconstituted after Stem-Cell Transplantation and Show Adaptive Clonal Expansion in Response to Viral Infection. Nat. Immunol. 2017, 18, 393–401. [Google Scholar] [CrossRef]
- Schultze-Florey, C.R.; Kuhlmann, L.; Raha, S.; Barros-Martins, J.; Odak, I.; Tan, L.; Xiao, Y.; Ravens, S.; Hambach, L.; Venturini, L.; et al. Clonal Expansion of CD8+ T Cells Reflects Graft-versus-Leukemia Activity and Precedes Durable Remission Following DLI. Blood Adv. 2021, 5, 4485–4499. [Google Scholar] [CrossRef] [PubMed]
- van Heijst, J.W.J.; Ceberio, I.; Lipuma, L.B.; Samilo, D.W.; Wasilewski, G.D.; Gonzales, A.M.R.; Nieves, J.L.; van den Brink, M.R.M.; Perales, M.A.; Pamer, E.G. Quantitative Assessment of T Cell Repertoire Recovery after Hematopoietic Stem Cell Transplantation. Nat. Med. 2013, 19, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Kanakry, C.G.; Coffey, D.G.; Towlerton, A.M.H.; Vulic, A.; Storer, B.E.; Chou, J.; Yeung, C.C.S.; Gocke, C.D.; Robins, H.S.; O’Donnell, P.V.; et al. Origin and Evolution of the T Cell Repertoire after Posttransplantation Cyclophosphamide. JCI Insight 2016, 1, e86252. [Google Scholar] [CrossRef] [PubMed]
- Odak, I.; Raha, S.; Schultze-Florey, C.; Tavil, S.; Ravens, S.; Ganser, A.; Förster, R.; Prinz, I.; Koenecke, C. Focusing of the Regulatory T-Cell Repertoire after Allogeneic Stem Cell Transplantation Indicates Protection from Graft-versus-Host Disease. Haematologica 2019, 104, e577–e580. [Google Scholar] [CrossRef] [PubMed]
- Leick, M.; Gittelman, R.M.; Yusko, E.; Sanders, C.; Robins, H.; DeFilipp, Z.; Nikiforow, S.; Ritz, J.; Chen, Y.-B. T Cell Clonal Dynamics Determined by High-Resolution TCR-β Sequencing in Recipients after Allogeneic Hematopoietic Cell Transplantation. Biol. Blood Marrow Transplant. 2020, 26, 1567–1574. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.A.; Haque, M.; Fawaz, M.; Abdeen, H.; Coffey, D.; Towlerton, A.; Abdeen, A.; Toor, A.; Warren, E.; Reed, J.; et al. T Cell Repertoire Evolution after Allogeneic Bone Marrow Transplantation: An Organizational Perspective. Biol. Blood Marrow Transplant. 2019, 25, 868–882. [Google Scholar] [CrossRef] [PubMed]
- Du, J.-W.; Gu, J.-Y.; Liu, J.; Cen, X.-N.; Zhang, Y.; Ou, Y.; Chu, B.; Zhu, P. TCR Spectratyping Revealed T Lymphocytes Associated with Graft-versus-Host Disease after Allogeneic Hematopoietic Stem Cell Transplantation. Leuk. Lymphoma 2007, 48, 1618–1627. [Google Scholar] [CrossRef]
- Liu, C.; He, M.; Rooney, B.; Kepler, T.B.; Chao, N.J. Longitudinal Analysis of T-Cell Receptor Variable Beta Chain Repertoire in Patients with Acute Graft-versus-Host Disease after Allogeneic Stem Cell Transplantation. Biol. Blood Marrow Transplant. 2006, 12, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Meyer, E.H.; Hsu, A.R.; Liliental, J.; Löhr, A.; Florek, M.; Zehnder, J.L.; Strober, S.; Lavori, P.; Miklos, D.B.; Johnson, D.S.; et al. A Distinct Evolution of the T-Cell Repertoire Categorizes Treatment Refractory Gastrointestinal Acute Graft-versus-Host Disease. Blood 2013, 121, 4955–4962. [Google Scholar] [CrossRef] [PubMed]
- Döhner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Büchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and Management of AML in Adults: 2017 ELN Recommendations from an International Expert Panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, P.L.; Tuechler, H.; Schanz, J.; Sanz, G.; Garcia-Manero, G.; Solé, F.; Bennett, J.M.; Bowen, D.; Fenaux, P.; Dreyfus, F.; et al. Revised International Prognostic Scoring System for Myelodysplastic Syndromes. Blood 2012, 120, 2454–2465. [Google Scholar] [CrossRef] [PubMed]
- Föhse, L.; Suffner, J.; Suhre, K.; Wahl, B.; Lindner, C.; Lee, C.; Schmitz, S.; Haas, J.D.; Lamprecht, S.; Koenecke, C.; et al. High TCR Diversity Ensures Optimal Function and Homeostasis of Foxp3+ Regulatory T Cells. Eur. J. Immunol. 2011, 41, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.N.; Meyerson, M. Antigen Identification for HLA Class I- and HLA Class II-Restricted T Cell Receptors Using Cytokine-Capturing Antigen-Presenting Cells. Sci. Immunol. 2021, 6, eabf4001. [Google Scholar] [CrossRef] [PubMed]
- Kisielow, J.; Obermair, F.-J.; Kopf, M. Deciphering CD4+ T Cell Specificity Using Novel MHC-TCR Chimeric Receptors. Nat. Immunol. 2019, 20, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Schoemans, H.M.; Lee, S.J.; Ferrara, J.L.; Wolff, D.; Levine, J.E.; Schultz, K.R.; Shaw, B.E.; Flowers, M.E.; Ruutu, T.; Greinix, H.; et al. EBMT−NIH−CIBMTR Task Force Position Statement on Standardized Terminology & Guidance for Graft-versus-Host Disease Assessment. Bone Marrow Transplant. 2018, 53, 1401–1415. [Google Scholar] [CrossRef] [PubMed]
- Prenek, L.; Litvai, T.; Balázs, N.; Kugyelka, R.; Boldizsár, F.; Najbauer, J.; Németh, P.; Berki, T. Regulatory T Cells Are Less Sensitive to Glucocorticoid Hormone Induced Apoptosis than CD4+ T Cells. Apoptosis 2020, 25, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Shah, O.; Tamaresis, J.S.; Kenyon, L.J.; Xu, L.; Zheng, P.; Gupta, P.; Rangarajan, K.; Lee, S.; Spellman, S.; Nikiforow, S.; et al. Analysis of the Whole CDR3 T Cell Receptor Repertoire after Hematopoietic Stem Cell Transplantation in 2 Clinical Cohorts. Biol. Blood Marrow Transplant. 2020, 26, 1050–1070. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.C.; Young, R.; Devine, S.; Hogan, W.J.; Ayuk, F.; Bunworasate, U.; Chanswangphuwana, C.; Efebera, Y.A.; Holler, E.; Litzow, M.; et al. International, Multicenter Standardization of Acute Graft-versus-Host Disease Clinical Data Collection: A Report from the Mount Sinai Acute GVHD International Consortium. Biol. Blood Marrow Transplant. 2016, 22, 4–10. [Google Scholar] [CrossRef]
- Jagasia, M.H.; Greinix, H.T.; Arora, M.; Williams, K.M.; Wolff, D.; Cowen, E.W.; Palmer, J.; Weisdorf, D.; Treister, N.S.; Cheng, G.-S.; et al. National Institutes of Health Consensus Development Project on Criteria for Clinical Trials in Chronic Graft-versus-Host Disease: I. The 2014 Diagnosis and Staging Working Group Report. Biol. Blood Marrow Transplant. 2015, 21, 389–401.e1. [Google Scholar] [CrossRef]
- Shugay, M.; Bagaev, D.V.; Turchaninova, M.A.; Bolotin, D.A.; Britanova, O.V.; Putintseva, E.V.; Pogorelyy, M.V.; Nazarov, V.I.; Zvyagin, I.V.; Kirgizova, V.I.; et al. VDJtools: Unifying Post-Analysis of T Cell Receptor Repertoires. PLoS Comput. Biol. 2015, 11, e1004503. [Google Scholar] [CrossRef]
- Nazarov, V.I.; Pogorelyy, M.V.; Komech, E.A.; Zvyagin, I.V.; Bolotin, D.A.; Shugay, M.; Chudakov, D.M.; Lebedev, Y.B.; Mamedov, I.Z. TcR: An R Package for T Cell Receptor Repertoire Advanced Data Analysis. BMC Bioinform. 2015, 16, 175. [Google Scholar] [CrossRef]
- Venturi, V.; Kedzierska, K.; Turner, S.J.; Doherty, P.C.; Davenport, M.P. Methods for Comparing the Diversity of Samples of the T Cell Receptor Repertoire. J. Immunol. Methods 2007, 321, 182–195. [Google Scholar] [CrossRef]
- Scrucca, L.; Santucci, A.; Aversa, F. Competing Risk Analysis Using R: An Easy Guide for Clinicians. Bone Marrow Transplant. 2007, 40, 381–387. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All | GVHD | noGVHD | p-Value (GVHD vs. noGVHD) | |
|---|---|---|---|---|
| N | 21 | 12 | 9 | |
| Sex, male | 9 (43) | 5 (42) | 4 (44) | >0.999 |
| Age at first DLI, years | 48 (23–72) | 46.5 (36–63) | 51 (23–72) | 0.675 |
| Disease details: AML | 15 (71) | 8 (67) | 7 (78) | 0.659 |
| Disease risk: adverse * | 5 (24) | 2 (17) | 3 (33) | 0.611 |
| Donor age at alloHSCT, years | 31 (22–61) | 31 (22–58) | 29 (22–61) | 0.417 |
| Female donor, male recipient | 2 (10) | 1 (8) | 1 (11) | >0.999 |
| Conditioning regimen: RIC | 14 (67) | 8 (67) | 6 (67) | >0.999 |
| aGVHD post alloHSCT | 8 (38) | 3 (25) | 5 (56) | 0.203 |
| c/oGVHD post alloHSCT | 2 (10) | 2 (17) | 0 | 0.486 |
| Antineoplastic treatment pre DLI | 12 (57) | 7 (58) | 5 (56) | >0.999 |
| First DLI, months post alloHSCT | 10.5 (4.6–75.7) | 12.6 (5.6–26.3) | 9.6 (4.6–75.7) | 0.508 |
| Dose of first DLI, CD3+/kg BW | 0.5 × 107 (0.1 × 107–2.5 × 107) | 0.75 × 107 (0.1 × 107–2.5 × 107) | 0.5 × 107 (0.1 × 107–1 × 107) | 0.811 |
| Total number of DLIs | 2 (1–7) | 2.5 (1–7) | 1 (1–4) | 0.097 |
| Cumulative amount of applied donor cells, CD3+/kg BW | 1 × 107 (0.5 × 107–32.5 × 107) | 4.7 × 107 (0.5 × 107–32.5 × 107) | 1 × 107 (0.5 × 107–5.6 × 107) | 0.022 |
| Antineoplastic treatment post DLI | 12 (57) | 5 (42) | 7 (78) | 0.184 |
| Number of relapse post DLI | 12 (57) | 5 (42) | 7 (78) | 0.184 |
| Relapse post DLI, months | 8.7 (0.2–19.3) | 11.7 (6.1–19.3) | 3.7 (0.2–15) | 0.057 |
| OS at 36 months post DLI, months | 17.8 (1.7–36) | 20.9 (2.6–36) | 16.4 (1.7–36) | 0.260 |
| Patients alive at 36 months post DLI | 6 (29) | 4 (33) | 2 (22) | 0.659 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, J.; Kuhlmann, L.; Xiao, Y.; Raha, S.; Bernhardt, G.; Stadler, M.; Thol, F.; Heuser, M.; Eder, M.; Ganser, A.; et al. Healthy-like CD4+ Regulatory and CD4+ Conventional T-Cell Receptor Repertoires Predict Protection from GVHD Following Donor Lymphocyte Infusion. Int. J. Mol. Sci. 2022, 23, 10914. https://doi.org/10.3390/ijms231810914
Schneider J, Kuhlmann L, Xiao Y, Raha S, Bernhardt G, Stadler M, Thol F, Heuser M, Eder M, Ganser A, et al. Healthy-like CD4+ Regulatory and CD4+ Conventional T-Cell Receptor Repertoires Predict Protection from GVHD Following Donor Lymphocyte Infusion. International Journal of Molecular Sciences. 2022; 23(18):10914. https://doi.org/10.3390/ijms231810914
Chicago/Turabian StyleSchneider, Jessica, Leonie Kuhlmann, Yankai Xiao, Solaiman Raha, Günter Bernhardt, Michael Stadler, Felicitas Thol, Michael Heuser, Matthias Eder, Arnold Ganser, and et al. 2022. "Healthy-like CD4+ Regulatory and CD4+ Conventional T-Cell Receptor Repertoires Predict Protection from GVHD Following Donor Lymphocyte Infusion" International Journal of Molecular Sciences 23, no. 18: 10914. https://doi.org/10.3390/ijms231810914
APA StyleSchneider, J., Kuhlmann, L., Xiao, Y., Raha, S., Bernhardt, G., Stadler, M., Thol, F., Heuser, M., Eder, M., Ganser, A., Ravens, S., Förster, R., Prinz, I., Koenecke, C., & Schultze-Florey, C. R. (2022). Healthy-like CD4+ Regulatory and CD4+ Conventional T-Cell Receptor Repertoires Predict Protection from GVHD Following Donor Lymphocyte Infusion. International Journal of Molecular Sciences, 23(18), 10914. https://doi.org/10.3390/ijms231810914