
Effect of Neonicotinoid Pesticides on Japanese Water Systems: Review with Focus on Reproductive Toxicity

 , , ,
, , ,  ,
,
Abstract
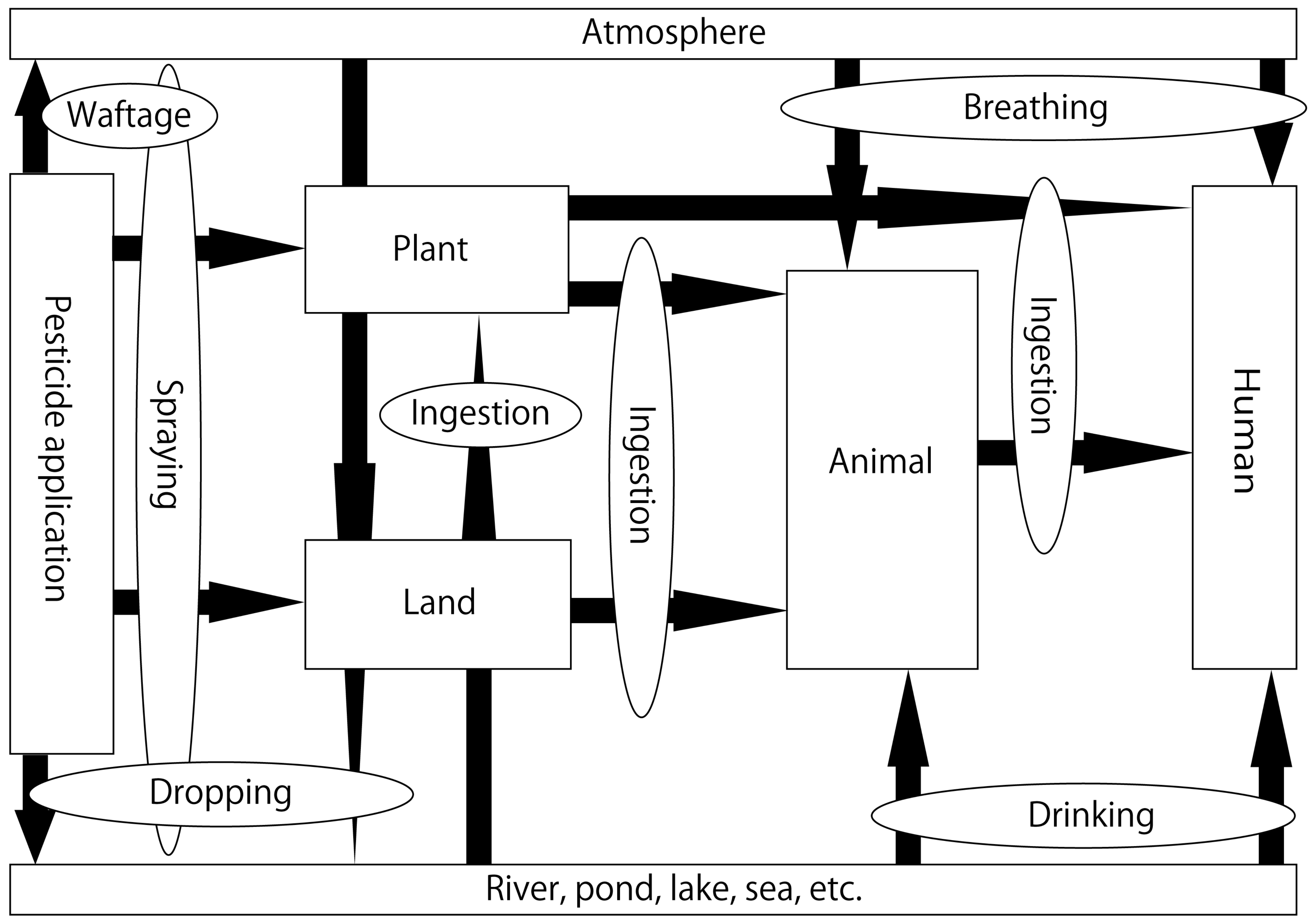
:1. Introduction
2. Literature Review
2.1. NPs
2.2. NPs in the Aquatic Environment
2.3. NP Concentrations in River Water in Japan
2.4. Safety of River Water in Japan
2.5. NPs and Endocrine Disruptors
2.6. Effects of NPs on the Testis
2.7. Effects of NPs on the Ovary
2.8. Mechanisms Underlying the Reproductive Toxicity of NPs
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NPs | neonicotinoid pesticides |
| DDT | dichlorodiphenyltrichloroethane |
| nAchR | nicotinic acetylcholine receptor |
| CYP | cytochrome P |
| ADI | acceptable daily intake |
| ARfD | acute reference dose |
| ED | endocrine disruptor |
| PCBs | polychlorinated biphenyl |
| LH | luteinizing hormone |
| NOAEL | no-observed-adverse-effect level |
| AOPP | advanced oxidation protein product |
| ATP | adenosine triphosphate |
| AR | androgen receptor |
| BAX | BCL2-associated X protein |
| cAMP | cyclic adenosine monophosphate |
| CAT | catalase |
| Cyp11a1 | cytochrome P450 family 11 subfamily A member 1 |
| Cyp17a1 | cytochrome P450 family 17 subfamily A |
| FSH | follicle-stimulating hormone |
| GPx | glutathione peroxidase |
| GPx4 | glutathione peroxidase 4 |
| GR | glutathione reductase |
| GSH | glutathione |
| GST | glutathione S-transferase |
| HMG-CoA | 3-hydroxy-3-methyl-glutaryl CoA synthase |
| HSD17b3 | 17β-hydroxysteroid dehydrogenase 3 |
| LDH-x | lactate dehydrogenase-x |
| LDL-C | low-density lipoprotein cholesterol |
| LPO | lipid peroxidation |
| MDA | malondialdehyde |
| NO | nitric oxide |
| NR5A1 | nuclear receptor family 5 group A member 1 |
| nAChRα4 | nicotinic acetylcholine receptor α4 |
| nAChRα7 | nicotinic acetylcholine receptor α7 |
| OGG1 | 8-oxoguanine glycosylase |
| PBR | peripheral benzodiazepine receptor |
| PCNA | proliferating cell nuclear antigen |
| P450scc | cytochrome P450 cholesterol side-chain cleavage enzyme |
| P45017a | cytochrome P450 family 17 subfamily A |
| ROS | reactive oxygen species |
| SDH | sorbitol dehydrogenase |
| SOD | superoxide dismutase |
| SR-B1 | scavenger receptor class B member 1 |
| StAR | steroidogenic acute regulatory protein |
| TAS | total antioxidant status |
| TC | total cholesterol |
| Top2a | DNA topoisomerase II alpha |
| TOS | total oxidant status |
| TTH | total thiol level |
| γ-GT | γ-glutamyltranspeptidase |
| 3β-HSD | 3β-hydroxysteroid dehydrogenase |
| 8-OHdG | 8-hydroxy-2′-deoxyguanosine |
| 17β-HSD | 17β-hydroxysteroid dehydrogenase |
| AOPP | advanced oxidation protein product |
| vitamin E | α-tocopherol |
| vitamin C | ascorbic acid |
References
- Iwafune, T.; Inao, K.; Horio, T.; Iwasaki, N.; Yokoyama, A.; Nagai, T. Behavior of paddy pesticides and major metabolites in the Sakura River, Ibaraki, Japan. J. Pestic. Sci. 2010, 35, 114–123. [Google Scholar] [CrossRef]
- Terayama, H.; Kiyoshima, D.; Hayashi, S.; Sakabe, K. Toxicity of neonicotinoid pesticides in the male reproductive organ. Agric. Biol. Publ. 2020, 4, 61–66. [Google Scholar]
- Ratcliffe, D.A. Decrease in eggshell weight in certain birds of prey. Nature 1967, 215, 208–210. [Google Scholar] [CrossRef] [PubMed]
- Newton, I.; Bogan, J.A. Organochlorine residues, eggshell thinning and hatching success in British sparrowhawks. Nature 1974, 249, 582–583. [Google Scholar] [CrossRef] [PubMed]
- Encarnação, T.; Santos, D.; Ferreira, S.; Valente, A.J.M.; Pereira, J.C.; Campos, M.G.; Burrows, H.D.; Pais, A.A.C.C. Removal of imidacloprid from water by microalgae Nannochloropsis sp. and its determination by a validated RP-HPLC method. Bull. Environ. Contam. Toxicol. 2021, 107, 131–139. [Google Scholar] [CrossRef]
- Ewere, E.E.; Reichelt-Brushett, A.; Benkendorff, K. Impacts of neonicotinoids on molluscs: What we know and what we need to know. Toxics 2021, 9, 21. [Google Scholar] [CrossRef]
- Tooker, J.F.; Pearsons, K.A. Newer characters, same story: Neonicotinoid insecticides disrupt food webs through direct and indirect effects. Curr. Opin. Insect Sci. 2021, 46, 50–56. [Google Scholar] [CrossRef]
- Shen, C.; Pan, X.; Wu, X.; Xu, J.; Dong, F.; Zheng, Y. Predicting and assessing the toxicity and ecological risk of seven widely used neonicotinoid insecticides and their aerobic transformation products to aquatic organisms. Sci. Total Environ. 2022, 847, 157670. [Google Scholar] [CrossRef]
- Ford, K.A.; Casida, J.E. Chloropyridinyl neonicotinoid insecticides: Diverse molecular substituents contribute to facile metabolism in mice. Chem. Res. Toxicol. 2006, 19, 944–951. [Google Scholar] [CrossRef]
- Ford, K.A.; Casida, J.E. Unique and common metabolites of thiamethoxam, clothianidin, and dinotefuran in mice. Chem. Res. Toxicol. 2006, 19, 1549–1556. [Google Scholar] [CrossRef]
- Blacquière, T.; Smagghe, G.; van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic insecticides (neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef]
- Taira, K. Health effects of neonicotinoid insecticides-Part 2: Pharmacology, application, regulation, and discussion. Jpn. J. Clin. Ecol. 2012, 21, 35–45. [Google Scholar]
- Terayama, H.; Endo, H.; Tsukamoto, H.; Matsumoto, K.; Umezu, M.; Kanazawa, T.; Ito, M.; Sato, T.; Naito, M.; Kawakami, S.; et al. Acetamiprid accumulates in different amounts in murine brain regions. Int. J. Environ. Res. Public Health 2016, 13, 937. [Google Scholar] [CrossRef]
- Burke, A.P.; Niibori, Y.; Terayama, H.; Ito, M.; Pidgeon, C.; Arsenault, J.; Camarero, P.R.; Cummins, C.L.; Mateo, R.; Sakabe, K.; et al. Mammalian susceptibility to a neonicotinoid insecticide after fetal and early postnatal exposure. Sci. Rep. 2018, 8, 16639. [Google Scholar] [CrossRef]
- Food Safety Commission of Japan, Fipronil. Available online: https://www.fsc.go.jp/fsciis/evaluationDocument/show/kya20151013447 (accessed on 2 September 2022).
- Food Safety Commission of Japan, Acetamiprid. Available online: https://www.fsc.go.jp/fsciis/evaluationDocument/show/kya20140702188 (accessed on 2 September 2022).
- Food Safety Commission of Japan, Imidacloprid. Available online: https://www.fsc.go.jp/fsciis/evaluationDocument/show/kya20151117469 (accessed on 2 September 2022).
- Ge, R.S.; Dong, Q.; Sottas, C.M.; Chen, H.; Zirkin, B.R.; Hardy, M.P. Gene expression in rat Leydig cells during development from the progenitor to adult stage: A cluster analysis. Biol. Reprod. 2005, 72, 1405–1415. [Google Scholar] [CrossRef]
- Schirmer, S.U.; Eckhardt, I.; Lau, H.; Klein, J.; DeGraaf, Y.C.; Lips, K.S.; Pineau, C.; Gibbins, I.L.; Kummer, W.; Meinhardt, A.; et al. The cholinergic system in rat testis is of non-neuronal origin. Reproduction 2011, 142, 157–166. [Google Scholar] [CrossRef]
- Fedotova, I.O. Influence of stimulation and blockade of α4β2 nicotinic acetylcholine receptors on learning of female rats in basic phases of ovary cycle. Ross. Fiziol. Zh. Im. I. M. Sechenova 2014, 100, 318–327. [Google Scholar]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A common pesticide decreases foraging success and survival in honey bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.J.; Ramos-Rodriguez, O.; Raine, N.E. Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature 2012, 491, 105–108. [Google Scholar] [CrossRef]
- Kawai, S. Aquatic Insects of Japan: Manual with Keys and Illustrations; Kawai, S., Tanida, K., Eds.; Tokai University Press: Tokyo, Japan, 2018; pp. 1–10. [Google Scholar]
- Karube, H.; Terayama, H.; Sakabe, K. Is the decline of Sympetrum maculatum in the Tonoh area in Gifu Prefecture caused by neonicotinoid insecticide? TOMBO 2019, 61, 1–7. [Google Scholar]
- Hayasaka, D.; Suzuki, K.; Korenaga, T.; Saito-Morooka, F.; Nomura, T.; Fukasawa, K.; Sánchez-Bayo, F.; Goka, K. Effects of two successive annual treatments of two systemic insecticides, imidacloprid and fipronil, on dragonfly nymph communities in experimental paddies. Jpn. J. Pestic. Sci. 2013, 38, 101–107. [Google Scholar] [CrossRef]
- Yamamuro, M.; Komuro, T.; Kamiya, H.; Kato, T.; Hasegawa, H.; Kameda, Y. Neonicotinoids disrupt aquatic food webs and decrease fishery yields. Science 2019, 366, 620–623. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Foppen, R.P.; van Turnhout, C.A.; de Kroon, H.; Jongejans, E. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 2014, 511, 341–343. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, P.J.; Van Smeden, J.M.; Bekele, R.S.; Dierick, W.; De Gelder, D.M.; Noteboom, M.; Roessink, I. Acute and chronic toxicity of neonicotinoids to nymphs of a mayfly species and some notes on seasonal differences. Environ. Toxicol. Chem. 2016, 35, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Uğurlu, P.; Ünlü, E.; Satar, E.İ. The toxicological effects of thiamethoxam on Gammarus kischineffensis (Schellenberg 1937) (Crustacea: Amphipoda). Environ. Toxicol. Pharmacol. 2015, 39, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Pisa, L.; Goulson, D.; Yang, E.C.; Gibbons, D.; Sánchez-Bayo, F.; Mitchell, E.; Aebi, A.; van der Sluijs, J.; MacQuarrie, C.J.K.; Giorio, C.; et al. An update of the Worldwide Integrated Assessment (WIA) on systemic insecticides. Part 2: Impacts on organisms and ecosystems. Environ. Sci. Pollut. Res. 2021, 28, 11749–11797. [Google Scholar] [CrossRef]
- Naoi, H.; Kamata, M. Evaluation for Neonicotinoid Pesticide in Water Environment and Water Purification Process. Bull. Inst. Sci. Technol. 2011, 39, 11–17. [Google Scholar]
- Sato, M.; Uemura, H.; Kosaka, K.; Asami, M.; Kamata, M. Survey of pesticide concentrations, including neonicotinoids, in the Sagami River, its tributaries and tap water. J. Jpn. Soc. Water Environ. 2016, 5, 153–162. [Google Scholar] [CrossRef]
- Umedu, M.; Terayama, H.; Ito, M.; Matumoto, K.; Tsukamoto, H.; Umemoto, K.; Qu, N.; Kawakami, S.; Fujino, Y.; Sakabe, K. Concentrations of neonicotinoid in the Kaname River system in Kanagawa Prefecture. Jpn. J. Clin. Ecol. 2019, 27, 47–55. [Google Scholar]
- Nakayama, S.; Mishima, S. Field survey of neonicotinoid pesticides and a related material in Kanagawa Prefecture. Bull. Kanagawa Environ. Res. Center 2019, 4216–4221. Available online: https://www.pref.kanagawa.jp/documents/60011/r01bull03.pdf (accessed on 22 September 2022).
- Takeuchi, Y.; Nishizawa, N. Survey of pesticide concentrations, including neonicotinoids, in Fukui Prefecture. Annu. Rep. Fukui Prefect. Inst. Public Health Environ. Sci. 2018, 71–75. Available online: http://www.erc.pref.fukui.jp/center/publish/report/2018/3-2-5.pdf (accessed on 28 July 2022).
- Nishino, K.; Kato, M.; Shimoma, S. Survey on the actual conditions of neonicotinoid pesticides in rivers in Tokyo. Tokyo Metrop. Inst. Environ. Sci. Annu. Rep. 2018, 64–65. Available online: https://dl.ndl.go.jp/info:ndljp/pid/11655321?tocOpened=1 (accessed on 28 July 2022).
- Terayama, H.; Kiyoshima, D.; Karube, H.; Fujino, Y.; Ito, M.; Matsumoto, K.; Kawakami, S.; Sato, T.; Sakabe, K. Neonicotinoid pesticides in river water. Jpn. J. Clin. Ecol. 2020, 29, 1–9. [Google Scholar]
- Japan Plant Protection Association. Noyakuyoran (Agrichemical Directory); Japan Plant Protection Association: Tokyo, Japan, 2017; pp. 96–456. [Google Scholar]
- Ministry of the Environment. Agricultural Chemical Registration Standards for Conservation of Water, Soil, Ground, and Marine Environment, and Prevention of Damage to Animals and Plants in the Living Environment of Water Areas. Available online: http://www.env.go.jp/water/sui-kaitei/kijun.html#list01-a (accessed on 11 May 2022).
- Ministry of the Environment. Pesticide Registration Standards for Water/Soil/Ground/Marine Environment Conservation and Water Pollution. Available online: https://www.env.go.jp/water/dojo/noyaku/odaku_kijun/kijun.html (accessed on 11 May 2022).
- Food Safety Commission of Japan, Cabinet Office, Pesticides. Available online: http://www.fsc.go.jp/fsciis/evaluationDocument/list?itemCategory=001 (accessed on 11 May 2022).
- Oyama, K.; Yabuki, Y.; Banno, A. Investigation of seasonal changes and ecological risk assessments of neonicotinoid pesticides in rivers in Osaka, Japan. Mizu Kankyo Gakkaishi 2019, 42, 277–284. [Google Scholar] [CrossRef]
- Cavallaro, M.C.; Morrissey, C.A.; Headley, J.V.; Peru, K.M.; Liber, K. Comparative chronic toxicity of Imidacloprid, clothianidin, and thiamethoxam to Chironomus dilutus and estimation of toxic equivalency factors. Environ. Toxicol. Chem. 2017, 36, 372–382. [Google Scholar] [CrossRef]
- World Health Organization/Institute of Peace and Conflict Studies. Global Assessment of the State-of-the-Science of Endocrine Disruptors; World Health Organisation: Geneva, Switzerland, 2002. [Google Scholar]
- Ito, T.; Yamazaki, S.; Ota, K.; Ohmura, M.; Oyadomari, S. Identification of a novel protein binding to the endocrine disruptor nonylphenol. Nippon Eiyo Shokuryo Gakkaishi 2015, 68, 63–68. [Google Scholar] [CrossRef]
- Reiter, L.W.; DeRosa, C.; Kavlock, R.J.; Lucier, G.; Mac, M.J.; Melillo, J.; Melnick, R.L.; Sinks, T.; Walton, B.T. The U.S. federal framework for research on endocrine disruptors and an analysis of research programs supported during fiscal year 1996. Environ. Health Perspec. 1998, 106, 105–113. [Google Scholar] [CrossRef]
- Chemsec SIN LIST. Endocrine Disrupting Chemicals. Available online: https://sinlist.chemsec.org/endocrine-disruptors/ (accessed on 11 May 2022).
- TEDX the Endocrine Disruption Exchange. Available online: https://endocrinedisruption.org/interactive-tools/tedx-list-of-potential-endocrine-disruptors/search-the-tedx-list (accessed on 11 May 2022).
- Beausoleil, C.; Emond, C.; Cravedi, J.P.; Antignac, J.P.; Applanat, M.; Appenzeller, B.R.; Beaudouin, R.; Belzunces, L.P.; Canivenc-Lavier, M.C.; Chevalier, N.; et al. Regulatory identification of BPA as an endocrine disruptor: Context and methodology. Mol. Cell. Endocrinol. 2018, 475, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Mnif, W.; Hassine, A.I.; Bouaziz, A.; Bartegi, A.; Thomas, O.; Roig, B. Effect of endocrine disruptor pesticides: A review. Int. J. Environ. Res. Public Health 2011, 8, 2265–2303. [Google Scholar] [CrossRef]
- Ewence, A.; Brescia, S.; Johnson, I.; Rumsby, P.C. An approach to the identification and regulation of endocrine disrupting pesticides. Food Chem. Toxicol. 2015, 78, 214–220. [Google Scholar] [CrossRef]
- Kabir, E.R.; Rahman, M.S.; Rahman, I. A review on endocrine disruptors and their possible impacts on human health. Environ. Toxicol. Pharmacol. 2015, 40, 241–258. [Google Scholar] [CrossRef] [PubMed]
- European Chemicals Agency. Candidate List of Substances of Very High Concern for Authorization. Available online: https://echa.europa.eu/candidate-list-table (accessed on 29 September 2022).
- Hirano, T.; Yanai, S.; Omotehara, T.; Hashimoto, R.; Umemura, Y.; Kubota, N.; Minami, K.; Nagahara, D.; Matsuo, E.; Aihara, Y.; et al. The combined effect of clothianidin and environmental stress on the behavioral and reproductive function in male mice. J. Vet. Med. Sci. 2015, 77, 1207–1215. [Google Scholar] [CrossRef]
- Terayama, H.; Qu, N.; Endo, H.; Ito, M.; Tsukamoto, H.; Umemoto, K.; Kawakami, S.; Fujino, Y.; Tatemichi, M.; Sakabe, K. Effect of acetamiprid on the immature murine testes. Int. J. Environ. Health Res. 2018, 28, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Bal, R.; Türk, G.; Tuzcu, M.; Yilmaz, O.; Kuloglu, T.; Gundogdu, R.; Gür, S.; Agca, A.; Ulas, M.; Cambay, Z.; et al. Assessment of imidacloprid toxicity on reproductive organ system of adult male rats. J. Environ. Sci. Health B 2012, 47, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Bal, R.; Naziroğlu, M.; Türk, G.; Yilmaz, Ö.; Kuloğlu, T.; Etem, E.; Baydas, G. Insecticide imidacloprid induces morphological and DNA damage through oxidative toxicity on the reproductive organs of developing male rats. Cell Biochem. Funct. 2012, 30, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.; Mohamed, W.A.M.; Khater, S.I. Imidacloprid induces various toxicological effects related to the expression of 3β-HSD, NR5A1, and OGG1 genes in mature and immature rats. Environ. Pollut. 2017, 221, 15–25. [Google Scholar] [CrossRef]
- Lonare, M.; Kumar, M.; Raut, S.; More, A.; Doltade, S.; Badgujar, P.; Telang, A. Evaluation of ameliorative effect of curcumin on imidacloprid-induced male reproductive toxicity in Wistar rats. Environ. Toxicol. 2016, 31, 1250–1263. [Google Scholar] [CrossRef] [PubMed]
- Bal, R.; Türk, G.; Yılmaz, Ö.; Etem, E.; Kuloğlu, T.; Baydaş, G.; Naziroğlu, M. Effects of clothianidin exposure on sperm quality, testicular apoptosis and fatty acid composition in developing male rats. Cell Biol. Toxicol. 2012, 28, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.; Zhang, J.; Hou, X.; Zhang, S.; Tan, J.; Chen, Y.; Yang, W.; Zeng, J.; Han, Y.; Liu, X.; et al. Acetamiprid inhibits testosterone synthesis by affecting the mitochondrial function and cytoplasmic adenosine triphosphate production in rat Leydig cells. Biol. Reprod. 2017, 96, 254–265. [Google Scholar] [CrossRef]
- Yuan, X.; Shen, J.; Zhang, X.; Tu, W.; Fu, Z.; Jin, Y. Imidacloprid disrupts the endocrine system by interacting with androgen receptor in male mice. Sci. Total Environ. 2020, 708, 135163. [Google Scholar] [CrossRef] [PubMed]
- Kammoun, I.; Bkhairia, I.; Ben Abdallah, F.; Jaballi, I.; Ktari, N.; Boudawara, O.; Nasri, M.; Gharsallah, N.; Hakim, A.; Ben Amara, I. Potential protective effects of polysaccharide extracted from Ulva lactuca against male reprotoxicity induced by thiacloprid. Arch. Physiol. Biochem. 2017, 123, 334–343. [Google Scholar] [CrossRef]
- Yanai, S.; Hirano, T.; Omotehara, T.; Takada, T.; Yoneda, N.; Kubota, N.; Yamamoto, A.; Mantani, Y.; Yokoyama, T.; Kitagawa, H.; et al. Prenatal and early postnatal NOAEL-dose clothianidin exposure leads to a reduction of germ cells in juvenile male mice. J. Vet. Med. Sci. 2017, 79, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Arıcan, E.Y.; Gökçeoğlu Kayalı, D.; Ulus Karaca, B.; Boran, T.; Öztürk, N.; Okyar, A.; Ercan, F.; Özhan, G. Reproductive effects of subchronic exposure to acetamiprid in male rats. Sci. Rep. 2020, 10, 8985. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, U.; Srivastava, M.K.; Srivastava, L.P. Toxicological impact of technical imidacloprid on ovarian morphology, hormones and antioxidant enzymes in female rats. Food Chem. Toxicol. 2011, 49, 3086–3089. [Google Scholar] [CrossRef] [PubMed]
- Mzid, M.; Ghlissi, Z.; Salem, M.B.; Khedir, S.B.; Chaabouni, K.; Ayedi, F.; Sahnoun, Z.; Hakim, A.; Rebai, T. Chemoprotective role of ethanol extract of Urtica urens L. against the toxicity of imidacloprid on endocrine disruption and ovarian morphometric in female rats, GC/MS analysis. Biomed. Pharmacother. 2018, 97, 518–527. [Google Scholar] [CrossRef]
- Kitauchi, S.; Maeda, M.; Hirano, T.; Ikenaka, Y.; Nishi, M.; Shoda, A.; Murata, M.; Mantani, Y.; Yokoyama, T.; Tabuchi, Y.; et al. Effects of in utero and lactational exposure to the no-observed-adverse-effect level (NOAEL) dose of the neonicotinoid clothianidin on the reproductive organs of female mice. J. Vet. Med. Sci. 2021, 83, 746–753. [Google Scholar] [CrossRef]
- Mikolić, A.; Karačonji, I.B. Imidacloprid as reproductive toxicant and endocrine disruptor: Investigations in laboratory animals. Arh. Hig. Rada. Toksikol. 2018, 69, 103–108. [Google Scholar] [CrossRef]
- Bal, R.; Türk, G.; Tuzcu, M.; Yılmaz, Ö.; Kuloğlu, T.; Baydaş, G.; Naziroğlu, M.; Yener, Z.; Etem, E.; Tuzcu, Z. Effects of the neonicotinoid insecticide, clothianidin, on the reproductive organ system in adult male rats. Drug Chem. Toxicol. 2013, 36, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, L.; Verma, P.K.; Raina, R.; Sood, S. Potentiating effect of imidacloprid on arsenic-induced testicular toxicity in Wistar rats. BMC Pharmacol. Toxicol. 2018, 19, 48. [Google Scholar] [CrossRef]
- Pandey, S.P.; Tsutsui, K.; Mohanty, B. Endocrine disrupting pesticides impair the neuroendocrine regulation of reproductive behaviors and secondary sexual characters of red munia (Amandava amandava). Physiol. Behav. 2017, 173, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.; Pandey, S.P.; Tsutsui, K. Thyroid disrupting pesticides impair the hypothalamic-pituitary-testicular axis of a wildlife bird, Amandava Amandava. Reprod Toxicol. 2017, 71, 32–41. [Google Scholar] [CrossRef]
- Baines, D.; Wilton, E.; Pawluk, A.; de Gorter, M.; Chomistek, N. Neonicotinoids act like endocrine disrupting chemicals in newly-emerged bees and winter bees. Sci. Rep. 2017, 7, 10979. [Google Scholar] [CrossRef] [PubMed]
- Caron-Beaudoin, E.; Viau, R.; Hudon-Thibeault, A.A.; Vaillancourt, C.; Sanderson, J.T. The use of a unique co-culture model of fetoplacental steroidogenesis as a screening tool for endocrine disruptors: The effects of neonicotinoids on aromatase activity and hormone production. Toxicol. Appl. Pharmacol. 2017, 332, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Duzguner, V.; Erdogan, S. Chronic exposure to imidacloprid induces inflammation and oxidative stress in the liver & central nervous system of rats. Pestic. Biochem. Physiol. 2012, 104, 58–64. [Google Scholar] [CrossRef]
- Xu, X.; Wang, X.; Yang, Y.; Ares, I.; Martínez, M.; Lopez-Torres, B.; Martínez-Larrañaga, M.R.; Wang, X.; Anadón, A.; Martinez, M.A. Neonicotinoids: Mechanisms of systemic toxicity based on oxidative stress-mitochondrial damage. Arch. Toxicol. 2022, 96, 1493–1520. [Google Scholar] [CrossRef]
- Hirano, T.; Minagawa, S.; Furusawa, Y.; Yunoki, T.; Ikenaka, Y.; Yokoyama, T.; Hoshi, N.; Tabuchi, Y. Growth and neurite stimulating effects of the neonicotinoid pesticide clothianidin on human neuroblastoma SH-SY5Y cells. Toxicol. Appl. Pharmacol. 2019, 383, 114777. [Google Scholar] [CrossRef]
- Martelli, F.; Zhongyuan, Z.; Wang, J.; Wong, C.O.; Karagas, N.E.; Roessner, U.; Rupasinghe, T.; Venkatachalam, K.; Perry, T.; Bellen, H.J.; et al. Low doses of the neonicotinoid insecticide imidacloprid induce ROS triggering neurological and metabolic impairments in Drosophila. Proc. Natl. Acad. Sci. USA 2020, 117, 25840–25850. [Google Scholar] [CrossRef]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar] [CrossRef] [Green Version]
- Food Safety Commission of Japan, Pesticides. Available online: https://www.mhlw.go.jp/file/06-Seisakujouhou-11130500-Shokuhinanzenbu/H27.5.19.pdf (accessed on 11 May 2022).


{kind=link}
| Region | Sampling Point | Time of Water Sampling | Compound | Maximum Concentration (mg/L) | Sampling Month with Maximum Concentration | Reference |
|---|---|---|---|---|---|---|
| Kanagawa | Upstream–downstream of the Tsurumi River | May–December 2009 | Imidacloprid | 0.42 | July | [33] |
| Acetamiprid | 0.06 | June | ||||
| Midstream–downstream of the Sagami River | Late April 2014–Mid-March 2015 | Imidacloprid | 0.104 | June | [34] | |
| Acetamiprid | 0.023 | - 1 | ||||
| Thiacloprid | 0.002 | - | ||||
| Thiamethoxam | 0.202 | - | ||||
| Dinotefuran | 0.048 | - | ||||
| Clothianidin | 0.085 | June | ||||
| Upstream–downstream of the Kaname River | April–December 2017 | Imidacloprid | 0.836 | June | [35] | |
| Acetamiprid | 0.779 | July | ||||
| Thiacloprid | 0.006 | November | ||||
| Thiamethoxam | 0.029 | November | ||||
| Nitenpyram | N.D. 2 | - | ||||
| Dinotefuran | 0.373 | August | ||||
| Clothianidin | 0.482 | May | ||||
| Midstream–downstream of the Kaname River * | April–August, November 2018, March 2019 | Imidacloprid | 0.095 | June | [36] | |
| Acetamiprid | 0.004 | June | ||||
| Thiacloprid | Unclear | - | ||||
| Thiamethoxam | 0.011 | May | ||||
| Nitenpyram | Unclear | - | ||||
| Dinotefuran | 0.043 | August | ||||
| Clothianidin | 0.053 | June | ||||
| Fipronil | 0.037 | June | ||||
| Fukui | Downstream of the Kuzuryu River | April–November 2018 | Imidacloprid | 0.055 | May | [37] |
| Acetamiprid | 0.0012 | August | ||||
| Thiacloprid | 0.0012 | - | ||||
| Thiamethoxam | 0.076 | May | ||||
| Nitenpyram | N.D. | - | ||||
| Dinotefuran | 0.27 | August | ||||
| Clothianidin | 0.13 | August | ||||
| Fipronil | 0.0045 | - | ||||
| Tokyo | Midstream of the Tama River | April–May 2017 | Imidacloprid | 0.0084 | - | [38] |
| Acetamiprid | 0.00094 | - | ||||
| Thiacloprid | 0.00045 | - | ||||
| Thiamethoxam | 0.0037 | - | ||||
| Nitenpyram | N.D. | - | ||||
| Dinotefuran | 0.0089 | - | ||||
| Clothianidin | 0.047 | - | ||||
| Fipronil | 0.0018 | - |
| Standard Type | Standard Name | Imidacloprid | Acetamiprid | Thiacloprid | Thiamethoxam | Nitenpyram | Dinotefuran | Clothianidin | Fipronil | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Environmental standard (μg/L) | Registration withholding standards for agricultural chemicals with reference to prevention of toxicity to aquatic plants and animals | 1.9 | 2.5 | 3.6 | 3.5 | 11 | 12 | 2.8 | 0.024 | [43] |
| Registration withholding standards for agricultural chemicals with reference to prevention of water pollution | 150 | 180 | 31 | 47 | 1400 | 580 | 250 | 0.5 | ||
| Food standard (μg/kg) | Acceptable daily intake (ADI) | 57 | 71 | 12 | 18 | 530 | 220 | 97 | 0.19 | |
| Acute reference dose (ARfD) | 100 | 100 | 31 | 500 | 600 | 1200 | 600 | 20 |
| Subject | Target Animal | Age at Initial Exposure | Dose (Per Day) | Exposure Period | Administration Method | Conclusion | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Testis Weight | Spermatogenesis | Androgen | Oxidative Stress | Cell Death | Other | |||||||
| Imidacloprid | Wistar albino rat | 8–9 weeks | 0.5, 2, 8 mg/kg | 90 days | Oral (gavage) | N.S. 1 | •Decreased epididymal sperm concentration (2, 8 mg/kg) and sperm motility (8 mg/kg) •Increased apoptosis indexes in the germinal epithelium of the testis, fragmentation of seminal DNA, and abnormal sperm rate (head) (8 mg/kg). | •Decreased serum testosterone (8 mg/kg) | •Decreased GSH in the testis (8 mg/kg) | - | •Decreased relative weights of epididymis, right cauda epididymis, and seminal vesicles (8 mg/kg) •Increased levels of oleic acid, linoleic acid (2, 8 mg/kg), stearic acid, and arachidonic acid in the testis (8 mg/kg) | [58] |
| Wistar albino rat | 7 days | 0.5, 2, 8 mg/kg | 90 days | Oral (gavage) | N.S. | •Decreased epididymal sperm concentration (8 mg/kg) •Increased proportion of abnormal sperm (head, tail, and total values) (8 mg/kg). | •Decreased serum testosterone (0.5, 2, 8 mg/kg) | •Increased MDA and decreased GSH in the testis (8 mg/kg) | - | •Increased testicular levels of palmitic, palmitoleic, stearic, oleic, linoleic, dihomo-γ-linolenic, arachidonic, docosapentaenoic acids as well as total lipid values (0.5, 2, 8 mg/kg). | [59] | |
| Wistar albino rat | 8 weeks | 45, 90 mg/kg | 28 days | Oral (gavage) | N.S. | •Decreased total epididymal sperm count, sperm motility, and live sperm count (45, 90 mg/kg) •Increased proportion of sperm with abnormal head or tail morphology (45, 90 mg/kg) •Increased proportion of abnormal sperm (45, 90 mg/kg) •Increased γ-GT activity, LDH-x, SDH in sperm suspension (45, 90 mg/kg) | •Decreased testicular 3β-HSD, 17β-HSD, and testosterone and serum testosterone (45, 90 mg/kg) | •Decreased protein concentration, reduced GSH, SOD, and GPx activity (45, 90 mg/kg) and CAT and GST activity in the testis (90 mg/kg) •Increased LPO and percent ROS-positive cells in a dose-dependent manner in the testis (45, 90 mg/kg) | - | •Decreased weight of epididymis (45, 90 mg/kg) | [61] | |
| Sprague–Dawley rat | 4 weeks and 7 weeks | 1 mg/kg | 65 days | Oral (- 3) | Decreased(4 and 7 weeks) | •Decreased epididymal sperm concentration, motility, intact seminal DNA percentage, and viable sperm percentage (4 and 7 weeks). | •Decreased serum testosterone, serum LH, serum estradiol, 3β-HSD mRNA, and NR5A1 mRNA (4 and 7 weeks). | •Increased 8-OHdG and OGG1 mRNA (4 and 7 weeks) | •Intense BAX-positive immunolabeling of the spermatogonial cell cytoplasm with moderate signals in the remaining spermatozoa (7 weeks)•Intense BAX-positive immunolabeling of the spermatogonial cell cytoplasm (3 weeks) | - | [60] | |
| Wistar rat | 12–14 weeks | 16.9 mg/kg | 28 days | Oral (-) | N.D. 2 | - | - | •Decreased GST, GR, GPx, CAT, and SOD activity and mean TTH in the testis •Increased testicular AOPP and MDA levels | - | - | [73] | |
| ICR mouse | 6 weeks | 3, 10, 30 mg/L | 70 days | Oral (-) | Decreased(30 mg/L) | • Decreased seminiferous tubule score determined by the Johnson scoring method in the testis (30 mg/L) | •Decreased serum testosterone, estradiol, and aromatase; testicular TC (30 mg/L) and serum LDL-C (10, 30 mg/L)•Decreased SR-B1 (3 mg/L), StAR, PBR, AR (10, 30 mg/L), P450scc, 3β-HSD (3, 10, 30 mg/L), P45017a, 17β-HSD, and HMG-CoA (30 mg/L) mRNA | - | - | •Imidacloprid showed binding affinity to the androgen receptor | [64] | |
| Thiacloprid | Wistar albino rat | Adult | 22.5 mg/kg | 30 days | Oral (gavage) | N.S. | •Decreased epididymal sperm concentration, sperm motility, sperm viability, and testicular DNA contents •Increased proportion of abnormal sperm | - | •Decreased SOD, CAT, and GPx activity and SOD, CAT, and GPx mRNA in the testis•Increased GSH, MDA, and AOPP in the testis | •Smear (a hallmark of necrosis) without ladder formation on agarose gel by the DNA fragmentation method in the testis | - | [65] |
| Clothianidin | Wistar albino rat | 7 days | 2, 8, 32 mg/kg | 90 days | Oral (gavage) | N.S. | •Decreased epididymal sperm concentration (32 mg/kg) •Increased proportion of sperm with abnormal head or tail morphology (8 and 32 mg/kg) •Increased proportion of abnormal sperm (8 and 32 mg/kg) | •Decreased serum testosterone protein (32 mg/kg) | •Decreased GSH (32 mg/kg) | •Increased apoptotic index in the germinal epithelium of the testis and fragmentation of the seminal DNA (32 mg/kg) | •Increased palmitic acid (8 mg/kg), arachidonic acid, docosapentaenoic acid (8, 32 mg/kg), testicular tissue cholesterol level (32 mg/kg), palmitoleic acid, and total lipid values (2, 8, 32 mg/kg) in the testis | [62] |
| Wistar albino rat | 8–9 weeks | 2, 8, 24 mg/kg | 90 days | Oral (gavage) | N.S. | N.S. | N.S. | - | - | •Decreased relative weights of epididymis (2, 8, 24 mg/kg), right cauda epididymis (8 mg/kg), and seminal vesicles (2, 8 mg/kg) •Increased LPO (measured as TBARS level), palmitic acid, linoleic acid, arachidonic acid, and cholesterol levels in the testis (2, 8, 24 mg/kg) | [72] | |
| C57BL/6NCrSlc Mouse | 8 weeks | 10, 50, 250 mg/kg | 28 days | Oral (gel intake) | N.S. | •Abnormal GPx4 immunoreactivity in Sertoli cells of the seminiferous tubules that also showed marked degeneration (250 mg/kg) | - | - | - | - | [56] | |
| C57BL/6NCrSlc Mouse(Male offspring postnatally on day 14) | Gestational day 1 | 10, 50 mg/kg | Approximately 30 days | Oral (gel intake) | N.S. | •Decreased number of germ cells per seminiferous tubule (50 mg/kg) | N.S. | - | - | - | [66] | |
| Acetamiprid | Sprague–Dawley rat | 5 weeks | 10, 30 mg/kg | 35 days | Oral (gavage) | Decreased(10, 30 mg/kg) | •Decreased number of spermatids and epididymal sperm (10, 30 mg/kg) | •Decreased plasma testosterone (30 mg/kg), Leydig cell number, StAR mRNA, Cyp11a1 mRNA, and 3β-HSD mRNA (10, 30 mg/kg) in the testis, cAMP (30 mg/kg), and ATP in the Leydig cells (10, 30 mg/kg)•Increased plasma LH (30 mg/kg), MDA, and NO in the Leydig cells (10, 30 mg/kg) | - | - | - | [63] |
| A/J mouse | 3 weeks | Average 2.6 and 21.4 mg/mouse | 180 days | Oral (through water intake) | N.S. | •Decreased Ki-67 (21.4 mg) and Top2a (2.6 and 21.4 mg) mRNA | •Decreased LH receptor, StAR, Cyp11a1, HSD17b3 (21.4 mg), and Cyp17a1mRNA (2.6 and 21.4 mg) in the testis | - | - | •Acetamiprid concentrations detected: 63.9 pg/mL in the serum and 7.1 pg/mL in the testis (21.4 mg) •Decreased nAChRα4 (21.4 mg) and nAChRα7 mRNA (2.6 and 21.4 mg) | [57] | |
| Sprague–Dawley rat | 8–10 weeks | 12.5, 25, 35 mg/kg | 90 days | Oral (gavage) | N.S. | •Decreased epididymal sperm concentration (25, 35 mg/kg) and seminiferous tubule score determined by the modified Johnson scoring method and proliferation index determined by the PCNA method in the testicular tissue (12.5, 25, 35 mg/kg). •Increased inhibin B (25 mg/kg) in the plasma | •Decreased cholesterol (25, 35 mg/kg) in the plasma •Increased FSH (12.5, 25, 35 mg/kg) and LH (12.5, 25 mg/kg) in the plasma | •Decreased GSH and TAS in the plasma and GSH and TAS in the testis (12.5, 25, 35 mg/kg) •Increased MDA (12.5, 25, 35 mg/kg) and TOS (35 mg/kg) in the plasma and P8MDA (25, 35 mg/kg) and TOS (35 mg/kg) in the testis | • Increased apoptotic index determined by the TUNEL method in the testicular tissue (12.5, 25, 35 mg/kg) | - | [67] | |
| Subject | Target Animal | Age at Initial Exposure | Dose (Per Day) | Exposure Period | Administration Method | Conclusion | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ovary Weight | Oogenesis | Female Hormones | Oxidative Stress | Cell Death | Other | |||||||
| Imidacloprid | Rattus norvigicus Wistar strain | 7 days | 5, 10, 20 mg/kg | 90 days | oral (gavage) | Decreased (20 mg/kg) | - 1 | •Increased serum FSH (20 mg/kg) •Decreased serum LH and progesterone (20 mg/kg) | •Increased LPO in the ovary (20 mg/kg) •Decreased GSH content, SOD, CAT, and GPX activity in the ovary (20 mg/kg) | - | - | [68] |
| Rat | 10–12 weeks | 50, 200, 300 mg/kg | 60 days | oral (gavage) | Decreased (50, 200, 300 mg/kg) | •Increased relative volume of atretic follicles against total follicles (50, 200, 300 mg/kg) •Increased follicular diameters at the primary (200, 300 mg/kg), secondary (200, 300 mg/kg), tertiary (50, 200, 300 mg/kg), and pre-ovulatory (50, 200, 300 mg/kg) stages | •Decreased serum 17β-estradiol (50, 200, 300 mg/kg) | •Increased ovarian AOPP and MDA (50, 200, 300 mg/kg) •Decreased vitamin E, GSH, SOD, CAT, and GPX activity in the ovary (50, 200, 300 mg/kg) | - | - | [69] | |
| Clothianidin | C57BL/6NCrSlc Mouse (postnatal female offspring at 3 weeks and 10 weeks) | Gestational period day 1.5 | 65 mg/kg | The dams (pregnant mouse) were given soft gel with or without CLO from gestational day 1.5 to postnatal day 21 period. | oral (gel intake) | Decreased (3 weeks) | N.S. 2 | •Decreased 17-hydroxyprogesterone (17-OH progesterone) and corticosterone in the blood (10 weeks) •Decreased activation of genes in the steroid hormone biosynthesis pathway in the ovary (3 weeks) | •Decreased GPx4 immunoreactivity in the ovary (3 weeks and 10 weeks) | - | - | [70] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terayama, H.; Sakabe, K.; Kiyoshima, D.; Qu, N.; Sato, T.; Suyama, K.; Hayashi, S.; Sakurai, K.; Todaka, E.; Mori, C. Effect of Neonicotinoid Pesticides on Japanese Water Systems: Review with Focus on Reproductive Toxicity. Int. J. Mol. Sci. 2022, 23, 11567. https://doi.org/10.3390/ijms231911567
Terayama H, Sakabe K, Kiyoshima D, Qu N, Sato T, Suyama K, Hayashi S, Sakurai K, Todaka E, Mori C. Effect of Neonicotinoid Pesticides on Japanese Water Systems: Review with Focus on Reproductive Toxicity. International Journal of Molecular Sciences. 2022; 23(19):11567. https://doi.org/10.3390/ijms231911567
Chicago/Turabian StyleTerayama, Hayato, Kou Sakabe, Daisuke Kiyoshima, Ning Qu, Tsutomu Sato, Kaori Suyama, Shogo Hayashi, Kenichi Sakurai, Emiko Todaka, and Chisato Mori. 2022. "Effect of Neonicotinoid Pesticides on Japanese Water Systems: Review with Focus on Reproductive Toxicity" International Journal of Molecular Sciences 23, no. 19: 11567. https://doi.org/10.3390/ijms231911567
APA StyleTerayama, H., Sakabe, K., Kiyoshima, D., Qu, N., Sato, T., Suyama, K., Hayashi, S., Sakurai, K., Todaka, E., & Mori, C. (2022). Effect of Neonicotinoid Pesticides on Japanese Water Systems: Review with Focus on Reproductive Toxicity. International Journal of Molecular Sciences, 23(19), 11567. https://doi.org/10.3390/ijms231911567