Abstract
The COVID-19 pandemic has had a deep impact on people worldwide since late 2019 when SARS-CoV-2 was first identified in Wuhan, China. In addition to its effect on public health, it has affected humans in various aspects of life, including social, economic, cultural, and political. It is also true that researchers have made vigorous efforts to overcome COVID-19 throughout the world, but they still have a long way to go. Accordingly, innumerable therapeutics and vaccine candidates have been studied for their efficacies and have been tried clinically in a very short span of time. For example, the versatility of extracellular vesicles, which are membrane-bound particles released from all types of cells, have recently been highlighted in terms of their effectiveness, biocompatibility, and safety in the fight against COVID-19. Thus, here, we tried to explain the use of extracellular vesicles as therapeutics and for the development of vaccines against COVID-19. Along with the mechanisms and a comprehensive background of their application in trapping the coronavirus or controlling the cytokine storm, we also discuss the obstacles to the clinical use of extracellular vesicles and how these could be resolved in the future.
1. Introduction
Coronavirus disease 2019 (COVID-19), caused by severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), a single-stranded, positive-sense RNA virus, was first identified in Wuhan, China, in 2019 and was declared a pandemic by the World Health Organization, with over 572 million confirmed cases and 6.3 million deaths by 25 July 2022 [1]. SARS-CoV-2 enters the host cell through its spike protein by binding to the receptor binding domain (RBD) of the S1 subunit to the angiotensin-converting enzyme 2 (ACE2), a receptor of the host cell surface, thereby initiating infection; this is further accompanied by the S2 subunit with the help of cellular proteases, fusing the virus the cell and successfully releasing the viral genome into the cell [2]. The spike protein of SARS-CoV-2 is the most important target for researchers in the development of therapeutics and vaccines for blocking infection compared with other interacting mediators [3,4]. Symptoms of COVID-19 can be mild, for example, rhinitis, fever, cough, sore throat, and shortness of breath, or they can progress to severe symptoms such as acute respiratory distress syndrome (ARDS), pneumonia, and acute lung injury, where mechanical ventilation is needed [5]. So far, researchers have focused on not only the pathogenicity of the virus itself but also the counter-immune response to it. Extracellular vesicles (EVs) are now considered one of the important entities in fighting against COVID-19 due to their roles in SARS-CoV-2 pathogenesis and their usefulness as novel therapeutics, delivery vehicles for drugs, and vaccine platforms [6,7,8].
EVs, comprising a variety of nano-scale vesicles ranging from 50 to 1000 nm in size, are released from all types of cells carrying a variety of lipids, proteins, and nucleic acids in a more protective manner than un-enveloped circulating biomolecules such as antibodies and cytokines from cellular DNases, RNases, proteases, and other degrading materials, due to the presence of the lipid bilayer membrane. Moreover, they have been shown to be nontoxic and unable to induce immune response, depending on the cell type [9], which is why they have been engineered to express targeting receptors or ligands artificially [10] or as a drug delivery system in cancer [11] and other various inflammatory diseases [12] and viral infections [13]. Similarly, EVs have recently been utilized in numerous ways as novel strategies against COVID-19, including for diagnostic purposes, therapeutics, and vaccine development [14].
In this review, we aim to summarize currently developed or developing EV-based vaccines and therapeutics produced by manipulating extracellular vesicles against COVID-19 to date, as summarized in Table 1. This includes the engineering of EVs, expressing a part of the spike (S), RBD of SARS-CoV-2; full-length S or ACE2 of the host cell for trapping virus; or reducing the cytokine storm depicted in Figure 1. Finally, we will highlight the issues and associated with developing EVs against COVID-19.

Table 1.
EV-based therapeutics and vaccine candidates under development for COVID-19.
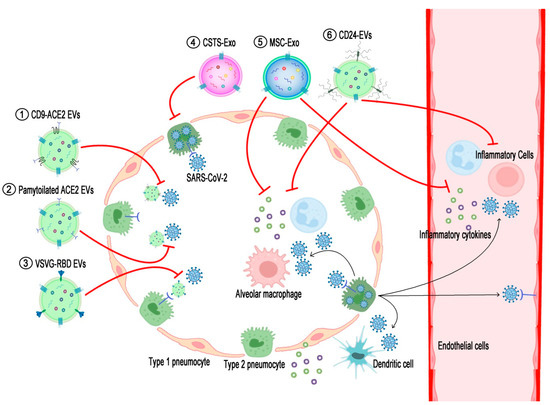
Figure 1.
Schematic diagram displaying action mechanisms of EV-based therapeutics against COVID-19. SARS-CoV-2 enters and proliferates in the type 2 pneumocytes of the lung and then spreads into the interstitial tissue and bloodstream. Components of either viruses or dying cells activate alveolar macrophages and dendritic cells to recruit inflammatory cells to the lung tissue, resulting in the over-secretion of pro-inflammatory cytokines (cytokine storm). (1) EVs expressing ACE2-fused with CD9 (CD9-ACE2 EVs) or (2) palmitoylated ACE2 (palmitoylated ACE2 EVs) prohibit the binding of viruses to cellular ACE2. (3) EV-expressing RBDs of SARS-CoV-2 spike proteins fused with the stem region of the VSVG protein (VSVG-RBD EVs) target ACE2-expressing cells and thereby introduce anti-viral siRNAs to inhibit the proliferation of the viruses. (4) COVID-19-specific T-cell-derived exosomes (CSTS-Exo) show anti-viral effects on virus-infected cells by their cargo such as IFNγ. (5) EVs from mesenchymal stem cells (MSC-Exo) or (6) EVs expressing CD24 (CD24-EVs) can ameliorate the cytokine storm induced by over-activated inflammatory cells in the severe phase of COVID-19. The black lines briefly indicate the pathogenic pathways of SARS-CoV-2 infection in the airway, and the red lines denote the action mechanisms of the EV-based therapeutics interrupting the COVID-19 pathogenesis.
Extracellular Vesicles
Extracellular vesicles, which are surrounded by a membranous lipid bilayer, are released into the extracellular space by all types of cells for the transport of cellular cargo to regulate a variety of biological processes, subsequently playing an essential role in intercellular communication [32,33].
EVs can be generated through one of two pathways: the endosomal pathway, which consists of the internalization of membrane proteins through the endocytic pathway being sequentially maturated into early endosomes, late endosomes, and multivesicular bodies, and the non-endosomal pathway, which refers to the externalization of membrane enclosing internal cellular cargos and membrane proteins in the way to first form buddings and then pinch off to form vesicles [34]. Even though EVs are secreted by both prokaryotic and eukaryotic cells, there has been little research so far on the secretion from prokaryotes; however, extensive studies have been performed on the release of EVs from mammalian cells such as neuronal cells [35], endothelial cells [36], mesenchymal stem cells (MSCs) [37], and epithelial cells [38]. They are found in many biological fluids, i.e., blood [39], synovial fluid [40], milk [41], urine [42], and saliva [43]. Although during the process of synthesis and secretion they share similar markers that make their classification difficult, on the basis of size, degree of similarity, and biogenetic mechanism, there are three major types of extracellular vesicles, i.e., exosomes, microvesicles, and apoptotic bodies [34]. However, the term EVs is used for exosomes and microvesicles in this review, as recommended by the International Society of Extracellular Vesicles (ISEV) guidelines, because of the heterogeneous nature of their preparation [44], unless otherwise specified in the study
Exosomes are characterized as the smallest EVs, ranging from 40 to 150 nm in diameter and originating from the endocytosis of the cellular membrane, resulting in the formation of multivesicular bodies (MVBs) by the inward invagination of the late endosomal membrane with the role of endosomal sorting complex required for transport (ESCRT) proteins [45]. Currently, exosomes can be purified using several methods, such as differential ultracentrifugation, rate-zonal centrifugation, ultrafiltration, poly-ethylene-glycol (PEG)-based precipitation, immunoaffinity capture, microfluidics, and size-exclusion capture [46]. Most popularly, exosomes are isolated through ultracentrifugation at 100,000× g after apoptotic bodies and microvesicles are removed, which are comparatively larger in size, and if further purification is needed, rate-zonal centrifugation is performed using the Optiprep gradient [47]. Exosomes express specific proteins, for example, Alix, TSG101, and clathrin, involved in the endocytosis of the plasma membrane and MVB formation. Similarly they express RAB proteins and annexins involved in membrane trafficking, and also tetraspanins, i.e., CD63, CD9, and CD81. The enrichment of these proteins make exosomes easily distinguishable for researchers from other non-classical exosomes that do not express exosomal markers such as CD63, CD9, and CD81 [48].
Microvesicles are defined as large vesicles [44] formed by the outward protrusion of the plasma membrane and range from 100 to 1000 nm in size but on average are 250–400 nm in diameter [49,50]. The pinching off and detachment of the membrane occurs at specific places and is influenced by the distribution of phospholipids along with the phosphorylation of myosin mediated by Rho-kinase along with contractile machinery [51,52], where they enclose nearby biomolecules, cell surface proteins, and fragmented rRNA and mRNA, which are planned to be trafficked towards the plasma membrane [53,54]. Microvesicles are composed of lipids displaying plasma membrane receptors and molecules of the cells from which they originated. They are smaller than apoptotic bodies and can be isolated from biofluids and cellular supernatants by centrifugation at 10,000× g after removing cell debris containing apoptotic bodies [34].
Apoptotic bodies, the largest among EVs, range from 1 to 5 um in diameter and are generated during the programmed self-destruction of the cell, and they can be pelleted down at speeds of 2000–4500 g; they share markers with microvesicles and exosomes and hence are distinguished from other vesicles based on size [44]. They originate from the protrusion and blebbing of the membranes of dying cells that contain intact cellular organelles and genomic DNAs, damaged nucleic acids, and randomly packaged cargo [55]. Cell death can be induced during normal physiological pathways or through any pathological pathways that initiate from the blebbing of the membrane, resulting in the formation of apoptotic projections such as microtubular spikes [56], apoptopodia [57], and beaded apoptopodia [55]. Apoptotic cells release “find-me” signals to recruit phagocytes and “eat-me” signals such as phosphatidylserine (PS) and exposed outside apoptotic bodies that bind to receptors in phagocytic cells, mediating the engulfment of apoptotic bodies and finally resulting in the clearance of apoptotic cells by phagocytes [58,59,60].
2. Roles of Extracellular Vesicles in COVID-19 Pathogenesis
2.1. Pathogenesis of COVID-19
The disease progression of COVID-19 is represented by three phases, depending on the clinical status: the viremia phase, the acute phase (pneumonia phase), and the recovery phase [61,62]. During the viremia phase, early infection by the virus is initiated in the epithelial cells of the nasal cavity and the larynx, the major site of virus entry, through angiotensin-converting enzyme 2(ACE2) receptors with the help of transmembrane serine protease 2 (TMPRSS2) [63,64].
Mature SARS-CoV-2 enters the alveolar epithelial cells after replicating itself in upper respiratory tracts using host cell machinery such as RNA polymerase, ribosomes, and cellular enzymes to synthesize its structural proteins such as spike, envelope, membrane, and nucleocapsid (S, E, M, and N) and other accessory proteins [2]. The virus divides itself into high numbers and enters the bloodstream from the lungs, resulting in viremia, which again attacks various organs including the lungs, kidneys, and gastrointestinal tracts [65]. In the acute phase (pneumonia phase), pattern-recognition receptors (PRRs) present in the immune cells are attracted to lung tissues, recognize danger-associated molecular patterns (DAMPs) from the destructed tissues and pathogen-associated molecular patterns (PAMPs) from the virus, and produce a variety of pro-inflammatory mediators such as cytokines, chemokines, and inflammatory mediators that are necessary for adaptive immune responses against SARS-CoV-2. In addition, in lymphopenia, a significant reduction in the number of lymphocytes is observed in patients in this phase [66].
Most patients progress to a convalescent phase by acquiring adaptive immunity to SARS-CoV-2, along with a decline in virus titers and a recovery of blood lymphocytes [67]. A small portion of patients from the pneumonia phase develop a severe phase. Most patients who progress into the severe phase are either immune-compromised or old in age and suffering from underlying chronic diseases such as diabetes, hypertension, respiratory diseases, and cancer [68]. In the severe phase, lymphopenia is worsened, and the increase in serum pro-inflammatory mediators is sustained [69]. An increase in cytokines and chemokines attracts other inflammatory cells such as neutrophils and monocytes to the lung tissues, leading to an influx in inflammatory cells that results in an increase in pathogenic inflammatory cells [70], finally causing a cytokine storm [71,72]. Pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukins 1 and 6 (IL-1 and IL-6) increase permeability in the host’s vascular system by the dilation of smooth muscle and the contraction of the endothelial cells of blood vessels, which leads to alveolar edema and alveolar collapse, thereby leading to refractory hypoxemia and finally ARDS. Thus, the exchange of gases in the lungs is impaired, thereby increasing respiratory and heart rate to compensate for the oxygen deficiency, which makes breathing extremely difficult for the patient [71,72,73]. At the same time, an excess of circulating cytokines causes other tissues to induce a systemic inflammatory response. Moreover, blood pressure is significantly reduced due to the vasodilation of the organs and tissues, resulting in multisystem organ failure [71]. In addition to ARDS, disseminated intravascular coagulation is a common critical factor in multisystem organ failure [74].
2.2. EVs in COVID-19 Pathogenesis
It is highly interesting to consider the developing theory that EVs contribute to the dissemination and persistence of genetic material and proteins of SARS-CoV-2 [75] due to the similarity in the entrance, budding, and mechanisms of biogenesis during infection. Previous research has already shown that EVs enhance infections caused by CMV, HIV-1, or HSV-1 by transferring viral proteins and genetic material from infected cells to healthy cells [76,77,78]. Tetraspanins such as CD9 are one of the most widely expressed proteins on the surface of EVs and are thought to work together with TMPRSS2 to cleave fused viral glycoproteins and speed up the entrance of Middle East respiratory syndrome coronavirus (MERS-CoV) into lung cells [79]. Additionally, CD9 enhances lentiviral infection and improves transduction efficiency in immune-competent cells such as T lymphocytes and B cells [80]. In SARS-CoV-2 infection, EVs expressing ACE2 on their surface are suggested to be responsible for spreading and accelerating infection by assisting viral entry into cells [81]. EVs from the plasma of COVID-19 patients incorporate SARS-CoV-2 spike-derived fragments of proteins and RNA, albeit in a low copy number, but are able to progress the disease and induce immune responses [6,82,83]. In other respects, ACE2 on EVs prevents SARS-CoV-2 infection through its function as a decoy receptor [16,17]. Given that EVs have some common physical and biogenic characteristics with viruses, EVs derived from convalescent patient serum also express spike protein on their surface and act as a decoy target for neutralizing antibodies by their competitive inhibition of binding between the antibodies and the mature virion [82].
3. EV-Based Therapeutics against SARS-CoV-2
The method of communication of EVs from cell to cell is by transferring intraluminal EV cargo such as proteins, lipid, mRNA, and miRNA, under normal physical conditions and also in diseased states, which makes EVs very interesting. EVs have hence been exploited by researchers as a therapeutic modality for delivering substances of interest (15). EVs on one hand have been engineered to evoke the immune response in terms of immunotherapies [84], while on other hand, they have been utilized as delivery drugs and immune modulators to treat various human diseases such as solid and hematologic cancers, autoimmune diseases, and infectious diseases [85,86]; now EVs have been used as novel therapeutics against COVID-19, using multiple approaches to blocking infection to healthy cells [16,87].
3.1. Extracellular Vesicles Tagged with RBD
The very first target of the SARS-CoV-2 virus is type 2 alveolar epithelial cells of the lungs, where it binds to the highly expressing ACE2 receptors [88]. Although EVs have a great capacity as a natural carrier to deliver cargos to the recipient cells, a majority of externally injected EVs are absorbed by the liver, spleen, and pancreas instead of being taken up by the specific target. Therefore, for the satisfactory localization of specifically targeted therapeutic particles, it is suitable to tag EVs with tissue-specific peptides or antibodies targeting specific antigens [89].
In previous studies, vesicular stomatitis virus-G protein (VSVG) has been used as a fusion backbone with a variety of reporter proteins that include luciferase, green fluorescent protein (GFP), and red fluorescent protein (RFP) for the tracking and detection of EVs [90]. Recently, VSVG has been engineered to be fused with RBD of SARS-CoV-2 virus protein, the key domain in virus attachment and entry, by replacing the ectodomain of VSVG with RBD, resulting in the production of pseudoviral particles expressing RBD–VSVG fusion protein, without changing the physical properties of the modified EVs [15]. This is because it has been found that EVs and VSVs have similar lipid envelope compositions resulting from shared intracellular trafficking [91].
The association between the RBD of SARS-CoV-2 and the ACE2 of the host has been thought to be the most crucial to be targeted for vaccine development [92], antibody neutralization, and small-molecule inhibitors [93], which block the entry of virus into the cell. The host cell receptor ACE2 is present in most cell types, including the heart, intestine, kidneys, and lungs, among which the lungs show the highest expression of ACE2s and are proven to have a stronger affinity for SARS-CoV-2 than other organs [88,94,95,96]. Similarly, it has been found that the cellular presence of ACE2 receptors was needed for the entry of RBD-tagged EVs, which is why they were successfully targeted to the lungs and other tissues expressing ACE2 and reduced the infection of SARS-CoV-2 by delivering siRNA incorporated into RBD-tagged EVs, which was conducted in a transgenic hACE2 mouse model. This study suggested that SARS-CoV-2 infection could be efficiently treated by the delivery of antiviral drugs and that RBD-tagged EVs could be a potential therapeutic approach for other diseases [15].
3.2. EVs Expressing Tetraspanins Fusion
Tetraspanins, CD9, CD63, CD81, CD82, and CD151, are transmembrane proteins concentrated in the plasma membrane lipid microdomain and largely expressed in the exosomes, which makes them exosomal markers. They have also been known to contribute to the biogenesis and cargo sorting of exosomes [97]. In the field of COVID-19 therapeutics, engineered EVs expressing a novel fusion of a tetraspanin, i.e., CD63 with anti-SARS-CoV-2 nanobody, inhibited the binding of SARS-CoV-2 to ACE2, thereby neutralizing the SARS-CoV-2 infection [4]. Similarly, EVs expressing soluble ACE2 on their surface by the fusion of truncated scaffold of CD9 serve as decoy receptors for SARS-CoV-2 and block infection by SARS-CoV-2 variants including D614G, δ, and β, as well as the wild type [3]. Consistent with the current data, EVs expressing ACE2 inhibited the infection of pseudotyped lentiviruses from variants of concern of SARS-CoV-2, which was even more enhanced along with TMPRSS2 [16].
In addition to engineered EVs expressing specific proteins for hindering the cellular entrance of SARS-CoV-2, EVs produced from cell lines such as Vero CCL-81 and Vero E6 infected with SARS-CoV-2 can display surface proteins of SARS-CoV-2 that can recognize alveolar macrophages. Therefore, these EVs can be used as vehicles encapsulating drug delivery platforms [98,99].
3.3. ACE2 Loading onto EVs to Block Virus Entry
Because of the quickly evolving variants of SARS-CoV-2, COVID-19 remains a matter of concern and a major challenge despite tremendous advances in vaccine development and novel therapeutics [100,101,102]. Therefore, it is crucial to develop a strategy to cope with newly emerging variants of SARS-CoV-2. In this critical situation, EVs expressing ACE2 (evACE2) could be an alternative strategy. Supporting these data, small EVs, as well as non-membranous extracellular nanoparticles, exomeres, are reported to express ACE2 on their surface which subsequently inhibit SARS-CoV-2 infection, acting as a decoy by binding to the virus [103].
Moreover, the loading of ACE2 onto EVs is highly dependent on ACE2 palmitoylation at two major sites, i.e., Cys141 and Cys498, called S-palmitoylation. EVs’ secretion of ACE2 from the plasma membrane is increased by palmitoylation through the zinc finger DHHC-Type palmitoyltransferase 3 (ZDHHC3) or decreased by de-palmitoylation through acyl protein thioesterase 1 (LYPLA1). During viral infection, ACE2 present on the membrane is secreted along with EVs, and it blocks infection with SARS-CoV-2 by binding to the RBD domain, preventing the binding of SARS-CoV-2 with cellular ACE2. Therefore, engineered EVs enriched with ACE2 on their surface by palmitoylation were bound efficiently to RBDs and subsequently neutralized both pseudo-typed and wild-type SARS-CoV-2 in human ACE2 transgenic mice [18].
3.4. CD24-Loaded EVs
CD24, expressed in a variety of cells, is a glycosylphosphatidylinositol (GPI)-anchored glycoprotein and is localized in lipid microdomain [104,105]; its interaction with Siglecs, which are mostly expressed by cells of the immune system, has been shown to transduce inhibitory signals on Siglec-expressing inflammatory cells [106,107,108]. Therefore, CD24 fused with Fc fragment of Ig (CD24-Fc) was tested to evaluate its effect in reducing over-activated inflammation in COVID-19, which is similar to human immunodeficiency virus type-1/simian immunodeficiency virus infection, where it provides protection to Chinese rhesus macaques (ChRMs) against disease progression [109,110]. Intriguingly, CD24 is also highly expressed in EVs, possibly due to its localization in lipid microdomain and GPI anchorage, prompting the development of CD24-expressing EVs as a novel therapeutic. Consequently, exosomes derived from CD24-overexpressing 293T cells, T-Rex™ (EXO-CD24), are developed for the treatment of COVID-19 cytokine storms with the format of inhalation. The rationale of this new drug is that exosomes expressing CD24 can attenuate the cytokine storm via activating anti-inflammatory immune cells through CD24 signaling by over-activating innate immune responses of SARS-CoV-2 patients [19].
3.5. EVs from Convalescents
Exosomes from the plasma of the convalescent phase of COVID-19 patients were recently reported to be equipped with all the components, including viral proteins, peptides, and RNAs for successful adaptive immune responses to SARS-CoV-2 [83].
The release of EVs expressing ACE2 (evACE2) was markedly increased in the convalescent serums of severely infected COVID-19 patients. They inhibited infections of SARS-CoV-2 including α, β, and δ strains with 135 times higher efficiency than recombinant human ACE2 (rhACE2), by competing with cellular ACE2 in the binding with RBD of SARS-CoV-2. In vivo, the ACE2-transgenic mouse model evACE2 was 60 to 80 times more potent than rhACE in inhibiting infection from both original and pseudotyped viruses and also protected mice from SARS-CoV-2-induced lung injury [17].
Following on this finding, virus-specific T cells should have similar capabilities with the plasma exosomes, and they are currently in clinical trials. COVID-19-specific T-cell-derived exosomes (CSTC-Exo) were purified from T cells of convalescent COVID-19 patients and expanded in vitro by stimulation with SARS-CoV-2-specific peptides and cytokines. An inhalable format of CSTC-Exo is now in clinical trials in COVID-19 patients with early pneumonia. Action mechanisms of CSTC-Exo have suggested that cytokines, including IFN-γ within the exosomes prepared from activated SARS-CoV-2-specific T cells, have anti-viral effects on COVID-19 patients [20].
Convalescent plasma infusion has been successfully tried in severe COVID-19 patients for treatment [111,112] and is now considered a possible treatment choice in severe COVID-19 patients not responding to existing therapies [113]. Among the components of convalescent plasma, it has been proposed that EVs from platelets are the major constituents responsible for the regeneration of damaged tissues [114]. Therefore, platelet-derived EVs could be used as a COVID-19 therapeutic. Supporting this notion, platelet-derived exosomes loaded with an anti-inflammatory drug, [5-(p-fluorophenyl)-2-ureido] thiophene-3-carboxamide (TPCA-1), have been shown to alleviate cytokine storm in pneumonia in a mice model [21].
4. EV-Based Vaccines against SARS-CoV-2
EVs have also been used as a novel platform of vaccines by manipulating them in numerous interesting ways, although lipid nanoparticle-based vaccines have successfully been used as delivery vehicles for SARS-CoV-2 mRNA. The lipid nanoparticle-based vaccine reportedly has some major shortcomings such as temperature instability, thus requiring the maintenance of ultra-low temperatures, as well as side effects and adverse reactions, and poor mucosal immunity [115,116,117]. Thus, EV-based vaccines can be proposed to overcome the drawbacks of lipid nanoparticle-based mRNA vaccines. Here, we want to introduce the achievements with significant advances among various projects recently forwarded.
Exosomes loaded with an mRNA-encoding spike and nucleocapsid proteins of SARS-CoV-2 induced long-lasting cellular and humoral immune responses in a mouse model and proved to be safer than other lipid nanoparticle-based mRNA vaccines [27].
Moreover, EVs purified from cells transfected with a SARS-CoV-2 spike DNA (CoVEXax™, Ciloa SAS) triggered antibody- and cell-mediated immunities in mice without using any adjuvants [28].
A candidate vaccine that was based on the lyophilized exosomes derived from lungs (S-Exos) expressing mRNA of SARS-CoV-2 spike protein [29] and exosomes conjugated with recombinant SARS-CoV-2 RBD (RBD-Exo) [30] was more potent in stimulating IgG and secretory IgA (SIgA) responses when delivered to rodents and nonhuman primates compared with synthetic nanoparticle liposomes loaded with the spike mRNA of SARS-CoV-2. These EV-based vaccines were stable, even at room temperature, for more than a month when formulated as either inhalable dry powder or in lyophilized form. They could also be directly distributed to the bronchioles and parenchyma of the lungs [29,30].
An intranasal vaccine based on bacterial EVs and outer membrane vesicles (OMV) of Salmonella typhimurium displaying RBD of SARS-CoV-2 produced high titers of neutralizing antibodies against Wuhan-type SARS-CoV-2, as well as delta variants, and protected immunized golden Syrian hamster from infection with SARS-CoV-2 with no loss of body mass and fewer viral titers in bronchoalveolar lavage fluids compared with the control [31]. Hence, it is believed that EV-based vaccines are ready to circumvent the current limitations of lipid nanoparticle-based mRNA vaccines.
5. Mesenchymal Stem Cell (MSC)-Derived EVs in COVID-19
MSCs are adult stem cells derived from multiple tissues such as umbilical cord, bone marrow, adipose tissue, and amniotic fluid and have the ability to regenerate and differentiate into cartilage, bone, nerve, muscle, and skin cells [118]. They are considered to be the most useful tool in regenerative medicine as they can repair damaged tissue and organs and release a variety of cytokines, chemokines, growth factors, and also EVs [119]. Compared with other cell types, MSCs are known to release tremendously high amounts of EVs, including exosomes. EVs from MSCs have been reported to have similar therapeutic effects to MSCs because they have active biomolecules analogous to MSCs and because EVs from MSCs are responsible for the paracrine effect of MSCs in MSC cell therapies [120]. Researchers have been testing stem-cell-derived EVs for the management of COVID-19, and the majority of these EVs are in clinical trials as summarized by Krishan et al. [121].
One of the therapeutically active parts of MSC exosomes is miRNAs inside exosomes. Thus, MSC exosomes containing miRNAs have been used as cell-free therapeutics in multiple diseases, such that miRNA in exosomes from umbilical cord-derived MSCs (UC-MSCs-Exo) inhibited hepatitis C virus (HCV) infection by silencing viral RNA in combination with other FDA-approved drugs [122]. Similarly, COVID-19 patients with mild pneumonia showed improved clinical outcomes when EVs were directly targeted to the lungs via inhalation by the nebulization of UC-MSCs-Exo and human adipose-derived exosomes (haMSC-Exos) without causing any side effects [22,23]. Owing to the regenerative and repair nature of MSC EVs as proved through in silico studies, miRNA from MSC EVs attenuated the cytokine storm, protected cells from damage, and blocked the activation of the coagulation pathway caused by COVID-19. Hence, these results show the effects of MSC EVs on multiple targets in COVID-19 [123].
Another promising therapeutic candidate based on exosomes derived from MSCs of allogeneic bone marrow (ExoFlo™, Direct Biologics, Austin, TX, USA) is currently being studied in human clinical trials. As a result, it has been proposed that it is very effective in moderate to severe COVID-19 patients. Given that MSCs-derived EVs are safer compared to MSCs because of their non-replicative, non-tumorigenic, and non-immunogenic nature, ExoFlo™ was quite useful in restoring plasma oxygen contents, lowering cytokine storm, and reconstituting immunity in infected patients during human trials [24].
MSC-Exo in COVID-19 patients may induce the differentiation of M2 macrophages by promoting the secretion of PEG2 and accordingly can inhibit inflammation by blocking the release of pro-inflammatory cytokines and can reduce the effects of pro-inflammatory cytokines through anti-inflammatory cytokines such as IL10. Similarly, MSC-Exo can regenerate and repair the damaged tissues through the induction of growth factors such as vascular endothelial growth factor (VEGF), keratinocyte growth factor (KGF), and hepatocyte growth factor (HGF), as already reported in multiple disease models [124,125,126]. Moreover, EVs derived from Wharton’s jelly MSC could be very effective in reducing nuclear the NF-kB-mediated cytokine storm in COVID-19 patients with diabetes mellitus or renal disease [26].
Other studies about EVs derived from amniotic fluid named Zofin™ have proved them to be successful in improving the conditions of three critical patients suffering from COVID-19 without showing any adverse effects or safety concerns [25,127].
6. Conclusions
In this review, we aimed to describe currently developing therapeutics and vaccine candidates based on EVs for COVID-19, together with their action mechanisms and comprehensive backgrounds for their application, as summarized in Table 1 and Figure 1. EVs have been an ideal natural therapeutic platform serving as a decoy or as a delivery vehicle carrying drugs and other cargo and thus communicating within or between cells in all organisms, from bacteria to humans.
However, it is also true that there are several clear issues that need to be overcome for the clinical use of EVs to be possible. Most researchers in the field of EVs are pointing out common problems in the commercialization of EVs: extremely low yields of EVs from the large-scale production of conditioned media of cultured cells; isolation of EVs and the challenges in each method of EV isolation; discrepancies between produced batches of EVs from different cell types; maintenance of efficacy during storage, transportation, and biodistribution of EVs, including targeting EVs to the desired tissues or organs without loss of efficacy; and also the issue of long-term endurance by the human body during clinical studies despite having better safety compared to cell therapies.
Considering these factors, we propose that most of the problems come from our incomplete understanding of the extreme heterogeneity of EVs. Our knowledge of the biogenesis of EVs in individual cells appears not to be sufficient despite significant advances in recent research.
Moreover, specific cells release their unique EVs with peculiar cargos using different mechanisms of synthesis depending on the tissues of origin and the cell condition. We are aware that stressed cells can release different kinds of EVs from those of normal cells, depending on the kind of stress.
Furthermore, the cell lines being used for the isolation of EVs for clinical purposes are limited to several non-cancerous cell lines and primary human cells because of their origin and biocompatibility issues. Therefore, a more precise understanding of the mechanisms of action of EVs, along with their biogenesis, is required for the commercialized usage of EVs to combat COVID-19 and other emerging diseases.
Author Contributions
Conceptualization, writing—original draft preparation, writing—review and editing: T.M. and Y.-J.C.; writing—original draft preparation and review: M.S.K.; compilation and organization of most of the information in tables: T.M.; generation of the figure: Y.-J.C.; writing—review and editing; D.A.C., D.W.K., J.K., K.R.H., Y.H., S.L. and D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant, funded by the Korean government (MSIT: Ministry of Science, ICR & Future Planning) (No. 2022R1A2C2003413); by a grant from the Korean Health Technology R&D Project through the Korean Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number: HV22C0073); and by a Korea Basic Science Institute (National Research Facilities and Equipment Center) grant, funded by the Ministry of Education (grant No. 2019R1A6C1010003).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they do not have a conflict of interest.
References
- WHO. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 1 August 2022).
- Huang, Y.; Yang, C.; Xu, X.; Xu, W.; Liu, S. Structural and Functional Properties of SARS-CoV-2 Spike Protein: Potential Antivirus Drug Development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Kim, H.K.; Cho, J.; Kim, E.; Kim, J.; Yang, J.-S.; Kim, K.-C.; Lee, J.-Y.; Shin, Y.; Palomera, L.F.; Park, J.; et al. Engineered Small Extracellular Vesicles Displaying ACE2 Variants on the Surface Protect against SARS-CoV-2 Infection. J. Extracell. Vesicles 2022, 11, e12179. [Google Scholar] [CrossRef]
- Scott, T.A.; Supramaniam, A.; Idris, A.; Cardoso, A.A.; Shrivastava, S.; Kelly, G.; Grepo, N.A.; Soemardy, C.; Ray, R.M.; McMillan, N.A.J.; et al. Engineered Extracellular Vesicles Directed to the Spike Protein Inhibit SARS-CoV-2. Mol. Ther.-Methods Clin. Dev. 2022, 24, 355–366. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72 314 Cases From the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Yim, K.H.W.; Borgoni, S.; Chahwan, R. Serum Extracellular Vesicles Profiling Is Associated with COVID-19 Progression and Immune Responses. J. Extracell. Biol. 2022, 1, e37. [Google Scholar] [CrossRef]
- Murphy, D.E.; de Jong, O.G.; Brouwer, M.; Wood, M.J.; Lavieu, G.; Schiffelers, R.M.; Vader, P. Extracellular Vesicle-Based Therapeutics: Natural versus Engineered Targeting and Trafficking. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef]
- Santos, P.; Almeida, F. Exosome-Based Vaccines: History, Current State, and Clinical Trials. Front. Immunol. 2021, 12, 711565. [Google Scholar] [CrossRef]
- Zhu, X.; Badawi, M.; Pomeroy, S.; Sutaria, D.S.; Xie, Z.; Baek, A.; Jiang, J.; Elgamal, O.A.; Mo, X.; Perle, K.L.; et al. Comprehensive Toxicity and Immunogenicity Studies Reveal Minimal Effects in Mice Following Sustained Dosing of Extracellular Vesicles Derived from HEK293T Cells. J. Extracell. Vesicles 2017, 6, 1324730. [Google Scholar] [CrossRef]
- Dang, X.T.T.; Kavishka, J.M.; Zhang, D.X.; Pirisinu, M.; Le, M.T.N. Extracellular Vesicles as an Efficient and Versatile System for Drug Delivery. Cells 2020, 9, 2191. [Google Scholar] [CrossRef]
- Tian, Y.; Li, S.; Song, J.; Ji, T.; Zhu, M.; Anderson, G.J.; Wei, J.; Nie, G. A Doxorubicin Delivery Platform Using Engineered Natural Membrane Vesicle Exosomes for Targeted Tumor Therapy. Biomaterials 2014, 35, 2383–2390. [Google Scholar] [CrossRef]
- Iyer, S.S.; Rojas, M. Anti-Inflammatory Effects of Mesenchymal Stem Cells: Novel Concept for Future Therapies. Expert Opin. Biol. Ther. 2008, 8, 569–581. [Google Scholar] [CrossRef]
- Zou, X.; Yuan, M.; Zhang, T.; Wei, H.; Xu, S.; Jiang, N.; Zheng, N.; Wu, Z. Extracellular Vesicles Expressing a Single-Chain Variable Fragment of an HIV-1 Specific Antibody Selectively Target Env + Tissues. Theranostics 2019, 9, 5657–5671. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, W.; Wang, Y.; Guo, Q.; Zhao, F.; Zhu, Z.; Xing, Y.; Zhang, H.; Aljofan, M.; Jarrahi, A.M.; et al. The Potential Role of Extracellular Vesicles in COVID-19 Treatment: Opportunity and Challenge. Front. Mol. Biosci. 2021, 8, 699929. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Xiong, S. Tagged Extracellular Vesicles with the RBD of the Viral Spike Protein for Delivery of Antiviral Agents against SARS-CoV-2 Infection. J. Control. Release 2021, 335, 584–595. [Google Scholar] [CrossRef]
- Cocozza, F.; Névo, N.; Piovesana, E.; Lahaye, X.; Buchrieser, J.; Schwartz, O.; Manel, N.; Tkach, M.; Théry, C.; Martin-Jaular, L. Extracellular Vesicles Containing ACE2 Efficiently Prevent Infection by SARS-CoV-2 Spike Protein-Containing Virus. J. Extracell. Vesicles 2020, 10, e12050. [Google Scholar] [CrossRef]
- El-Shennawy, L.; Hoffmann, A.D.; Dashzeveg, N.K.; McAndrews, K.M.; Mehl, P.J.; Cornish, D.; Yu, Z.; Tokars, V.L.; Nicolaescu, V.; Tomatsidou, A.; et al. Circulating ACE2-Expressing Extracellular Vesicles Block Broad Strains of SARS-CoV-2. Nat. Commun. 2022, 13, 405. [Google Scholar] [CrossRef]
- Xie, F.; Su, P.; Pan, T.; Zhou, X.; Li, H.; Huang, H.; Wang, A.; Wang, F.; Huang, J.; Yan, H.; et al. Engineering Extracellular Vesicles Enriched with Palmitoylated ACE2 as COVID-19 Therapy. Adv. Mater. 2021, 33, 2103471. [Google Scholar] [CrossRef]
- Shapira, S.; Ben Shimon, M.; Hay-Levi, M.; Shenberg, G.; Choshen, G.; Bannon, L.; Tepper, M.; Kazanov, D.; Seni, J.; Lev-Ari, S.; et al. A Novel Platform for Attenuating Immune Hyperactivity Using EXO-CD24 in COVID-19 and Beyond. EMBO Mol. Med. 2022, 14, e15997. [Google Scholar] [CrossRef]
- Cetin, M. Aerosol Inhalation of the Exosomes Derived from Allogenic COVID-19 T Cell in the Treatment of Early Stage Novel Coronavirus Pneumonia. Available online: https://clinicaltrials.gov/ct2/show/NCT04389385. (accessed on 29 August 2022).
- Ma, Q.; Fan, Q.; Xu, J.; Bai, J.; Han, X.; Dong, Z.; Zhou, X.; Liu, Z.; Gu, Z.; Wang, C. Calming Cytokine Storm in Pneumonia by Targeted Delivery of TPCA-1 Using Platelet-Derived Extracellular Vesicles. Matter 2020, 3, 287–301. [Google Scholar] [CrossRef]
- Chu, M.; Wang, H.; Bian, L.; Huang, J.; Wu, D.; Zhang, R.; Fei, F.; Chen, Y.; Xia, J. Nebulization Therapy with Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes for COVID-19 Pneumonia. Stem Cell Rev. Rep. 2022. [Google Scholar] [CrossRef]
- Zhu, Y.-G.; Shi, M.-M.; Monsel, A.; Dai, C.-X.; Dong, X.; Shen, H.; Li, S.-K.; Chang, J.; Xu, C.-L.; Li, P.; et al. Nebulized Exosomes Derived from Allogenic Adipose Tissue Mesenchymal Stromal Cells in Patients with Severe COVID-19: A Pilot Study. Stem Cell Res. Ther. 2022, 13, 220. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19. Stem Cells Dev. 2020, 29, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Mitrani, M.I.; Bellio, M.A.; Meglin, A.; Khan, A.; Xu, X.; Haskell, G.; Arango, A.; Shapiro, G.C. Treatment of a COVID-19 Long Hauler with an Amniotic Fluid-Derived Extracellular Vesicle Biologic. Respir. Med. Case Rep. 2021, 34, 101502. [Google Scholar] [CrossRef] [PubMed]
- Khanh, V.C.; Fukushige, M.; Chang, Y.H.; Hoang, N.N.; Yamashita, T.; Obata-Yasuoka, M.; Hamada, H.; Osaka, M.; Hiramatsu, Y.; Ohneda, O. Wharton’s Jelly Mesenchymal Stem Cell-Derived Extracellular Vesicles Reduce SARS-CoV2-Induced Inflammatory Cytokines Under High Glucose and Uremic Toxin Conditions. Stem Cells Dev. 2021, 30, 758–772. [Google Scholar] [CrossRef]
- Tsai, S.J.; Atai, N.A.; Cacciottolo, M.; Nice, J.; Salehi, A.; Guo, C.; Sedgwick, A.; Kanagavelu, S.; Gould, S.J. Exosome-Mediated MRNA Delivery In Vivo Is Safe and Can Be Used to Induce SARS-CoV-2 Immunity. J. Biol. Chem. 2021, 297, 101266. [Google Scholar] [CrossRef] [PubMed]
- Polak, K.; Greze, N.; Lachat, M.; Merle, D.; Chiumento, S.; Bertrand-Gaday, C.; Trentin, B.; Mamoun, R.Z. Extracellular Vesicle-Based Vaccine Platform Displaying Native Viral Envelope Proteins Elicits a Robust Anti-SARS-CoV-2 Response in Mice 2020. bioRxiv 2020. [Google Scholar] [CrossRef]
- Popowski, K.D.; Moatti, A.; Scull, G.; Silkstone, D.; Lutz, H.; López de Juan Abad, B.; George, A.; Belcher, E.; Zhu, D.; Mei, X.; et al. Inhalable Dry Powder MRNA Vaccines Based on Extracellular Vesicles. Matter 2022, 5, 2960–2974. [Google Scholar] [CrossRef]
- Wang, Z.; Popowski, K.D.; Zhu, D.; de Juan Abad, B.L.; Wang, X.; Liu, M.; Lutz, H.; De Naeyer, N.; DeMarco, C.T.; Denny, T.N.; et al. Exosomes Decorated with a Recombinant SARS-CoV-2 Receptor-Binding Domain as an Inhalable COVID-19 Vaccine. Nat. Biomed. Eng. 2022, 6, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Driedonks, T.A.P.; Jong, W.S.P.; Dhakal, S.; Bart van den Berg van Saparoea, H.; Sitaras, I.; Zhou, R.; Caputo, C.; Littlefield, K.; Lowman, M.; et al. A Bacterial Extracellular Vesicle-Based Intranasal Vaccine against SARS-CoV-2 Protects against Disease and Elicits Neutralizing Antibodies to Wild-Type and Delta Variants. J. Extracell. Vesicles 2022, 11, e12192. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Skog, J.; Würdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Sena-Esteves, M.; Curry, W.T.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma Microvesicles Transport RNA and Proteins That Promote Tumour Growth and Provide Diagnostic Biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef]
- Cheng, L.; Hill, A.F. Therapeutically Harnessing Extracellular Vesicles. Nat. Rev. Drug Discov. 2022, 21, 379–399. [Google Scholar] [CrossRef]
- Laulagnier, K.; Javalet, C.; Hemming, F.J.; Chivet, M.; Lachenal, G.; Blot, B.; Chatellard, C.; Sadoul, R. Amyloid Precursor Protein Products Concentrate in a Subset of Exosomes Specifically Endocytosed by Neurons. Cell. Mol. Life Sci. CMLS 2018, 75, 757–773. [Google Scholar] [CrossRef] [PubMed]
- van Balkom, B.W.M.; de Jong, O.G.; Smits, M.; Brummelman, J.; den Ouden, K.; de Bree, P.M.; van Eijndhoven, M.A.J.; Pegtel, D.M.; Stoorvogel, W.; Würdinger, T.; et al. Endothelial Cells Require MiR-214 to Secrete Exosomes That Suppress Senescence and Induce Angiogenesis in Human and Mouse Endothelial Cells. Blood 2013, 121, 3997–4006. [Google Scholar] [CrossRef]
- Zhu, F.; Chong Lee Shin, O.L.S.; Pei, G.; Hu, Z.; Yang, J.; Zhu, H.; Wang, M.; Mou, J.; Sun, J.; Wang, Y.; et al. Adipose-Derived Mesenchymal Stem Cells Employed Exosomes to Attenuate AKI-CKD Transition through Tubular Epithelial Cell Dependent Sox9 Activation. Oncotarget 2017, 8, 70707–70726. [Google Scholar] [CrossRef]
- Tauro, B.J.; Mathias, R.A.; Greening, D.W.; Gopal, S.K.; Ji, H.; Kapp, E.A.; Coleman, B.M.; Hill, A.F.; Kusebauch, U.; Hallows, J.L.; et al. Oncogenic H-Ras Reprograms Madin-Darby Canine Kidney (MDCK) Cell-Derived Exosomal Proteins Following Epithelial-Mesenchymal Transition. Mol. Cell. Proteomics MCP 2013, 12, 2148–2159. [Google Scholar] [CrossRef]
- Yang, T.T.; Liu, C.G.; Gao, S.C.; Zhang, Y.; Wang, P.C. The Serum Exosome Derived MicroRNA-135a, -193b, and -384 Were Potential Alzheimer’s Disease Biomarkers. Biomed. Environ. Sci. BES 2018, 31, 87–96. [Google Scholar] [CrossRef]
- Foers, A.D.; Chatfield, S.; Dagley, L.F.; Scicluna, B.J.; Webb, A.I.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Enrichment of Extracellular Vesicles from Human Synovial Fluid Using Size Exclusion Chromatography. J. Extracell. Vesicles 2018, 7, 1490145. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Yi, D.Y. Analysis of the Human Breast Milk Microbiome and Bacterial Extracellular Vesicles in Healthy Mothers. Exp. Mol. Med. 2020, 52, 1288–1297. [Google Scholar] [CrossRef]
- Cheng, L.; Sun, X.; Scicluna, B.J.; Coleman, B.M.; Hill, A.F. Characterization and Deep Sequencing Analysis of Exosomal and Non-Exosomal MiRNA in Human Urine. Kidney Int. 2014, 86, 433–444. Available online: https://pubmed.ncbi.nlm.nih.gov/24352158/ (accessed on 3 August 2022). [CrossRef] [PubMed]
- Yap, T.; Koo, K.; Cheng, L.; Vella, L.J.; Hill, A.F.; Reynolds, E.; Nastri, A.; Cirillo, N.; Seers, C.; McCullough, M. Predicting the Presence of Oral Squamous Cell Carcinoma Using Commonly Dysregulated MicroRNA in Oral Swirls. Cancer Prev. Res. Phila. Pa 2018, 11, 491–502. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Wollert, T.; Hurley, J.H. Molecular Mechanism of Multivesicular Body Biogenesis by ESCRT Complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. Available online: https://pubmed.ncbi.nlm.nih.gov/32899828/ (accessed on 22 August 2022). [CrossRef]
- Coleman, B.M.; Hanssen, E.; Lawson, V.A.; Hill, A.F. Prion-Infected Cells Regulate the Release of Exosomes with Distinct Ultrastructural Features. FASEB J. 2012, 26, 4160–4173. Available online: https://pubmed.ncbi.nlm.nih.gov/22767229/ (accessed on 4 August 2022). [CrossRef]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445.e18. [Google Scholar] [CrossRef] [PubMed]
- Koifman, N.; Biran, I.; Aharon, A.; Brenner, B.; Talmon, Y. A Direct-Imaging Cryo-EM Study of Shedding Extracellular Vesicles from Leukemic Monocytes. J. Struct. Biol. 2017, 198, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of Secretion and Uptake of Exosomes and Other Extracellular Vesicles for Cell-to-Cell Communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Piccin, A.; Murphy, W.G.; Smith, O.P. Circulating Microparticles: Pathophysiology and Clinical Implications. Blood Rev. 2007, 21, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Huttner, W.B.; Zimmerberg, J. Implications of Lipid Microdomains for Membrane Curvature, Budding and Fission. Curr. Opin. Cell Biol. 2001, 13, 478–484. [Google Scholar] [CrossRef]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct RNA Profiles in Subpopulations of Extracellular Vesicles: Apoptotic Bodies, Microvesicles and Exosomes. J. Extracell. Vesicles 2013, 2, 20677. [Google Scholar] [CrossRef] [PubMed]
- Lunavat, T.R.; Cheng, L.; Kim, D.-K.; Bhadury, J.; Jang, S.C.; Lässer, C.; Sharples, R.A.; López, M.D.; Nilsson, J.; Gho, Y.S.; et al. Small RNA Deep Sequencing Discriminates Subsets of Extracellular Vesicles Released by Melanoma Cells--Evidence of Unique MicroRNA Cargos. RNA Biol. 2015, 12, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Atkin-Smith, G.K.; Tixeira, R.; Paone, S.; Mathivanan, S.; Collins, C.; Liem, M.; Goodall, K.J.; Ravichandran, K.S.; Hulett, M.D.; Poon, I.K.H. A Novel Mechanism of Generating Extracellular Vesicles during Apoptosis via a Beads-on-a-String Membrane Structure. Nat. Commun. 2015, 6, 7439. [Google Scholar] [CrossRef]
- Moss, D.K.; Betin, V.M.; Malesinski, S.D.; Lane, J.D. A Novel Role for Microtubules in Apoptotic Chromatin Dynamics and Cellular Fragmentation. J. Cell Sci. 2006, 119, 2362–2374. [Google Scholar] [CrossRef] [PubMed]
- Poon, I.K.; Chiu, Y.H.; Armstrong, A.J.; Kinchen, J.M.; Juncadella, I.J.; Bayliss, D.A.; Ravichandran, K.S. Unexpected Link between an Antibiotic, Pannexin Channels and Apoptosis. Nature 2014, 507, 329–334. Available online: https://pubmed.ncbi.nlm.nih.gov/24646995/ (accessed on 4 August 2022). [CrossRef]
- Lane, J.D.; Allan, V.J.; Woodman, P.G. Active Relocation of Chromatin and Endoplasmic Reticulum into Blebs in Late Apoptotic Cells. J. Cell Sci. 2005, 118, 4059–4071. [Google Scholar] [CrossRef]
- Torr, E.E.; Gardner, D.H.; Thomas, L.; Goodall, D.M.; Bielemeier, A.; Willetts, R.; Griffiths, H.R.; Marshall, L.J.; Devitt, A. Apoptotic Cell-Derived ICAM-3 Promotes Both Macrophage Chemoattraction to and Tethering of Apoptotic Cells. Cell Death Differ. 2012, 19, 671–679. [Google Scholar] [CrossRef]
- Ravichandran, K.S. Beginnings of a Good Apoptotic Meal: The Find-Me and Eat-Me Signaling Pathways. Immunity 2011, 35, 445–455. [Google Scholar] [CrossRef]
- Lin, L.; Lu, L.; Cao, W.; Li, T. Hypothesis for Potential Pathogenesis of SARS-CoV-2 Infection—A Review of Immune Changes in Patients with Viral Pneumonia. Emerg. Microbes Infect. 2020, 9, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Gautret, P.; Million, M.; Jarrot, P.-A.; Camoin-Jau, L.; Colson, P.; Fenollar, F.; Leone, M.; La Scola, B.; Devaux, C.; Gaubert, J.Y.; et al. Natural History of COVID-19 and Therapeutic Options. Expert Rev. Clin. Immunol. 2020, 16, 1159–1184. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H.; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef]
- Yang, L.; Xie, X.; Tu, Z.; Fu, J.; Xu, D.; Zhou, Y. The Signal Pathways and Treatment of Cytokine Storm in COVID-19. Signal Transduct. Target. Ther. 2021, 6, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Tavakolpour, S.; Rakhshandehroo, T.; Wei, E.X.; Rashidian, M. Lymphopenia during the COVID-19 Infection: What It Shows and What Can Be Learned. Immunol. Lett. 2020, 225, 31–32. [Google Scholar] [CrossRef]
- Mitsuyama, Y.; Yamakawa, K.; Kayano, K.; Maruyama, M.; Wada, T.; Fujimi, S. Prolonged Enhancement of Cytotoxic T Lymphocytes in the Post-Recovery State of Severe COVID-19. J. Intensive Care 2021, 9, 76. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Zou, Z.; Ren, D.; Chen, R.; Yu, B.; Liu, Y.; Huang, J.; Yang, Z.; Zhou, Z.; Feng, Y.; Wu, M. Persistent Lymphopenia after Diagnosis of COVID-19 Predicts Acute Respiratory Distress Syndrome: A Retrospective Cohort Study. Eur. J. Inflamm. 2021, 19, 20587392211036824. [Google Scholar] [CrossRef]
- Channappanavar, R.; Fehr, A.R.; Vijay, R.; Mack, M.; Zhao, J.; Meyerholz, D.K.; Perlman, S. Dysregulated Type I Interferon and Inflammatory Monocyte-Macrophage Responses Cause Lethal Pneumonia in SARS-CoV-Infected Mice. Cell Host Microbe 2016, 19, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, B.; Mao, J. The Pathogenesis and Treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Kaler, J.; Tabrez, E.; Tabrez, S.; Tabrez, S.S.M. Novel COVID-19: A Comprehensive Review of Transmission, Manifestation, and Pathogenesis. Cureus 2020, 12, e8184. [Google Scholar] [CrossRef]
- Bellani, G.; Laffey, J.G.; Pham, T.; Fan, E.; Brochard, L.; Esteban, A.; Gattinoni, L.; van Haren, F.; Larsson, A.; McAuley, D.F.; et al. Epidemiology, Patterns of Care, and Mortality for Patients With Acute Respiratory Distress Syndrome in Intensive Care Units in 50 Countries. JAMA 2016, 315, 788–800. [Google Scholar] [CrossRef]
- Arachchillage, D.R.J.; Laffan, M. Abnormal Coagulation Parameters Are Associated with Poor Prognosis in Patients with Novel Coronavirus Pneumonia. J. Thromb. Haemost. 2020, 18, 1233–1234. [Google Scholar] [CrossRef]
- Barberis, E.; Vanella, V.V.; Falasca, M.; Caneapero, V.; Cappellano, G.; Raineri, D.; Ghirimoldi, M.; De Giorgis, V.; Puricelli, C.; Vaschetto, R.; et al. Circulating Exosomes Are Strongly Involved in SARS-CoV-2 Infection. Front. Mol. Biosci. 2021, 8, 632290. [Google Scholar] [CrossRef]
- Zicari, S.; Arakelyan, A.; Palomino, R.A.Ñ.; Fitzgerald, W.; Vanpouille, C.; Lebedeva, A.; Schmitt, A.; Bomsel, M.; Britt, W.; Margolis, L. Human Cytomegalovirus-Infected Cells Release Extracellular Vesicles That Carry Viral Surface Proteins. Virology 2018, 524, 97–105. [Google Scholar] [CrossRef]
- Chelvanambi, S.; Bogatcheva, N.V.; Bednorz, M.; Agarwal, S.; Maier, B.; Alves, N.J.; Li, W.; Syed, F.; Saber, M.M.; Dahl, N.; et al. HIV-Nef Protein Persists in the Lungs of Aviremic Patients with HIV and Induces Endothelial Cell Death. Am. J. Respir. Cell Mol. Biol. 2019, 60, 357–366. [Google Scholar] [CrossRef]
- Dogrammatzis, C.; Saleh, S.; Deighan, C.; Kalamvoki, M. Diverse Populations of Extracellular Vesicles with Opposite Functions during Herpes Simplex Virus 1 Infection. J. Virol. 2021, 95, e02357-20. Available online: https://pubmed.ncbi.nlm.nih.gov/33361424/ (accessed on 16 August 2022). [CrossRef]
- Earnest, J.T.; Hantak, M.P.; Li, K.; Jr, P.B.M.; Perlman, S.; Gallagher, T. The Tetraspanin CD9 Facilitates MERS-Coronavirus Entry by Scaffolding Host Cell Receptors and Proteases. PLoS Pathog. 2017, 13, e1006546. [Google Scholar] [CrossRef]
- Böker, K.O.; Lemus-Diaz, N.; Rinaldi Ferreira, R.; Schiller, L.; Schneider, S.; Gruber, J. The Impact of the CD9 Tetraspanin on Lentivirus Infectivity and Exosome Secretion. Mol. Ther. 2018, 26, 634–647. [Google Scholar] [CrossRef]
- Tey, S.K.; Lam, H.; Wong, S.W.K.; Zhao, H.; To, K.K.-W.; Yam, J.W.P. ACE2-Enriched Extracellular Vesicles Enhance Infectivity of Live SARS-CoV-2 Virus. J. Extracell. Vesicles 2022, 11, e12231. [Google Scholar] [CrossRef]
- Troyer, Z.; Alhusaini, N.; Tabler, C.O.; Sweet, T.; de Carvalho, K.I.L.; Schlatzer, D.M.; Carias, L.; King, C.L.; Matreyek, K.; Tilton, J.C. Extracellular Vesicles Carry SARS-CoV-2 Spike Protein and Serve as Decoys for Neutralizing Antibodies. J. Extracell. Vesicles 2021, 10, e12112. [Google Scholar] [CrossRef]
- Pesce, E.; Manfrini, N.; Cordiglieri, C.; Santi, S.; Bandera, A.; Gobbini, A.; Gruarin, P.; Favalli, A.; Bombaci, M.; Cuomo, A.; et al. Exosomes Recovered From the Plasma of COVID-19 Patients Expose SARS-CoV-2 Spike-Derived Fragments and Contribute to the Adaptive Immune Response. Front. Immunol. 2021, 12, 785941. [Google Scholar] [CrossRef]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B Lymphocytes Secrete Antigen-Presenting Vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef]
- Bruno, S.; Collino, F.; Deregibus, M.C.; Grange, C.; Tetta, C.; Camussi, G. Microvesicles Derived from Human Bone Marrow Mesenchymal Stem Cells Inhibit Tumor Growth. Stem Cells Dev. 2013, 22, 758–771. [Google Scholar] [CrossRef]
- Claridge, B.; Lozano, J.; Poh, Q.H.; Greening, D.W. Development of Extracellular Vesicle Therapeutics: Challenges, Considerations, and Opportunities. Front. Cell Dev. Biol. 2021, 9, 734720. [Google Scholar] [CrossRef]
- Inal, J.M. Decoy ACE2-Expressing Extracellular Vesicles That Competitively Bind SARS-CoV-2 as a Possible COVID-19 Therapy. Clin. Sci. 2020, 134, 1301–1304. [Google Scholar] [CrossRef]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J. Delivery of SiRNA to the Mouse Brain by Systemic Injection of Targeted Exosomes. Nat. Biotechnol. 2011, 29, 341–345. Available online: https://www.nature.com/articles/nbt.1807 (accessed on 5 August 2022). [CrossRef]
- Yang, Y.; Hong, Y.; Nam, G.-H.; Chung, J.H.; Koh, E.; Kim, I.-S. Virus-Mimetic Fusogenic Exosomes for Direct Delivery of Integral Membrane Proteins to Target Cell Membranes. Adv. Mater. 2017, 29, 1605604. [Google Scholar] [CrossRef]
- Nolte-‘t Hoen, E.; Cremer, T.; Gallo, R.C.; Margolis, L.B. Extracellular Vesicles and Viruses: Are They Close Relatives? Proc. Natl. Acad. Sci. USA 2016, 113, 9155–9161. Available online: https://www.pnas.org/doi/full/10.1073/pnas.1605146113 (accessed on 5 August 2022). [CrossRef]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Zhou, Y.; Du, L. Characterization of the Receptor-Binding Domain (RBD) of 2019 Novel Coronavirus: Implication for Development of RBD Protein as a Viral Attachment Inhibitor and Vaccine. Cell. Mol. Immunol. 2020, 17, 613–620. [Google Scholar] [CrossRef]
- Benítez-Cardoza, C.G.; Vique-Sánchez, J.L. Potential Inhibitors of the Interaction between ACE2 and SARS-CoV-2 (RBD), to Develop a Drug. Life Sci. 2020, 256, 117970. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell Entry Mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Li, M.Y.; Li, L.; Zhang, Y.; Wang, X.S. Expression of the SARS-CoV-2 Cell Receptor Gene ACE2 in a Wide Variety of Human Tissues. Infect. Dis. Poverty 2020, 9, 23–29. Available online: https://mednexus.org/doi/full/10.1186/s40249-020-00662-x (accessed on 4 August 2022). [CrossRef]
- Lv, Z.; Deng, Y.Q.; Ye, Q.; Cao, L.; Sun, C.Y.; Fan, C.; Huang, W.; Sun, S.; Sun, Y.; Zhu, L.; et al. Structural Basis for Neutralization of SARS-CoV-2 and SARS-CoV by a Potent Therapeutic Antibody. Science 2020, 369, 1505–1509. Available online: https://www.science.org/doi/10.1126/science.abc5881 (accessed on 4 August 2022). [CrossRef]
- Andreu, Z.; Yáñez-Mó, M. Tetraspanins in Extracellular Vesicle Formation and Function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef]
- Harcourt, J.; Tamin, A.; Lu, X.; Kamili, S.; Sakthivel, S.K.; Murray, J.; Queen, K.; Tao, Y.; Paden, C.R.; Zhang, J.; et al. Isolation and Characterization of SARS-CoV-2 from the First US COVID-19 Patient. bioRxiv 2020. [Google Scholar] [CrossRef]
- Xia, X.; Yuan, P.; Liu, Y.; Wang, Y.; Cao, W.; Zheng, J.C. Emerging Roles of Extracellular Vesicles in COVID-19, a Double-Edged Sword? Immunology 2021, 163, 416–430. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Rothan, H.A.; Byrareddy, S.N. The Epidemiology and Pathogenesis of Coronavirus Disease (COVID-19) Outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef]
- Zhang, Q.; Jeppesen, D.K.; Higginbotham, J.N.; Franklin, J.L.; Crowe, J.E.; Coffey, R.J. Angiotensin-Converting Enzyme 2–Containing Small Extracellular Vesicles and Exomeres Bind the Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein. Gastroenterology 2021, 160, 958–961. [Google Scholar] [CrossRef]
- Ayre, D.C.; Christian, S.L. CD24: A Rheostat That Modulates Cell Surface Receptor Signaling of Diverse Receptors. Front. Cell Dev. Biol. 2016, 4, 146. [Google Scholar] [CrossRef]
- Altevogt, P.; Sammar, M.; Hüser, L.; Kristiansen, G. Novel Insights into the Function of CD24: A Driving Force in Cancer. Int. J. Cancer 2021, 148, 546–559. [Google Scholar] [CrossRef]
- Kristiansen, G.; Machado, E.; Bretz, N.; Rupp, C.; Winzer, K.-J.; König, A.-K.; Moldenhauer, G.; Marmé, F.; Costa, J.; Altevogt, P. Molecular and Clinical Dissection of CD24 Antibody Specificity by a Comprehensive Comparative Analysis. Lab. Investig. J. Tech. Methods Pathol. 2010, 90, 1102–1116. [Google Scholar] [CrossRef]
- Chen, G.-Y.; Tang, J.; Zheng, P.; Liu, Y. CD24 and Siglec-10 Selectively Repress Tissue Damage-Induced Immune Responses. Science 2009, 323, 1722–1725. [Google Scholar] [CrossRef]
- Avril, T.; Freeman, S.D.; Attrill, H.; Clarke, R.G.; Crocker, P.R. Siglec-5 (CD170) Can Mediate Inhibitory Signaling in the Absence of Immunoreceptor Tyrosine-Based Inhibitory Motif Phosphorylation. J. Biol. Chem. 2005, 280, 19843–19851. [Google Scholar] [CrossRef]
- Tian, R.-R.; Zhang, M.-X.; Zhang, L.-T.; Zhang, P.; Ma, J.-P.; Liu, M.; Devenport, M.; Zheng, P.; Zhang, X.-L.; Lian, X.-D.; et al. CD24 and Fc Fusion Protein Protects SIVmac239-Infected Chinese Rhesus Macaque against Progression to AIDS. Antivir. Res. 2018, 157, 9–17. [Google Scholar] [CrossRef]
- Sagiv, E.; Portman, M.A. CD24 for Cardiovascular Researchers: A Key Molecule in Cardiac Immunology, Marker of Stem Cells and Target for Drug Development. J. Pers. Med. 2021, 11, 260. [Google Scholar] [CrossRef]
- Duan, K.; Liu, B.; Li, C.; Zhang, H.; Yu, T.; Qu, J.; Zhou, M.; Chen, L.; Meng, S.; Hu, Y.; et al. Effectiveness of Convalescent Plasma Therapy in Severe COVID-19 Patients. Proc. Natl. Acad. Sci. USA 2020, 117, 9490–9496. [Google Scholar] [CrossRef]
- Meher, B.R.; Padhy, B.M.; Das, S.; Mohanty, R.R.; Agrawal, K. Effectiveness of Convalescent Plasma Therapy in the Treatment of Moderate to Severe COVID 19 Patients: A Systematic Review and Meta-Analysis. J. Assoc. Physicians India 2020, 68, 35–43. [Google Scholar]
- Klamt, F.; Parsons, R.B.; Jones, M.H. Plasma Therapy to Prevent Severe COVID-19 in Older Adults. N. Engl. J. Med. 2021, 384, e104. [Google Scholar] [CrossRef] [PubMed]
- Karn, V.; Ahmed, S.; Tsai, L.-W.; Dubey, R.; Ojha, S.; Singh, H.N.; Kumar, M.; Gupta, P.K.; Sadhu, S.; Jha, N.K.; et al. Extracellular Vesicle-Based Therapy for COVID-19: Promises, Challenges and Future Prospects. Biomedicines 2021, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Nanomedicine and the COVID-19 Vaccines. Nat. Nanotechnol. 2020, 15, 963. [CrossRef] [PubMed]
- SeyedAlinaghi, S.; Karimi, A.; Pashaei, Z.; Afzalian, A.; Mirzapour, P.; Ghorbanzadeh, K.; Ghasemzadeh, A.; Dashti, M.; Nazarian, N.; Vahedi, F.; et al. Safety and Adverse Events Related to COVID-19 MRNA Vaccines; A Systematic Review. Arch. Acad. Emerg. Med. 2022, 10, e41. [Google Scholar] [CrossRef]
- Anand, P.; Stahel, V.P. Review the Safety of Covid-19 MRNA Vaccines: A Review. Patient Saf. Surg. 2021, 15, 20. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Li, X.; Zhang, Y.; Han, Y.; Chang, F.; Ding, J. Mesenchymal Stem Cells for Regenerative Medicine. Cells 2019, 8, 886. [Google Scholar] [CrossRef] [PubMed]
- Satija, N.K.; Singh, V.K.; Verma, Y.K.; Gupta, P.; Sharma, S.; Afrin, F.; Sharma, M.; Sharma, P.; Tripathi, R.P.; Gurudutta, G.U. Mesenchymal Stem Cell-Based Therapy: A New Paradigm in Regenerative Medicine. J. Cell. Mol. Med. 2009, 13, 4385–4402. [Google Scholar] [CrossRef]
- Uemura, R.; Xu, M.; Ahmad, N.; Ashraf, M. Bone Marrow Stem Cells Prevent Left Ventricular Remodeling of Ischemic Heart through Paracrine Signaling. Circ. Res. 2006, 98, 1414–1421. [Google Scholar] [CrossRef]
- Krishnan, A.; Muthusamy, S.; Fernandez, F.B.; Kasoju, N. Mesenchymal Stem Cell-Derived Extracellular Vesicles in the Management of COVID19-Associated Lung Injury: A Review on Publications, Clinical Trials and Patent Landscape. Tissue Eng. Regen. Med. 2022, 19, 659–673. [Google Scholar] [CrossRef]
- Qian, X.; Xu, C.; Fang, S.; Zhao, P.; Wang, Y.; Liu, H.; Yuan, W.; Qi, Z. Exosomal MicroRNAs Derived From Umbilical Mesenchymal Stem Cells Inhibit Hepatitis C Virus Infection. Stem Cells Transl. Med. 2016, 5, 1190–1203. [Google Scholar] [CrossRef]
- Schultz, I.C.; Bertoni, A.P.S.; Wink, M.R. Mesenchymal Stem Cell-Derived Extracellular Vesicles Carrying MiRNA as a Potential Multi Target Therapy to COVID-19: An In Silico Analysis. Stem Cell Rev. Rep. 2021, 17, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Vasandan, A.B.; Jahnavi, S.; Shashank, C.; Prasad, P.; Kumar, A.; Prasanna, S.J. Human Mesenchymal Stem Cells Program Macrophage Plasticity by Altering Their Metabolic Status via a PGE2-Dependent Mechanism. Sci. Rep. 2016, 6, 38308. [Google Scholar] [CrossRef] [PubMed]
- Devaney, J.; Horie, S.; Masterson, C.; Elliman, S.; Barry, F.; O’Brien, T.; Curley, G.F.; O’Toole, D.; Laffey, J.G. Human Mesenchymal Stromal Cells Decrease the Severity of Acute Lung Injury Induced by E. Coli in the Rat. Thorax 2015, 70, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-S.; Svennerholm, K.; Shelke, G.V.; Bandeira, E.; Lässer, C.; Jang, S.C.; Chandode, R.; Gribonika, I.; Lötvall, J. Mesenchymal Stromal Cell-Derived Nanovesicles Ameliorate Bacterial Outer Membrane Vesicle-Induced Sepsis via IL-10. Stem Cell Res. Ther. 2019, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Mitrani, M.I.; Bellio, M.A.; Sagel, A.; Saylor, M.; Kapp, W.; VanOsdol, K.; Haskell, G.; Stewart, D.; Abdullah, Z.; Santos, I.; et al. Case Report: Administration of Amniotic Fluid-Derived Nanoparticles in Three Severely Ill COVID-19 Patients. Front. Med. 2021, 8, 583842. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).