
Lysyl Oxidases: Orchestrators of Cellular Behavior and ECM Remodeling and Homeostasis

{kind=link}
{kind=link}
Abstract
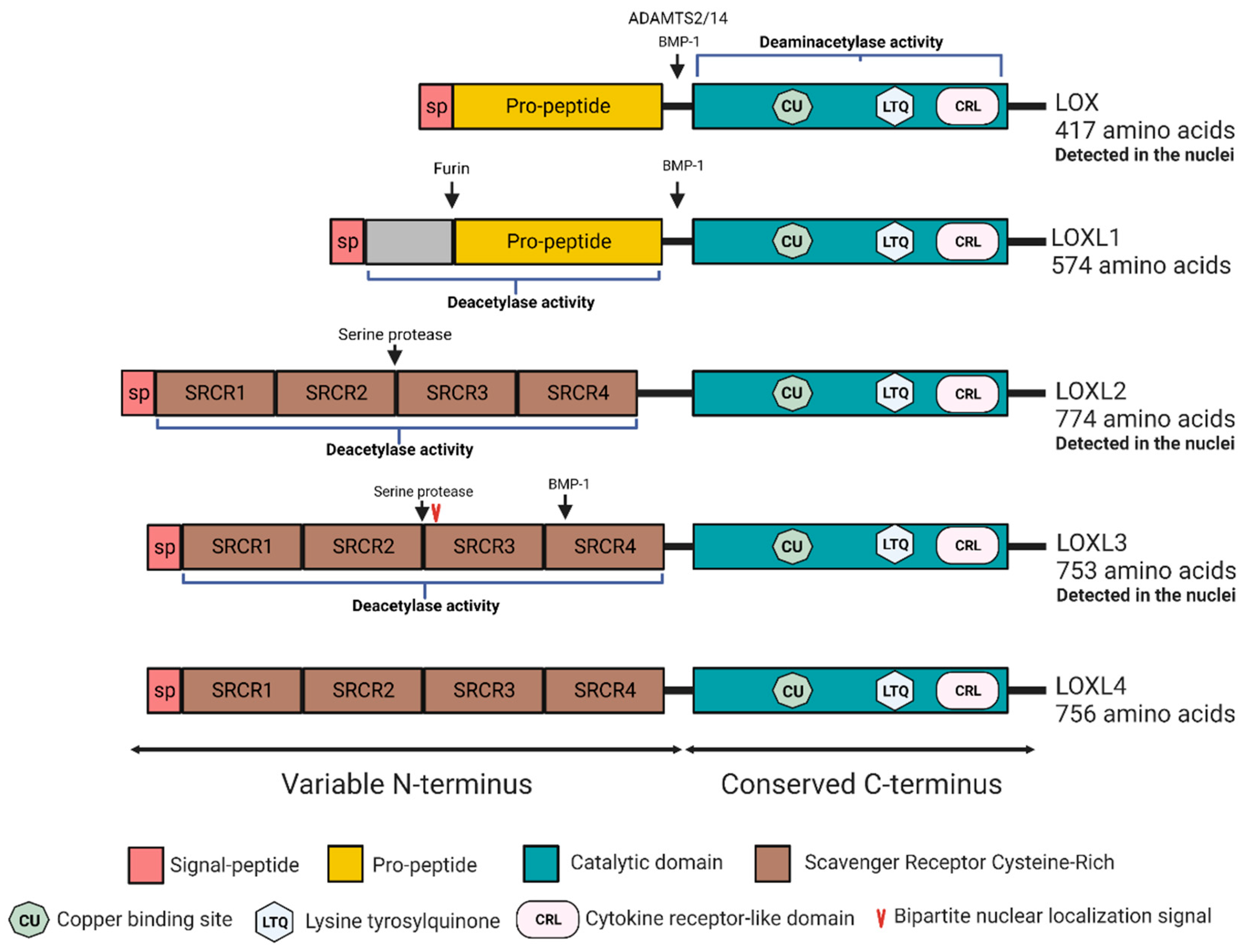
:1. Introduction
2. Lysyl Oxidases: Multi-Faceted Regulators—The Case of EMT
3. Lysyl Oxidases Regulate Cytoskeletal Assembly
4. LOX-Dependent Transcriptional Regulation
5. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kagan, H.M.; Trackman, P.C. Properties and Function of Lysyl Oxidase. Am. J. Respir. Cell Mol. Biol. 1991, 5, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Palamakumbura, A.H.; Trackman, P.C. A Fluorometric Assay for Detection of Lysyl Oxidase Enzyme Activity in Biological Samples. Anal. Biochem. 2002, 300, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Kagan, H.M.; Li, W. Lysyl Oxidase: Properties, Specificity, and Biological Roles inside and Outside of the Cell. J. Cell Biochem. 2003, 88, 660–672. [Google Scholar] [CrossRef] [PubMed]
- Pinnell, S.R.; Martin, G.R. The Cross-Linking of Collagen and Elastin: Enzymatic Conversion of Lysine in Peptide Linkage to Alpha-Aminoadipic-Delta-Semialdehyde (Allysine) by an Extract from Bone. Proc. Natl. Acad. Sci. USA 1968, 61, 708–716. [Google Scholar] [CrossRef]
- Levene, C.I. Plant Toxins and Human Disease. Collagen and Lathyrism. Proc. R. Soc. Med. 1966, 59, 757–758. [Google Scholar]
- Selye, H. Lathyrism. Rev. Can. De Biol. 1957, 16, 1–82. [Google Scholar]
- Wang, S.X.; Mure, M.; Medzihradszky, K.F.; Burlingame, A.L.; Brown, D.E.; Dooley, D.M.; Smith, A.J.; Kagan, H.M.; Klinman, J.P. A Crosslinked Cofactor in Lysyl Oxidase: Redox Function for Amino Acid Side Chains. Science 1996, 273, 1078–1084. [Google Scholar] [CrossRef]
- Rosell-García, T.; Paradela, A.; Bravo, G.; Dupont, L.; Bekhouche, M.; Colige, A.; Rodriguez-Pascual, F. Differential Cleavage of Lysyl Oxidase by the Metalloproteinases BMP1 and ADAMTS2/14 Regulates Collagen Binding through a Tyrosine Sulfate Domain. J. Biol. Chem. 2019, 294, 11087–11100. [Google Scholar] [CrossRef]
- Rosell-García, T.; Rivas-Muñoz, S.; Colige, A.; Rodriguez-Pascual, F. Cleavage of LOXL1 by BMP1 and ADAMTS14 Proteases Suggests a Role for Proteolytic Processing in the Regulation of LOXL1 Function. Int. J. Mol. Sci. 2022, 23, 3285. [Google Scholar] [CrossRef]
- Gabay Yehezkely, R.; Zaffryar-Eilot, S.; Kaganovsky, A.; Fainshtain Malka, N.; Aviram, R.; Livneh, I.; Hasson, P. Intracellular Role for the Matrix Modifying Enzyme Lox in Regulating Transcription Factor Subcellular Localization and Activity in Muscle Regeneration. Dev. Cell 2020, 53, 406–417. [Google Scholar] [CrossRef]
- Yaseen, W.; Kraft-Sheleg, O.; Zaffryar-Eilot, S.; Melamed, S.; Sun, C.; Millay, D.P.; Hasson, P. Fibroblast Fusion to the Muscle Fiber Regulates Myotendinous Junction Formation. Nat. Commun. 2021, 12, 3852. [Google Scholar] [CrossRef] [PubMed]
- Kutchuk, L.; Laitala, A.; Soueid-Bomgarten, S.; Shentzer, P.; Rosendahl, A.-H.; Eilot, S.; Grossman, M.; Sagi, I.; Sormunen, R.; Myllyharju, J.; et al. Muscle Composition Is Regulated by a Lox-TGFβ Feedback Loop. Development (Cambridge) 2015, 142, 983–993. [Google Scholar] [CrossRef] [Green Version]
- Mäki, J.M.; Sormunen, R.; Lippo, S.; Kaarteenaho-Wiik, R.; Soininen, R.; Myllyharju, J. Lysyl Oxidase Is Essential for Normal Development and Function of the Respiratory System and for the Integrity of Elastic and Collagen Fibers in Various Tissues. Am. J. Pathol. 2005, 167, 927–936. [Google Scholar] [CrossRef]
- Hornstra, I.K.; Birge, S.; Starcher, B.; Bailey, A.J.; Mecham, R.P.; Shapiro, S.D. Lysyl Oxidase Is Required for Vascular and Diaphragmatic Development in Mice. J. Biol. Chem. 2003, 278, 14387–14393. [Google Scholar] [CrossRef] [PubMed]
- Vallet, S.D.; Ricard-Blum, S. Lysyl Oxidases: From Enzyme Activity to Extracellular Matrix Cross-Links. Essays Biochem. 2019, 63, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Peinado, H.; Del Carmen Iglesias-de la Cruz, M.; Olmeda, D.; Csiszar, K.; Fong, K.S.; Vega, S.; Nieto, M.A.; Cano, A.; Portillo, F. A Molecular Role for Lysyl Oxidase-like 2 Enzyme in Snail Regulation and Tumor Progression. EMBO J. 2005, 24, 3446–3458. [Google Scholar] [CrossRef]
- Moon, H.J.; Finney, J.; Xu, L.; Moore, D.; Welch, D.R.; Mure, M. MCF-7 Cells Expressing Nuclear Associated Lysyl Oxidase-like 2 (LOXL2) Exhibit an Epithelial-to-Mesenchymal Transition (EMT) Phenotype and Are Highly Invasive in Vitro. J. Biol. Chem. 2013, 288, 30000–30008. [Google Scholar] [CrossRef]
- Schietke, R.; Warnecke, C.; Wacker, I.; Schödel, J.; Mole, D.R.; Campean, V.; Amann, K.; Goppelt-Struebe, M.; Behrens, J.; Eckardt, K.U.; et al. The Lysyl Oxidases LOX and LOXL2 Are Necessary and Sufficient to Repress E-Cadherin in Hypoxia: Insights into Cellular Transformation Processes Mediated by HIF-1. J. Biol. Chem. 2010, 285, 6658–6669. [Google Scholar] [CrossRef]
- Kasashima, H.; Yashiro, M.; Kinoshita, H.; Fukuoka, T.; Morisaki, T.; Masuda, G.; Sakurai, K.; Kubo, N.; Ohira, M.; Hirakawa, K. Lysyl Oxidase Is Associated with the Epithelial-Mesenchymal Transition of Gastric Cancer Cells in Hypoxia. Gastric Cancer Off. J. Int. Gastric Cancer Assoc. Jpn. Gastric Cancer Assoc. 2016, 19, 431–442. [Google Scholar] [CrossRef]
- Hay, E.D. An Overview of Epithelio-Mesenchymal Transformation. Acta Anat. 1995, 154, 8–20. [Google Scholar] [CrossRef]
- Akiri, G.; Sabo, E.; Dafni, H.; Vadasz, Z.; Kartvelishvily, Y.; Gan, N.; Kessler, O.; Cohen, T.; Resnick, M.; Neeman, M.; et al. Lysyl Oxidase-Related Protein-1 Promotes Tumor Fibrosis and Tumor Progression in Vivo. Cancer Res. 2003, 63, 1657–1666. [Google Scholar]
- Moreno-Bueno, G.; Portillo, F.; Cano, A. Transcriptional Regulation of Cell Polarity in EMT and Cancer. Oncogene 2008, 27, 6958–6969. [Google Scholar] [CrossRef]
- Hugo, H.; Ackland, M.L.; Blick, T.; Lawrence, M.G.; Clements, J.A.; Williams, E.D.; Thompson, E.W. Epithelial–Mesenchymal and Mesenchymal–Epithelial Transitions in Carcinoma Progression. J. Cell. Physiol. 2007, 213, 374–383. [Google Scholar] [CrossRef]
- Brekhman, V.; Lugassie, J.; Zaffryar-Eilot, S.; Sabo, E.; Kessler, O.; Smith, V.; Golding, H.; Neufeld, G. Receptor Activity Modifying Protein-3 Mediates the Protumorigenic Activity of Lysyl Oxidase-like Protein-2. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 55–65. [Google Scholar] [CrossRef]
- Their, J.P. Epithelial-Mesenchymal Transitions in Tumor Progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar] [CrossRef]
- Cano, A.; Pérez-Moreno, M.A.; Rodrigo, I.; Locascio, A.; Blanco, M.J.; Del Barrio, M.G.; Portillo, F.; Nieto, M.A. The Transcription Factor Snail Controls Epithelial-Mesenchymal Transitions by Repressing E-Cadherin Expression. Nat. Cell Biol. 2000, 2, 76–83. [Google Scholar] [CrossRef]
- Canesin, G.; Cuevas, E.P.; Santos, V.; López-Menéndez, C.; Moreno-Bueno, G.; Huang, Y.; Csiszar, K.; Portillo, F.; Peinado, H.; Lyden, D.; et al. Lysyl Oxidase-like 2 (LOXL2) and E47 EMT Factor: Novel Partners in E-Cadherin Repression and Early Metastasis Colonization. Oncogene 2015, 34, 951–964. [Google Scholar] [CrossRef]
- Boufraqech, M.; Zhang, L.; Nilubol, N.; Sadowski, S.M.; Kotian, S.; Quezado, M.; Kebebew, E. Lysyl Oxidase (LOX) Transcriptionally Regulates SNAI2 Expression and TIMP4 Secretion in Human Cancers. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 4491–4504. [Google Scholar] [CrossRef]
- Peng, L.; Ran, Y.L.; Hu, H.; Yu, L.; Liu, Q.; Zhou, Z.; Sun, Y.M.; Sun, L.C.; Pan, J.; Sun, L.X.; et al. Secreted LOXL2 Is a Novel Therapeutic Target That Promotes Gastric Cancer Metastasis via the Src/FAK Pathway. Carcinogenesis 2009, 30, 1660–1669. [Google Scholar] [CrossRef]
- Payne, S.L.; Fogelgren, B.; Hess, A.R.; Seftor, E.A.; Wiley, E.L.; Fong, S.F.; Csiszar, K.; Hendrix, M.J.; Kirschmann, D.A. Lysyl Oxidase Regulates Breast Cancer Cell Migration and Adhesion through a Hydrogen Peroxide-Mediated Mechanism. Cancer Res. 2005, 65, 11429–11436. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Bueno, G.; Salvador, F.; Martín, A.; Floristán, A.; Cuevas, E.P.; Santos, V.; Montes, A.; Morales, S.; Castilla, M.A.; Rojo-Sebastián, A.; et al. Lysyl Oxidase-like 2 (LOXL2), a New Regulator of Cell Polarity Required for Metastatic Dissemination of Basal-like Breast Carcinomas. EMBO Mol. Med. 2011, 3, 528–544. [Google Scholar] [CrossRef]
- Zhan, X.H.; Jiao, J.W.; Zhang, H.F.; Xu, X.E.; He, J.Z.; Li, R.L.; Zou, H.Y.; Wu, Z.Y.; Wang, S.H.; Wu, J.Y.; et al. LOxL2 Upregulates Phosphorylation of Ezrin to Promote Cytoskeletal Reorganization and Tumor Cell Invasion. Cancer Res. 2019, 79, 4951–4964. [Google Scholar] [CrossRef]
- Aviram, R.; Zaffryar-Eilot, S.; Melamed, S.; Ganesh, K.S.; Hasson, P. Novel intracellular activities of Lysyl oxidase in smooth muscle cells suggest a critical role in aneurysm prevention through cytoskeletal regulation. Department of Genetics and Developmental Biology, The Rappaport Faculty of Medicine and Research Institute, Technion–Israel Institute of Technology, Haifa, Israel. 2022; manuscript in preparation. [Google Scholar]
- Shao, J.; Lu, J.; Zhu, W.; Yu, H.; Jing, X.; Wang, Y.L.; Wang, X.; Wang, X.J. Derepression of LOXL4 Inhibits Liver Cancer Growth by Reactivating Compromised P53. Cell Death Differ. 2019, 26, 2237–2252. [Google Scholar] [CrossRef]
- Pobbati, A.V.; Hong, W. Emerging Roles of TEAD Transcription Factors and Its Coactivators in Cancers. Cancer Biol. Therapy 2013, 14, 390–398. [Google Scholar] [CrossRef]
- Joshi, S.; Davidson, G.; Le Gras, S.; Watanabe, S.; Braun, T.; Mengus, G.; Davidson, I. TEAD Transcription Factors Are Required for Normal Primary Myoblast Differentiation in Vitro and Muscle Regeneration in Vivo. PLoS Genet. 2017, 13, e1006600. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Fong, K.S.; Mercier, F.; Boyd, C.D.; Csiszar, K.; Hayashi, M. Comparative Immunocytochemical Localization of Lysyl Oxidase (LOX) and the Lysyl Oxidase-like (LOXL) Proteins: Changes in the Expression of LOXL during Development and Growth of Mouse Tissues. J. Mol. Histol. 2004, 35, 845–855. [Google Scholar] [CrossRef]
- Ma, L.; Huang, C.; Wang, X.J.; Xin, D.E.; Wang, L.S.; Zou, Q.C.; Zhang, Y.S.; Tan, M.D.; Wang, Y.M.; Zhao, T.C.; et al. Lysyl Oxidase 3 Is a Dual-Specificity Enzyme Involved in STAT3 Deacetylation and Deacetylimination Modulation. Mol. Cell 2017, 65, 296–309. [Google Scholar] [CrossRef]
- Li, C.; Sharma-Bhandari, A.; Seo, J.H.; Kim, Y. Lysyl Oxidase-Variant 2 (LOX-v2) Colocalizes with Promyelocytic Leukemia-Nuclear Bodies in the Nucleus. IUBMB Life 2020, 72, 2400–2408. [Google Scholar] [CrossRef]
- Peinado, H.; Moreno-Bueno, G.; Hardisson, D.; Pérez-Gómez, E.; Santos, V.; Mendiola, M.; De Diego, J.I.; Nistal, M.; Quintanilla, M.; Portillo, F.; et al. Lysyl Oxidase-like 2 as a New Poor Prognosis Marker of Squamous Cell Carcinomas. Cancer Res. 2008, 68, 4541–4550. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Nellaiappan, K.; Strassmaier, T.; Graham, L.; Thomas, K.M.; Kagan, H.M. Localization and Activity of Lysyl Oxidase within Nuclei of Fibrogenic Cells. Proc. Natl. Acad. Sci. USA 1997, 94, 12817–12822. [Google Scholar] [CrossRef]
- Zhong, Y.; Mahoney, R.C.; Khatun, Z.; Chen, H.H.; Nguyen, C.T.; Caravan, P.; Roberts, J.D. Lysyl Oxidase Regulation and Protein Aldehydes in the Injured Newborn Lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 322, 1. [Google Scholar] [CrossRef]
- Kenyon, K.; Contente, S.; Trackman, P.C.; Tang, J.; Kagan, H.M.; Friedman, R.M. Lysyl Oxidase and Rrg Messenger RNA. Science 1991, 253, 802. [Google Scholar] [CrossRef]
- Friedman, R.M.; Yeh, A.; Gutman, P.; Contente, S.; Kenyon, K. Reversion by Deletion of Transforming Oncogene Following Interferon-Beta and Retinoic Acid Treatment. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 1997, 17, 647–651. [Google Scholar] [CrossRef]
- Contente, S.; Yeh, T.J.A.; Friedman, R.M. Tumor Suppressive Effect of Lysyl Oxidase Proenzyme. Biochim. Biophys. Acta 2009, 1793, 1272–1278. [Google Scholar] [CrossRef]
- Mello, M.L.S.; Contente, S.; Vidal, B.C.; Planding, W.; Schenck, U. Modulation of Ras Transformation Affecting Chromatin Supraorganization as Assessed by Image Analysis. Exp. Cell Res. 1995, 220, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Kagan, H.M.; Williams, M.A.; Calaman, S.D.; Berkowitz, E.M. Histone H1 Is a Substrate for Lysyl Oxidase and Contains Endogenous Sodium Borotritide-Reducible Residues. Biochem. Biophys. Res. Commun. 1983, 115, 186–192. [Google Scholar] [CrossRef]
- Giampuzzi, M.; Oleggini, R.; Di Donato, A. Demonstration of in Vitro Interaction between Tumor Suppressor Lysyl Oxidase and Histones H1 and H2: Definition of the Regions Involved. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2003, 1647, 245–251. [Google Scholar] [CrossRef]
- Starcher, B.; Conrad, N.; Hinek, A.; Hill, C.H. Antibody Raised to AKAAAKAAAKA Sequence on Tropoelastin Recognizes Tropoelastin but Not Mature Crosslinked Elastin: A New Tool in Metabolic and Structural Studies of Elastogenesis. Connect. Tissue Res. 1999, 40, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Giampuzzi, M.; Oleggini, R.; Di Donato, A. Altered Adhesion Features and Signal Transduction in NRK-49F Cells Transformed by down-Regulation of Lysyl Oxidase. Biochim. Biophys. Acta 2003, 1647, 239–244. [Google Scholar] [CrossRef]
- Herranz, N.; Dave, N.; Millanes-Romero, A.; Pascual-Reguant, L.; Morey, L.; Diaz, V.M.; Lorenz-Fonfria, V.; Gutierrez-Gallego, R.; Jeronimo, C.; Iturbide, A.; et al. Lysyl Oxidase-like 2 (LOXL2) Oxidizes Trimethylated Lysine 4 in Histone H3. FEBS J. 2016, 283, 4263–4273. [Google Scholar] [CrossRef] [PubMed]
- Okkelman, I.A.; Sukaeva, A.Z.; Kirukhina, E.V.; Korneenko, T.V.; Pestov, N.B. Nuclear Translocation of Lysyl Oxidase Is Promoted by Interaction with Transcription Repressor P66β. Cell Tissue Res. 2014, 358, 481–489. [Google Scholar] [CrossRef]
- Fu, J.; Qin, L.; He, T.; Qin, J.; Hong, J.; Wong, J.; Liao, L.; Xu, J. The TWIST/Mi2/NuRD Protein Complex and Its Essential Role in Cancer Metastasis. Cell Res. 2011, 21, 275–289. [Google Scholar] [CrossRef]
- Martin, A.; Salvador, F.; Moreno-Bueno, G.; Floristán, A.; Ruiz-Herguido, C.; Cuevas, E.P.; Morales, S.; Santos, V.; Csiszar, K.; Dubus, P.; et al. Lysyl Oxidase-like 2 Represses Notch1 Expression in the Skin to Promote Squamous Cell Carcinoma Progression. EMBO J. 2015, 34, 1090–1109. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.E.; Weinberg, S.H.; Lemmon, C.A. Mechanochemical Signaling of the Extracellular Matrix in Epithelial-Mesenchymal Transition. Front. Cell Dev. Biol. 2019, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Mäki, J.M.; Rasanen, J.; Tikkanen, H.; Sormunen, R.; Makikallio, K.; Kivirikko, K.I.; Soininen, R. Inactivation of the Lysyl Oxidase Gene Lox Leads to Aortic Aneurysms, Cardiovascular Dysfunction, and Perinatal Death in Mice. Circulation 2002, 106, 2503–2509. [Google Scholar] [CrossRef]
- Wagenseil, J.E.; Mecham, R.P. Vascular Extracellular Matrix and Arterial Mechanics. Physiol. Rev. 2009, 89, 957–989. [Google Scholar] [CrossRef]
- Salvador, F.; Martin, A.; López-Menéndez, C.; Moreno-Bueno, G.; Santos, V.; Vázquez-Naharro, A.; Santamaria, P.G.; Morales, S.; Dubus, P.R.; Muinelo-Romay, L.; et al. Lysyl Oxidase-like Protein LOXL2 Promotes Lung Metastasis of Breast Cancer. Cancer Res. 2017, 77, 5846–5859. [Google Scholar] [CrossRef]
- Lucero, H.A.; Kagan, H.M. Lysyl Oxidase: An Oxidative Enzyme and Effector of Cell Function. Cell. Mol. Life Sci. 2006, 63, 2304–2316. [Google Scholar] [CrossRef]
- Mäki, J.M.; Kivirikko, K.I. Cloning and Characterization of a Fourth Human Lysyl Oxidase Isoenzyme. Biochem. J. 2001, 355, 381–387. [Google Scholar] [CrossRef]
- Nellaiappan, K.; Risitano, A.; Liu, G.; Nicklas, G.; Kagan, H.M. Fully Processed Lysyl Oxidase Catalyst Translocates from the Extracellular Space into Nuclei of Aortic Smooth-Muscle Cells. J. Cell. Biochem. 2000, 79, 576–582. [Google Scholar] [CrossRef]
- Vallet, S.D.; Berthollier, C.; Salza, R.; Muller, L.; Ricard-Blum, S. The Interactome of Cancer-Related Lysyl Oxidase and Lysyl Oxidase-Like Proteins. Cancers 2021, 13, 71. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaffryar-Eilot, S.; Hasson, P. Lysyl Oxidases: Orchestrators of Cellular Behavior and ECM Remodeling and Homeostasis. Int. J. Mol. Sci. 2022, 23, 11378. https://doi.org/10.3390/ijms231911378
Zaffryar-Eilot S, Hasson P. Lysyl Oxidases: Orchestrators of Cellular Behavior and ECM Remodeling and Homeostasis. International Journal of Molecular Sciences. 2022; 23(19):11378. https://doi.org/10.3390/ijms231911378
Chicago/Turabian StyleZaffryar-Eilot, Shelly, and Peleg Hasson. 2022. "Lysyl Oxidases: Orchestrators of Cellular Behavior and ECM Remodeling and Homeostasis" International Journal of Molecular Sciences 23, no. 19: 11378. https://doi.org/10.3390/ijms231911378
APA StyleZaffryar-Eilot, S., & Hasson, P. (2022). Lysyl Oxidases: Orchestrators of Cellular Behavior and ECM Remodeling and Homeostasis. International Journal of Molecular Sciences, 23(19), 11378. https://doi.org/10.3390/ijms231911378