

Facilitation of Insulin Effects by Ranolazine in Astrocytes in Primary Culture
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
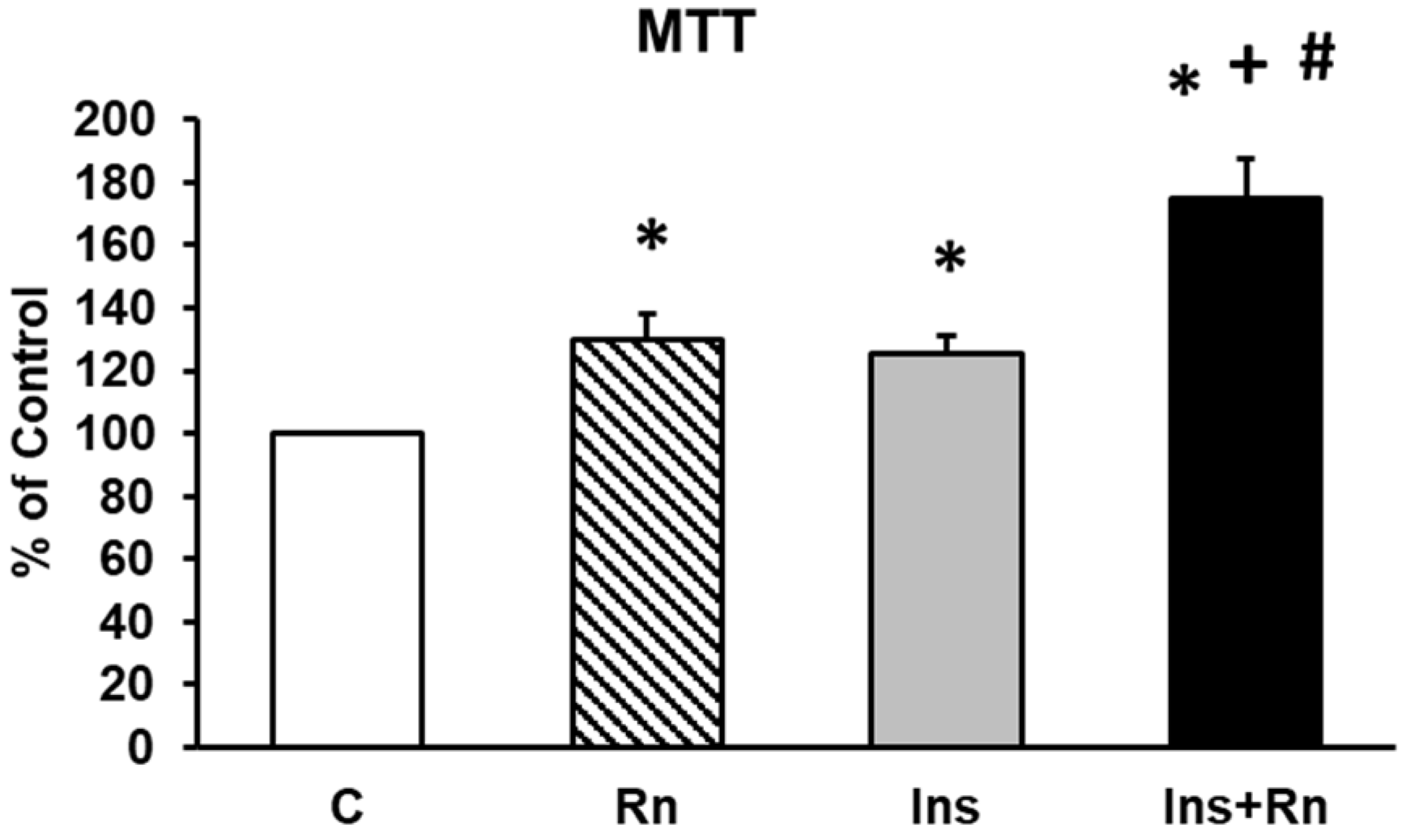
2.1. Cell Viability
2.2. Cell Proliferation
2.3. Protein Expression of p-AKT
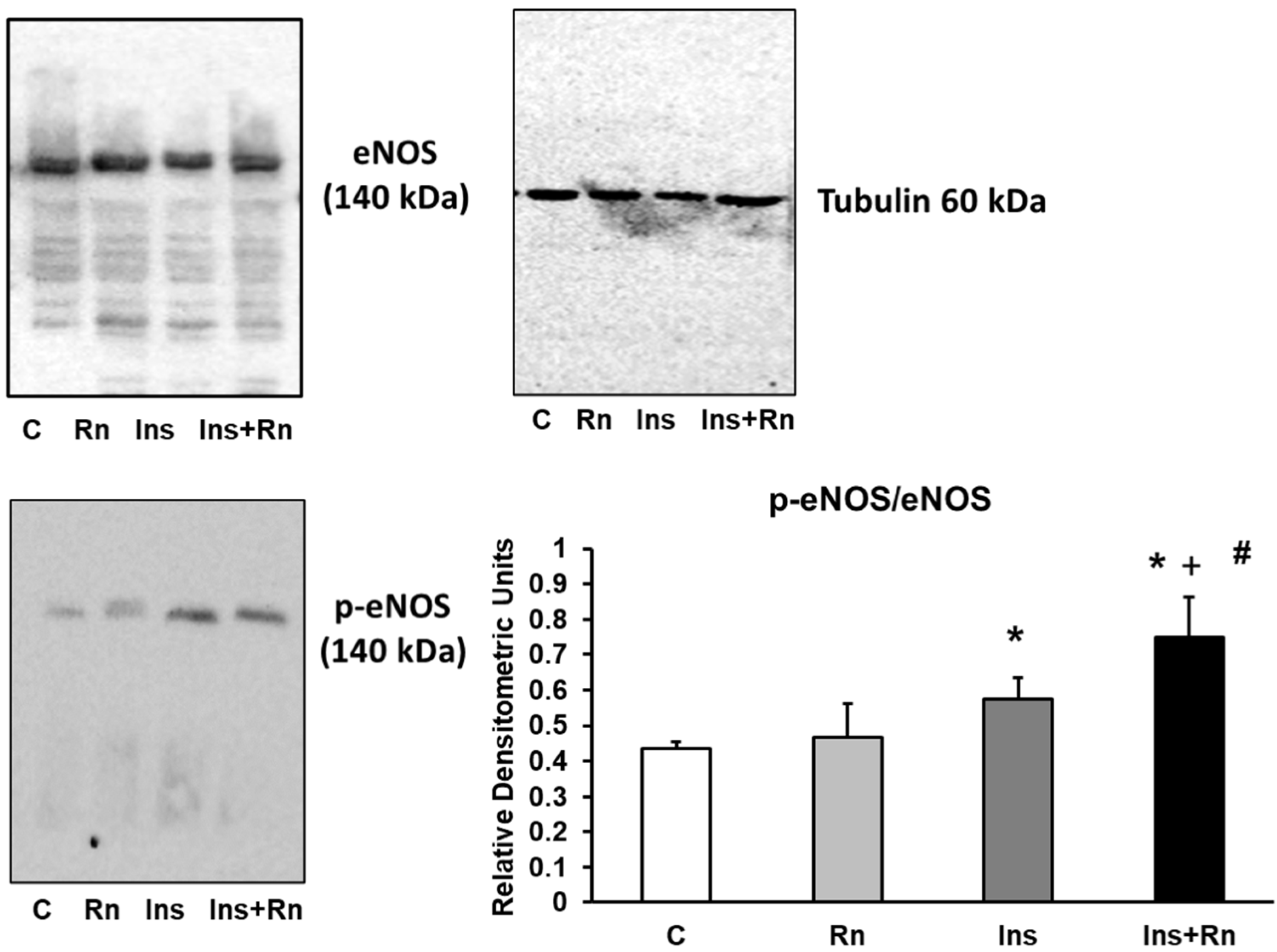
2.4. Expression of p-eNOS Protein
2.5. p-ERK Protein Expression
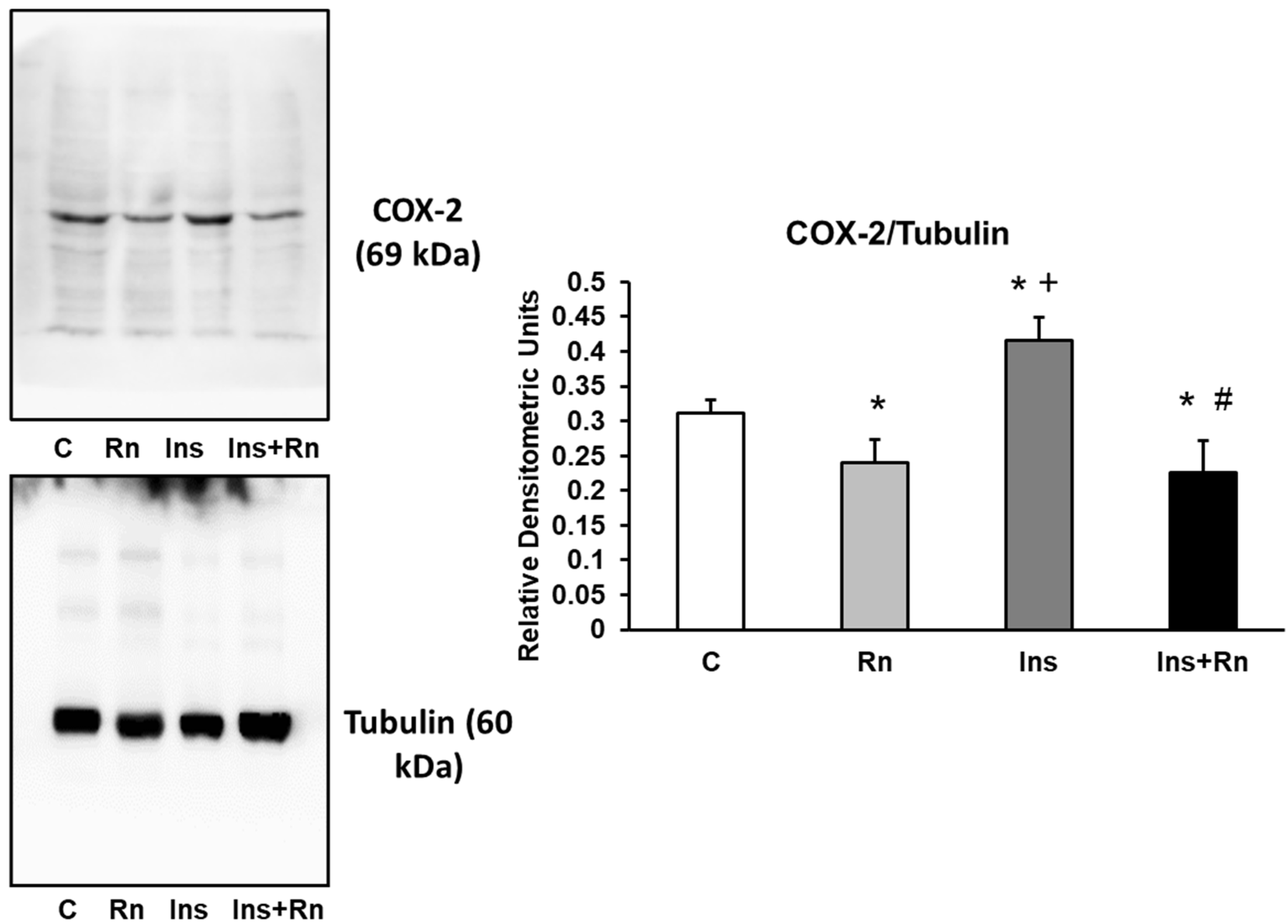
2.6. COX-2 Protein Expression
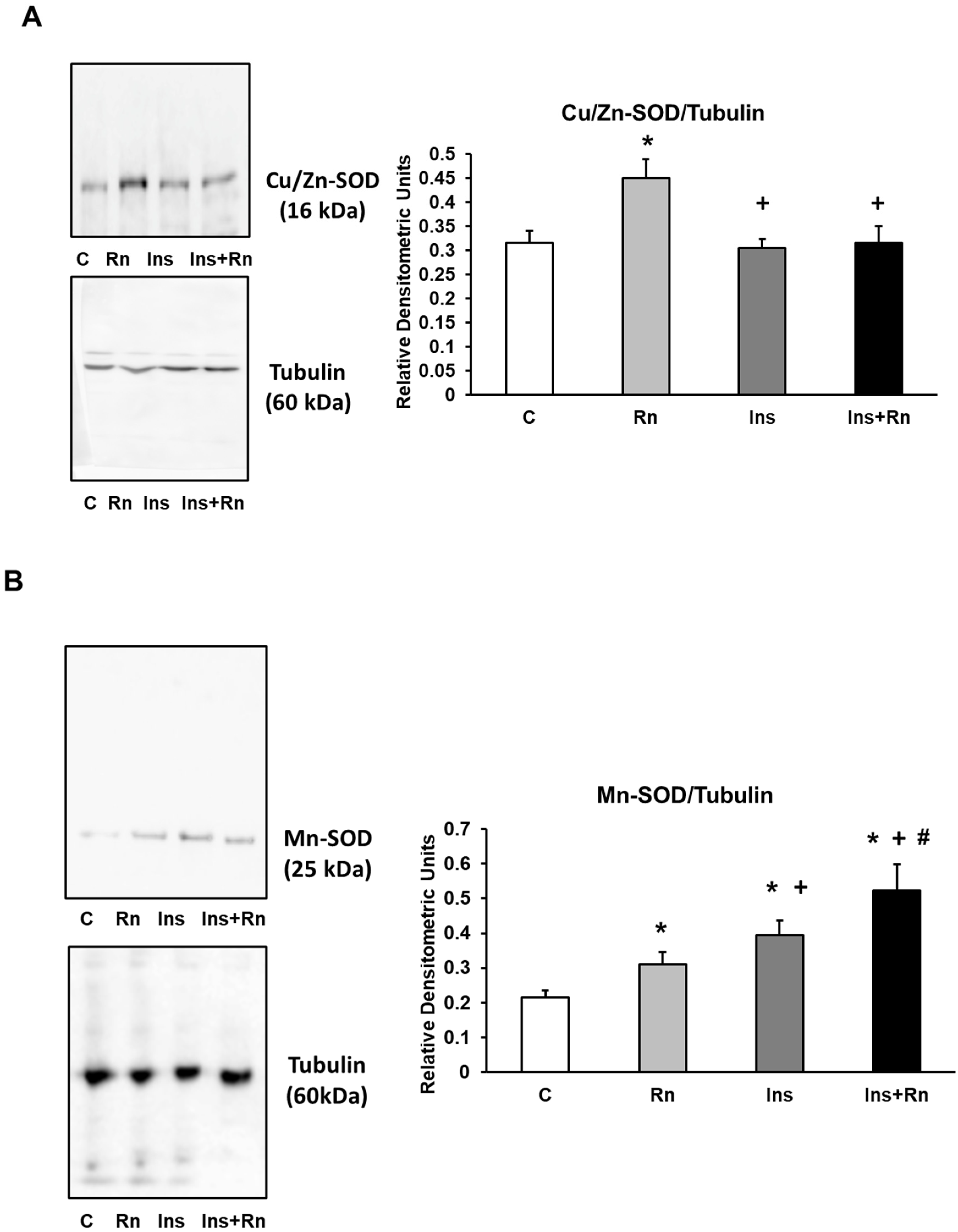
2.7. Expression of Cu/Zn-SOD and Mn-SOD Proteins
2.8. NF-κB and IκB Expression
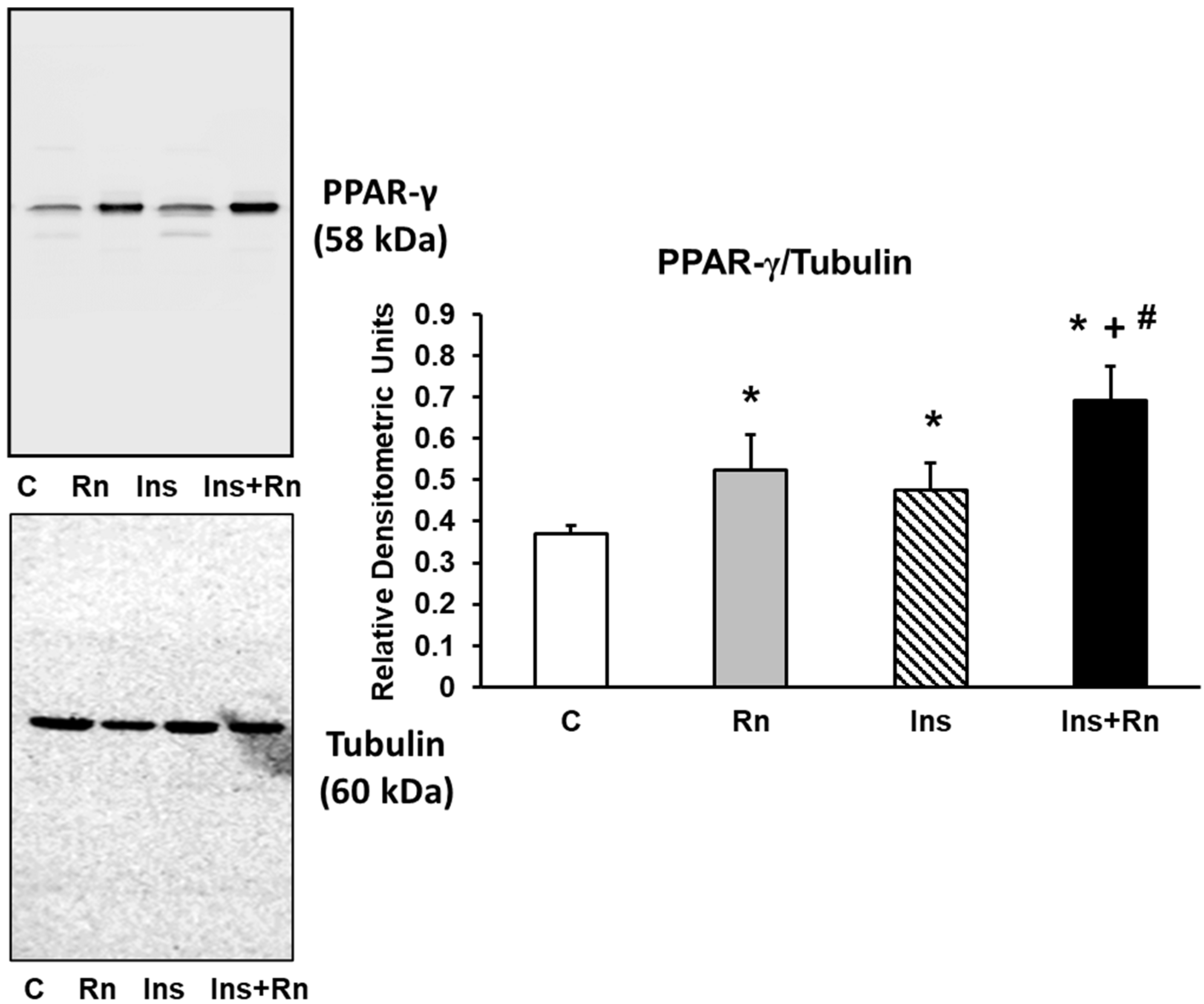
2.9. PPAR-γ Expression
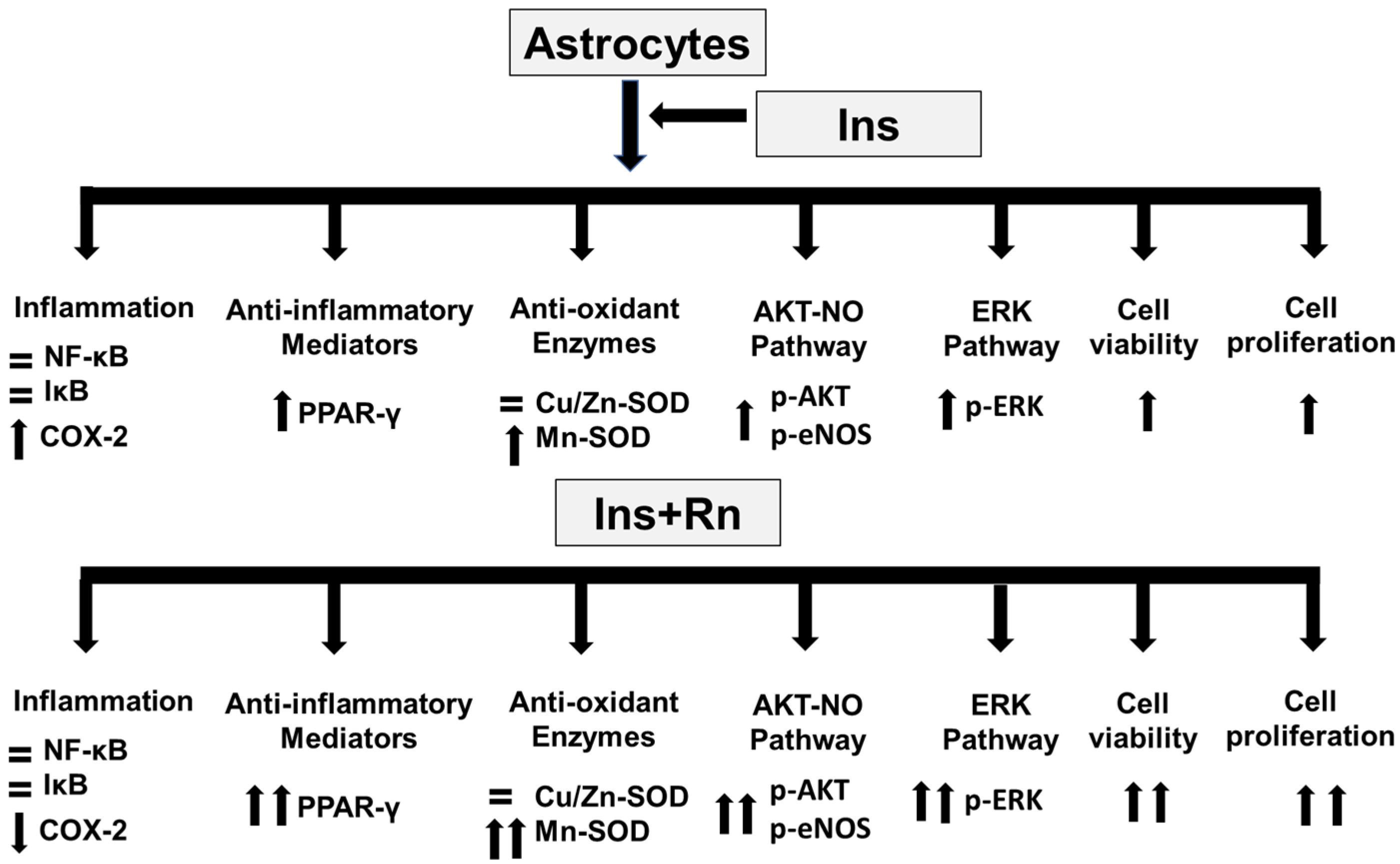
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Primary Culture of Cortical Astrocytes
4.3. MTT Assay
4.4. Trypan Blue Assay
4.5. Western Blot Analysis
4.6. Statistical Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rajasekar, N.; Dwivedi, S.; Nath, C.; Hanif, K.; Shukla, R. Protection of streptozotocin induced insulin receptor dysfunction, neuroinflammation and amyloidogenesis in astrocytes by insulin. Neuropharmacology 2014, 86, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Engelhardt, B. The anatomical and cellular basis of immune surveillance in the central nervous system. Nat. Rev. Immunol. 2012, 12, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldasoro, M.; Guerra-Ojeda, S.; Aguirre-Rueda, D.; Mauricio, M.D.; Vila, J.M.; Marchio, P.; Iradi, A.; Aldasoro, C.; Jorda, A.; Obrador, E.; et al. Effects of Ranolazine on Astrocytes and Neurons in Primary Culture. PLoS ONE 2016, 11, e0150619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiebaud, D.; Jacot, E.; DeFronzo, R.A.; Maeder, E.; Jequier, E.; Felber, J.P. The effect of graded doses of insulin on total glucose uptake, glucose oxidation, and glucose storage in man. Diabetes 1982, 31, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.M.; Barrett, E.J. Insulin transport into the brain. American journal of physiology. Cell Physiol. 2018, 315, C125–C136. [Google Scholar] [CrossRef] [PubMed]
- Shaughness, M.; Acs, D.; Brabazon, F.; Hockenbury, N.; Byrnes, K.R. Role of Insulin in Neurotrauma and Neurodegeneration: A Review. Front. Neurosci. 2020, 14, 547175. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.; Huang, J.; Chen, H.; Guo, X.; Yuan, Z. Insulin inhibits lipopolysaccharide-induced nitric oxide synthase expression in rat primary astrocytes. Brain Res. 2013, 1506, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Heni, M.; Hennige, A.M.; Peter, A.; Siegel-Axel, D.; Ordelheide, A.M.; Krebs, N.; Machicao, F.; Fritsche, A.; Häring, H.U.; Staiger, H. Insulin promotes glycogen storage and cell proliferation in primary human astrocytes. PLoS ONE 2011, 6, e21594. [Google Scholar] [CrossRef] [PubMed]
- Shahriyary, L.; Riazi, G.; Lornejad, M.R.; Ghezlou, M.; Bigdeli, B.; Delavari, B.; Mamashli, F.; Abbasi, S.; Davoodi, J.; Saboury, A.A. Effect of glycated insulin on the blood-brain barrier permeability: An in vitro study. Arch. Biochem. Biophys. 2018, 647, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Son, S.M.; Cha, M.Y.; Choi, H.; Kang, S.; Choi, H.; Lee, M.S.; Park, S.A.; Mook-Jung, I. Insulin-degrading enzyme secretion from astrocytes is mediated by an autophagy-based unconventional secretory pathway in Alzheimer disease. Autophagy 2016, 12, 784–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, L.; Guarda, M.; Meneses, M.J.; Macedo, M.P.; Vicente Miranda, H. Insulin-degrading enzyme: An ally against metabolic and neurodegenerative diseases. J. Pathol. 2021, 255, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.V.; Echouffo-Tcheugui, J.B.; Lam, C.S.P.; Inzucchi, S.E.; Tang, F.; McGuire, D.K.; Goyal, A.; Maddox, T.M.; Sperling, L.S.; Fonarow, G.C.; et al. Patterns of glucose-lowering medication use in patients with type 2 diabetes and heart failure. Insights from the Diabetes Collaborative Registry (DCR). Am. Heart J. 2018, 203, 25–29. [Google Scholar] [CrossRef]
- Heni, M.; Eckstein, S.S.; Schittenhelm, J.; Böhm, A.; Hogrefe, N.; Irmler, M.; Beckers, J.; Hrabě de Angelis, M.; Häring, H.U.; Fritsche, A.; et al. Ectopic fat accumulation in human astrocytes impairs insulin action. R. Soc. Open Sci. 2020, 7, 200701. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.W.; Boyd, F.T., Jr.; Kappy, M.S.; Raizada, M.K. Insulin binds to specific receptors and stimulates 2-deoxy-D-glucose uptake in cultured glial cells from rat brain. J. Biol. Chem. 1984, 259, 11672–11675. [Google Scholar] [CrossRef]
- Siddiqui, M.A.; Keam, S.J. Ranolazine: A review of its use in chronic stable angina pectoris. Drugs 2006, 66, 693–710. [Google Scholar] [CrossRef]
- Storey, K.M.; Wang, J.; Garberich, R.F.; Bennett, N.M.; Traverse, J.H.; Arndt, T.L.; Schmidt, C.W.; Henry, T.D. Long-Term (3 Years) Outcomes of Ranolazine Therapy for Refractory Angina Pectoris (from the Ranolazine Refractory Registry). Am. J. Cardiol. 2020, 129, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Stone, P.H.; Chaitman, B.R.; Stocke, K.; Sano, J.; DeVault, A.; Koch, G.G. The anti-ischemic mechanism of action of ranolazine in stable ischemic heart disease. J. Am. Coll. Cardiol. 2010, 56, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Marchio, P.; Guerra-Ojeda, S.; Aldasoro, M.; Valles, S.L.; Martín-Gonzalez, I.; Martínez-León, J.B.; Mauricio, M.D.; Vila, J.M. Relaxant and antiadrenergic effects of ranolazine in human saphenous vein. Eur. J. Cardio-Thorac. Surg. 2020, 58, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Nusca, A.; Bernardini, F.; Mangiacapra, F.; Maddaloni, E.; Melfi, R.; Ricottini, E.; Piccirillo, F.; Manfrini, S.; Ussia, G.P.; Grigioni, F. Ranolazine Improves Glycemic Variability and Endothelial Function in Patients with Diabetes and Chronic Coronary Syndromes: Results from an Experimental Study. J. Diabetes Res. 2021, 2021, 4952447. [Google Scholar] [CrossRef] [PubMed]
- Shryock, J.C.; Belardinelli, L. Inhibition of late sodium current to reduce electrical and mechanical dysfunction of ischaemic myocardium. Br. J. Pharmacol. 2008, 153, 1128–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaitman, B.R. Ranolazine for the treatment of chronic angina and potential use in other cardiovascular conditions. Circulation 2006, 113, 2462–2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belardinelli, R.; Lacalaprice, F.; Faccenda, E.; Volpe, L. Clinical benefits of a metabolic approach in the cardiac rehabilitation of patients with coronary artery disease. Am. J. Cardiol. 2006, 98, 25J–33J. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.; Amin, G.; Abidi, E.; Altara, R.; Booz, G.W.; Zouein, F.A. Role of ranolazine in heart failure: From cellular to clinic perspective. Eur. J. Pharmacol. 2022, 919, 174787. [Google Scholar] [CrossRef]
- Aldakkak, M.; Camara, A.K.; Heisner, J.S.; Yang, M.; Stowe, D.F. Ranolazine reduces Ca2+ overload and oxidative stress and improves mitochondrial integrity to protect against ischemia reperfusion injury in isolated hearts. Pharmacol. Res. 2011, 64, 381–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambarat, C.A.; Elgendy, I.Y.; Handberg, E.M.; Bairey Merz, C.N.; Wei, J.; Minissian, M.B.; Nelson, M.D.; Thomson, L.; Berman, D.S.; Shaw, L.J.; et al. Late sodium channel blockade improves angina and myocardial perfusion in patients with severe coronary microvascular dysfunction: Women’s Ischemia Syndrome Evaluation-Coronary Vascular Dysfunction ancillary study. Int. J. Cardiol. 2019, 276, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.C.; Lee, H.L.; Chang, G.J.; Wo, H.T.; Yen, T.H.; Wen, M.S.; Chu, Y.; Liu, H.T.; Chang, P.C. Mechanisms of ranolazine pretreatment in preventing ventricular tachyarrhythmias in diabetic db/db mice with acute regional ischemia-reperfusion injury. Sci. Rep. 2020, 10, 20032. [Google Scholar] [CrossRef]
- Deshmukh, S.H.; Patel, S.R.; Pinassi, E.; Mindrescu, C.; Hermance, E.V.; Infantino, M.N.; Coppola, J.T.; Staniloae, C.S. Ranolazine improves endothelial function in patients with stable coronary artery disease. Coron. Artery Dis. 2009, 20, 343–347. [Google Scholar] [CrossRef]
- Morrow, D.A.; Scirica, B.M.; Chaitman, B.R.; McGuire, D.K.; Murphy, S.A.; Karwatowska-Prokopczuk, E.; McCabe, C.H.; Braunwald, E.; MERLIN-TIMI 36 Investigators. Evaluation of the glycometabolic effects of ranolazine in patients with and without diabetes mellitus in the MERLIN-TIMI 36 randomized controlled trial. Circulation 2019, 119, 2032–2039. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.V.; McGuire, D.K.; Spertus, J.A.; Li, Y.; Yue, P.; Ben-Yehuda, O.; Belardinelli, L.; Jones, P.G.; Olmsted, A.; Chaitman, B.R.; et al. Effectiveness of ranolazine in patients with type 2 diabetes mellitus and chronic stable angina according to baseline hemoglobin A1c. Am. Heart J. 2014, 168, 457–465.e2. [Google Scholar] [CrossRef] [PubMed]
- Koltun, D.O.; Parkhill, E.Q.; Elzein, E.; Kobayashi, T.; Jiang, R.H.; Li, X.; Perry, T.D.; Avila, B.; Wang, W.Q.; Hirakawa, R.; et al. Discovery of triazolopyridinone GS-462808, a late sodium current inhibitor (Late INai) of the cardiac Nav1.5 channel with improved efficacy and potency relative to ranolazine. Bioorg. Med. Chem. Lett. 2016, 26, 3207–3211. [Google Scholar] [CrossRef] [PubMed]
- Chunchai, T.; Arinno, A.; Ongnok, B.; Pantiya, P.; Khuanjing, T.; Prathumsap, N.; Maneechote, C.; Chattipakorn, N.; Chattipakorn, S.C. Ranolazine alleviated cardiac/brain dysfunction in doxorubicin-treated rats. Exp. Mol. Pathol. 2022, 127, 104818. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.V.; Kosiborod, M.; McGuire, D.K.; Li, Y.; Yue, P.; Ben-Yehuda, O.; Spertus, J.A. Effects of ranolazine on quality of life among patients with diabetes mellitus and stable angina. JAMA Intern. Med. 2014, 174, 1403–1405. [Google Scholar] [CrossRef]
- Ning, Y.; Zhen, W.; Fu, Z.; Jiang, J.; Liu, D.; Belardinelli, L.; Dhalla, A.K. Ranolazine increases β-cell survival and improves glucose homeostasis in low-dose streptozotocin-induced diabetes in mice. J. Pharmacol. Exp. Ther. 2011, 337, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Peters, C.H.; Sokolov, S.; Rajamani, S.; Ruben, P.C. Effects of the antianginal drug, ranolazine, on the brain sodium channel Na(V)1.2 and its modulation by extracellular protons. Br. J. Pharmacol. 2013, 169, 704–716. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.Y.; Johnston, D.; Gray, R. Slowly inactivating component of Na+ current in peri-somatic region of hippocampal CA1 pyramidal neurons. J. Neurophysiol. 2013, 109, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Virsolvy, A.; Farah, C.; Pertuit, N.; Kong, L.; Lacampagne, A.; Reboul, C.; Aimond, F.; Richard, S. Antagonism of Nav channels and α1-adrenergic receptors contributes to vascular smooth muscle effects of ranolazine. Sci. Rep. 2015, 5, 17969. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.S.; Lo, Y.C.; Peng, H.; Hsu, T.I.; Wu, S.N. Effects of ranolazine, a novel anti-anginal drug, on ion currents and membrane potential in pituitary tumor GH (3) cells and NG108-15 neuronal cells. J. Pharmacol. Sci. 2009, 110, 295–305. [Google Scholar] [CrossRef]
- Liao, Y.; Hung, M.C. Physiological regulation of Akt activity and stability. Am. J. Transl. Res. 2010, 2, 19–42. [Google Scholar] [PubMed]
- Gerrits, A.J.; Koekman, C.A.; van Haeften, T.W.; Akkerman, J.W. Platelet tissue factor synthesis in type 2 diabetic patients is resistant to inhibition by insulin. Diabetes 2010, 59, 1487–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, W.; Qi, Y. An updated meta-analysis of endothelial nitric oxide synthase gene: Three well-characterized polymorphisms with hypertension. PLoS ONE 2011, 6, e24266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoukry, A.; Shalaby, S.M.; Abdelazim, S.; Abdelazim, M.; Ramadan, A.; Ismail, M.I.; Fouad, M. Endothelial nitric oxide synthase gene polymorphisms and the risk of diabetic nephropathy in type 2 diabetes mellitus. Genet. Test. Mol. Biomark. 2012, 16, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Eröz, R.; Bahadir, A.; Dikici, S.; Tasdemir, S. Association of endothelial nitric oxide synthase gene polymorphisms (894G/T, -786T/C, G10T) and clinical findings in patients with migraine. Neuromolecular Med. 2014, 16, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kahn, C.R. Insulin regulation of mitogen-activated protein kinase kinase (MEK), mitogen-activated protein kinase and casein kinase in the cell nucleus: A possible role in the regulation of gene expression. Biochem. J. 1997, 323 Pt 3, 621–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabatabaie, T.; Waldon, A.M.; Jacob, J.M.; Floyd, R.A.; Kotake, Y. COX-2 inhibition prevents insulin-dependent diabetes in low-dose streptozotocin-treated mice. Biochem. Biophys. Res. Commun. 2000, 273, 699–704. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide dismutases: You’ve come a long way, baby. Antioxid. Redox Signal. 2014, 20, 1548–1549. [Google Scholar] [CrossRef] [PubMed]
- Muscogiuri, G.; Salmon, A.B.; Aguayo-Mazzucato, C.; Li, M.; Balas, B.; Guardado-Mendoza, R.; Giaccari, A.; Reddick, R.L.; Reyna, S.M.; Weir, G.; et al. Genetic disruption of SOD1 gene causes glucose intolerance and impairs β-cell function. Diabetes 2013, 62, 4201–4207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karve, I.P.; Taylor, J.M.; Crack, P.J. The contribution of astrocytes and microglia to traumatic brain injury. Br. J. Pharmacol. 2016, 173, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D. The brain as a target for inflammatory processes and neuroprotective strategies. Ann. N. Y. Acad. Sci. 2007, 1122, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Rueda, D.; Guerra-Ojeda, S.; Aldasoro, M.; Iradi, A.; Obrador, E.; Ortega, A.; Mauricio, M.D.; Vila, J.M.; Valles, S.L. Astrocytes protect neurons from Aβ1-42 peptide-induced neurotoxicity increasing TFAM and PGC-1 and decreasing PPAR-γ and SIRT-1. Int. J. Med. Sci. 2015, 12, 48–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouyakdan, K.; Martin, H.; Liénard, F.; Budry, L.; Taib, B.; Rodaros, D.; Chrétien, C.; Biron, É.; Husson, Z.; Cota, D.; et al. The gliotransmitter ACBP controls feeding and energy homeostasis via the melanocortin system. J. Clin. Investig. 2019, 129, 2417–2430. [Google Scholar] [CrossRef]
- MacDonald, A.J.; Holmes, F.E.; Beall, C.; Pickering, A.E.; Ellacott, K. Regulation of food intake by astrocytes in the brainstem dorsal vagal complex. Glia 2020, 68, 1241–1254. [Google Scholar] [CrossRef] [Green Version]
- González-García, I.; Gruber, T.; García-Cáceres, C. Insulin action on astrocytes: From energy homeostasis to behaviour. J. Neuroendocrinol. 2021, 33, e12953. [Google Scholar] [CrossRef] [PubMed]
- Spielman, L.J.; Bahniwal, M.; Little, J.P.; Walker, D.G.; Klegeris, A. Insulin Modulates In Vitro Secretion of Cytokines and Cytotoxins by Human Glial Cells. Curr. Alzheimer Res. 2015, 12, 684–693. [Google Scholar] [CrossRef]
- Haas, C.B.; de Carvalho, A.K.; Muller, A.P.; Eggen, B.; Portela, L.V. Insulin activates microglia and increases COX-2/IL-1β expression in young but not in aged hippocampus. Brain Res. 2020, 1741, 146884. [Google Scholar] [CrossRef] [PubMed]
- Dimmeler, S.; Fleming, I.; Fisslthaler, B.; Hermann, C.; Busse, R.; Zeiher, A.M. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999, 399, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, M.; Naghdi, N.; Choopani, S. Intra CA1 insulin microinjection improves memory consolidation and retrieval. Peptides 2007, 28, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Choopani, S.; Moosavi, M.; Naghdi, N. Involvement of nitric oxide in insulin induced memory improvement. Peptides 2008, 29, 898–903. [Google Scholar] [CrossRef]
- McCrimmon, R.J.; Ryan, C.M.; Frier, B.M. Diabetes and cognitive dysfunction. Lancet 2012, 379, 2291–2299. [Google Scholar] [CrossRef]
- Nefs, G.; Hendrieckx, C.; Reddy, P.; Browne, J.L.; Bot, M.; Dixon, J.; Kyrios, M.; Speight, J.; Pouwer, F. Comorbid elevated symptoms of anxiety and depression in adults with type 1 or type 2 diabetes: Results from the International Diabetes MILES Study. J. Diabetes Its Complicat. 2019, 33, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Arvanitakis, Z.; Wilson, R.S.; Bienias, J.L.; Evans, D.A.; Bennett, D.A. Diabetes mellitus and risk of Alzheimer disease and decline in cognitive function. Arch. Neurol. 2004, 61, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, A.; Sah, S.P. Insulin signaling pathway and related molecules: Role in neurodegeneration and Alzheimer’s disease. Neurochem. Int. 2020, 135, 104707. [Google Scholar] [CrossRef]
- Cheong, J.; de Pablo-Fernandez, E.; Foltynie, T.; Noyce, A.J. The Association Between Type 2 Diabetes Mellitus and Parkinson’s Disease. J. Parkinson’s Dis. 2020, 10, 775–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabbouj, S.; Ryhänen, S.; Marttinen, M.; Wittrahm, R.; Takalo, M.; Kemppainen, S.; Martiskainen, H.; Tanila, H.; Haapasalo, A.; Hiltunen, M.; et al. Altered Insulin Signaling in Alzheimer’s Disease Brain - Special Emphasis on PI3K-Akt Pathway. Front. Neurosci. 2019, 13, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbot, K.; Wang, H.Y.; Kazi, H.; Han, L.Y.; Bakshi, K.P.; Stucky, A.; Fuino, R.L.; Kawaguchi, K.R.; Samoyedny, A.J.; Wilson, R.S.; et al. Demonstrated brain insulin resistance in Alzheimer’s disease patients is associated with IGF-1 resistance, IRS-1 dysregulation, and cognitive decline. J. Clin. Investig. 2012, 122, 1316–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarchoan, M.; Toledo, J.B.; Lee, E.B.; Arvanitakis, Z.; Kazi, H.; Han, L.Y.; Louneva, N.; Lee, V.M.; Kim, S.F.; Trojanowski, J.Q.; et al. Abnormal serine phosphorylation of insulin receptor substrate 1 is associated with tau pathology in Alzheimer’s disease and tauopathies. Acta Neuropathol. 2014, 128, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metz, H.E.; Houghton, A.M. Insulin receptor substrate regulation of phosphoinositide 3-kinase. Clin. Cancer Res. 2011, 17, 206–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassano, V.; Leo, A.; Tallarico, M.; Nesci, V.; Cimellaro, A.; Fiorentino, T.V.; Citraro, R.; Hribal, M.L.; De Sarro, G.; Perticone, F.; et al. Metabolic and Cognitive Effects of Ranolazine in Type 2 Diabetes Mellitus: Data from an in vivo Model. Nutrients 2020, 12, 382. [Google Scholar] [CrossRef] [PubMed]
- Marasciulo, F.L.; Montagnani, M.; Potenza, M.A. Endothelin-1: The yin and yang on vascular function. Curr. Med. Chem. 2006, 13, 1655–1665. [Google Scholar] [CrossRef]
- Lee, J.H.; Jahrling, J.B.; Denner, L.; Dineley, K.T. Targeting Insulin for Alzheimer’s Disease: Mechanisms, Status and Potential Directions. J. Alzheimer’s Dis. 2018, 64, S427–S453. [Google Scholar] [CrossRef] [PubMed]
- Brabazon, F.; Bermudez, S.; Shaughness, M.; Khayrullina, G.; Byrnes, K.R. The effects of insulin on the inflammatory activity of BV2 microglia. PLoS ONE 2018, 13, e0201878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, F.; Auwerx, J. PPAR(gamma) and glucose homeostasis. Annu. Rev. Nutr. 2002, 22, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Norris, A.W.; Chen, L.; Fisher, S.J.; Szanto, I.; Ristow, M.; Jozsi, A.C.; Hirshman, M.F.; Rosen, E.D.; Goodyear, L.J.; Gonzalez, F.J.; et al. Muscle-specific PPARgamma-deficient mice develop increased adiposity and insulin resistance but respond to thiazolidinediones. J. Clin. Investig. 2003, 112, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Montaigne, D.; Butruille, L.; Staels, B. PPAR control of metabolism and cardiovascular functions. Nature reviews. Cardiology 2021, 18, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Gao, M.; Yang, P.; Liu, D.; Wang, D.; Song, F.; Zhang, X.; Liu, Y. Insulin promotes macrophage phenotype transition through PI3K/Akt and PPAR-γ signaling during diabetic wound healing. J. Cell. Physiol. 2019, 234, 4217–4231. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Schiffrin, E.L. Peroxisome proliferator-activated receptors in vascular biology-molecular mechanisms and clinical implications. Vasc. Pharmacol. 2006, 45, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Cho, J.H.; Lee, S.; Lee, W.; Chang, S.C.; Chung, H.Y.; Moon, H.R.; Lee, J. Neuroprotective effects of MHY908, a PPAR α/γ dual agonist, in a MPTP-induced Parkinson’s disease model. Brain Res. 2019, 1704, 47–58. [Google Scholar] [CrossRef]
- Rajasekar, N.; Nath, C.; Hanif, K.; Shukla, R. Intranasal Insulin Administration Ameliorates Streptozotocin (ICV)-Induced Insulin Receptor Dysfunction, Neuroinflammation, Amyloidogenesis, and Memory Impairment in Rats. Mol. Neurobiol. 2017, 54, 6507–6522. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Eyster, C.; Newhardt, M.F.; Giorgione, J.R.; Kinter, C.; Young, Z.T.; Kinter, M.; Humphries, K.M. Insulin signaling alters antioxidant capacity in the diabetic heart. Redox Biol. 2021, 47, 102140. [Google Scholar] [CrossRef] [PubMed]
- Ramalingayya, G.V.; Sonawane, V.; Cheruku, S.P.; Kishore, A.; Nayak, P.G.; Kumar, N.; Shenoy, R.S.; Nandakumar, K. Insulin Protects against Brain Oxidative Stress with an Apparent Effect on Episodic Memory in Doxorubicin-Induced Cognitive Dysfunction in Wistar Rats. J. Environ. Pathol. Toxicol. Oncol. 2017, 36, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, K.L.; Salmon, A.B.; Hohnen-Behrens, C.; Turner, N.; Hoy, A.J.; Maghzal, G.J.; Stocker, R.; Van Remmen, H.; Kraegen, E.W.; Cooney, G.J.; et al. Insulin resistance is a cellular antioxidant defense mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 17787–17792. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Ding, W.; Bei, Y.; Xiao, Y.; Tong, H.D.; Wang, L.B.; Ai, L.Y. Insulin is a potential antioxidant for diabetes-associated cognitive decline via regulating Nrf2 dependent antioxidant enzymes. Biomed. Pharmacother. 2018, 104, 474–484. [Google Scholar] [CrossRef]
- McCormack, J.G.; Barr, R.L.; Wolff, A.A.; Lopaschuk, G.D. Ranolazine stimulates glucose oxidation in normoxic, ischemic, and reperfused ischemic rat hearts. Circulation 1996, 93, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Zhao, L.; Chai, W.; Dong, Z.; Cao, W.; Liu, Z. Ranolazine recruits muscle microvasculature and enhances insulin action in rats. J. Physiol. 2013, 591, 5235–5249. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhang, Y.; Lin, J.; Zheng, H.; Peng, J.; Huang, W. Efficacy and Safety of Ranolazine in Diabetic Patients: A Systematic Review and Meta-analysis. Ann. Pharmacother. 2017, 52, 415–424. [Google Scholar] [CrossRef]
- Gilbert, B.W.; Sherard, M.; Little, L.; Branstetter, J.; Meister, A.; Huffman, J. Antihyperglycemic and Metabolic Effects of Ranolazine in Patients with Diabetes Mellitus. Am. J. Cardiol. 2018, 121, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.; Goncalves, E. Diabetogenic effects of cardioprotective drugs. Diabetes Obes. Metab. 2021, 23, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Teoh, I.H.; Banerjee, M. Effect of ranolazine on glycaemia in adults with and without diabetes: A meta-analysis of randomised controlled trials. Open Heart 2018, 5, e000706. [Google Scholar] [CrossRef] [PubMed]
- Terruzzi, I.; Montesano, A.; Senesi, P.; Vacante, F.; Benedini, S.; Luzi, L. Ranolazine promotes muscle differentiation and reduces oxidative stress in C2C12 skeletal muscle cells. Endocrine 2017, 58, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Caminiti, G.; Fossati, C.; Battaglia, D.; Massaro, R.; Rosano, G.; Volterrani, M. Ranolazine improves insulin resistance in non-diabetic patients with coronary heart disease. A pilot study. Int. J. Cardiol. 2016, 219, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Theile, J.W.; Cummins, T.R. Recent developments regarding voltage-gated sodium channel blockers for the treatment of inherited and acquired neuropathic pain syndromes. Front. Pharmacol. 2011, 2, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahlig, K.M.; Lepist, I.; Leung, K.; Rajamani, S.; George, A.L. Ranolazine selectively blocks persistent current evoked by epilepsy-associated Naν1.1 mutations. Br. J. Pharmacol. 2010, 161, 1414–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nodera, H.; Rutkove, S.B. Changes of the peripheral nerve excitability in vivo induced by the persistent Na+ current blocker ranolazine. Neurosci. Lett. 2012, 518, 36–40. [Google Scholar] [CrossRef]
- Elkholy, S.E.; Elaidy, S.M.; El-Sherbeeny, N.A.; Toraih, E.A.; El-Gawly, H.W. Neuroprotective effects of ranolazine versus pioglitazone in experimental diabetic neuropathy: Targeting Nav1.7 channels and PPAR-γ. Life Sci. 2020, 250, 117557. [Google Scholar] [CrossRef] [PubMed]
- Rouhana, S.; Virsolvy, A.; Fares, N.; Richard, S.; Thireau, J. Ranolazine: An Old Drug with Emerging Potential; Lessons from Pre-Clinical and Clinical Investigations for Possible Repositioning. Pharmaceuticals 2021, 15, 31. [Google Scholar] [CrossRef]
- Efentakis, P.; Andreadou, I.; Bibli, S.I.; Vasileiou, S.; Dagres, N.; Zoga, A.; Lougiakis, N.; Kremastinos, D.T.; Iliodromitis, E.K. Ranolazine triggers pharmacological preconditioning and postconditioning in anesthetized rabbits through activation of RISK pathway. Eur. J. Pharmacol. 2016, 789, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Stockert, J.C.; Blázquez-Castro, A.; Cañete, M.; Horobin, R.W.; Villanueva, A. MTT assay for cell viability: Intracellular localization of the formazan product is in lipid droplets. Acta Histochemical. 2012, 114, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. Mitochondrial oxygen radical generation and leak: Sites of production in states 4 and 3, organ specificity, and relation to aging and longevity. J. Bioenerg. Biomembr. 1999, 31, 347–366. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seeding Cells (×104/35 mm Dish) | 5 Days of Culture | 24 h Treatment | % Proliferation | |
|---|---|---|---|---|
| Control | 7 | 12.86 ± 0.32 | 12.97 ± 0.24 | 0.85 |
| Rn | 7 | 12.87 ± 0.25 | 16.77 ± 0.35 | 30.31 * |
| Ins | 7 | 12.85 ± 0.23 | 16.60 ± 0.37 | 29.18 * |
| Ins+Rn | 7 | 12.88 ± 0.26 | 17.25 ± 0.35 | 33.91 *,# |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jordá, A.; Aldasoro, M.; Campo-Palacio, I.; Vila, J.M.; Aldasoro, C.; Campos-Campos, J.; Colmena, C.; Singh, S.K.; Obrador, E.; Valles, S.L. Facilitation of Insulin Effects by Ranolazine in Astrocytes in Primary Culture. Int. J. Mol. Sci. 2022, 23, 11969. https://doi.org/10.3390/ijms231911969
Jordá A, Aldasoro M, Campo-Palacio I, Vila JM, Aldasoro C, Campos-Campos J, Colmena C, Singh SK, Obrador E, Valles SL. Facilitation of Insulin Effects by Ranolazine in Astrocytes in Primary Culture. International Journal of Molecular Sciences. 2022; 23(19):11969. https://doi.org/10.3390/ijms231911969
Chicago/Turabian StyleJordá, Adrián, Martin Aldasoro, Ignacio Campo-Palacio, Jose M. Vila, Constanza Aldasoro, Juan Campos-Campos, Carlos Colmena, Sandeep Kumar Singh, Elena Obrador, and Soraya L. Valles. 2022. "Facilitation of Insulin Effects by Ranolazine in Astrocytes in Primary Culture" International Journal of Molecular Sciences 23, no. 19: 11969. https://doi.org/10.3390/ijms231911969