
Calcium Imaging Reveals Fast Tuning Dynamics of Hippocampal Place Cells and CA1 Population Activity during Free Exploration Task in Mice

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
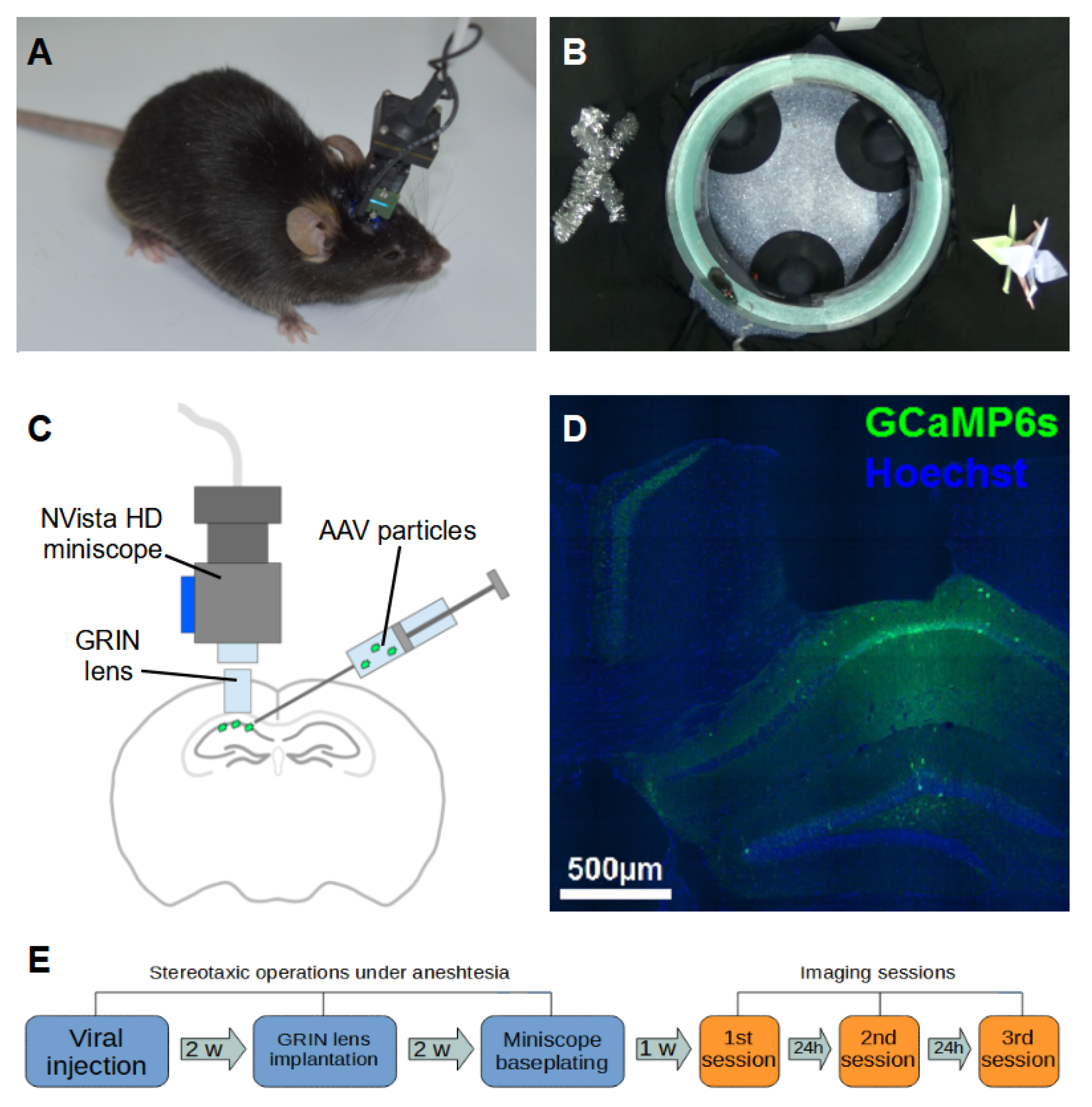
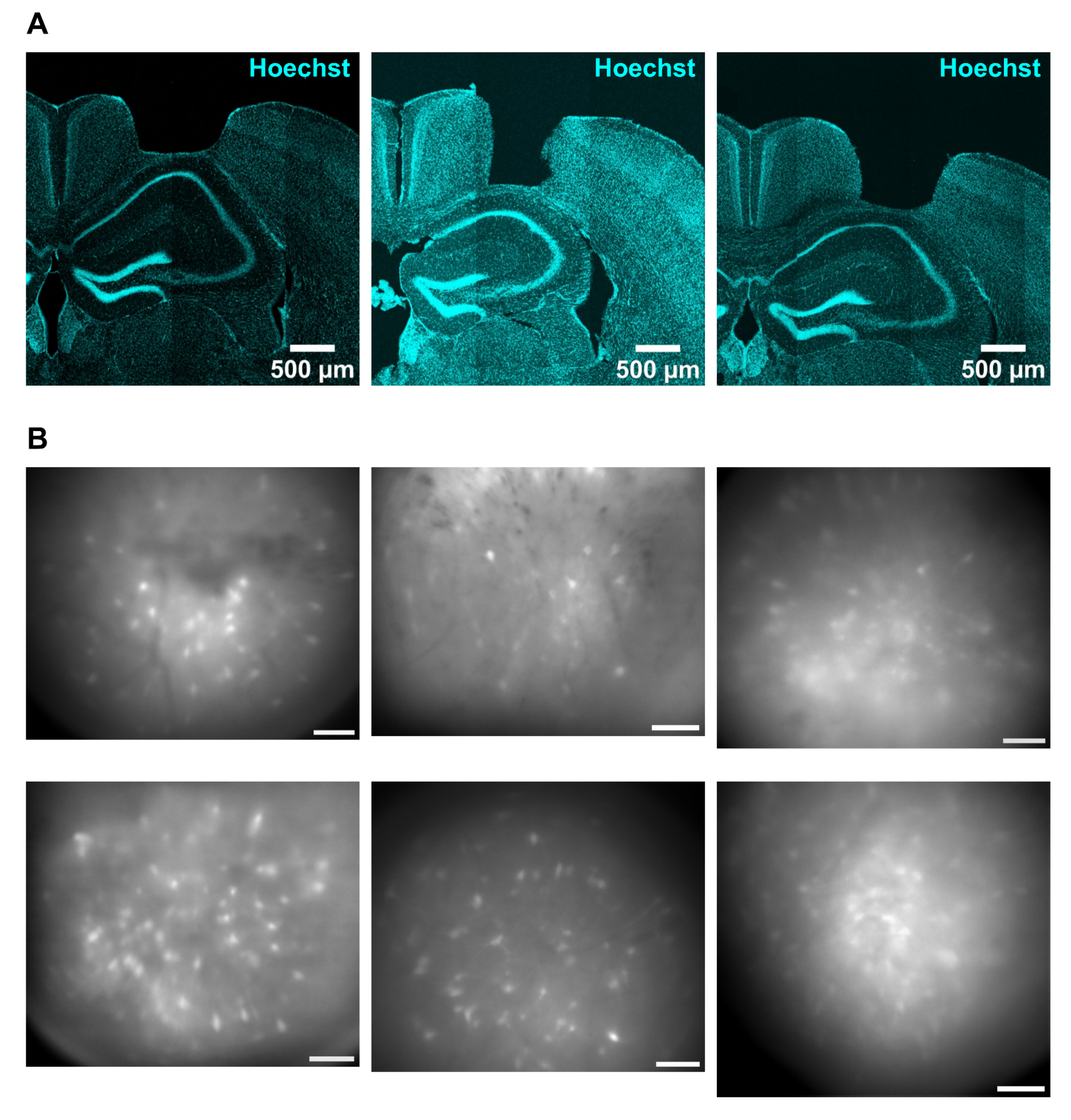
2.1. Animals and Surgical Procedures
2.2. Miniscope Imaging in Freely Behaving Mice
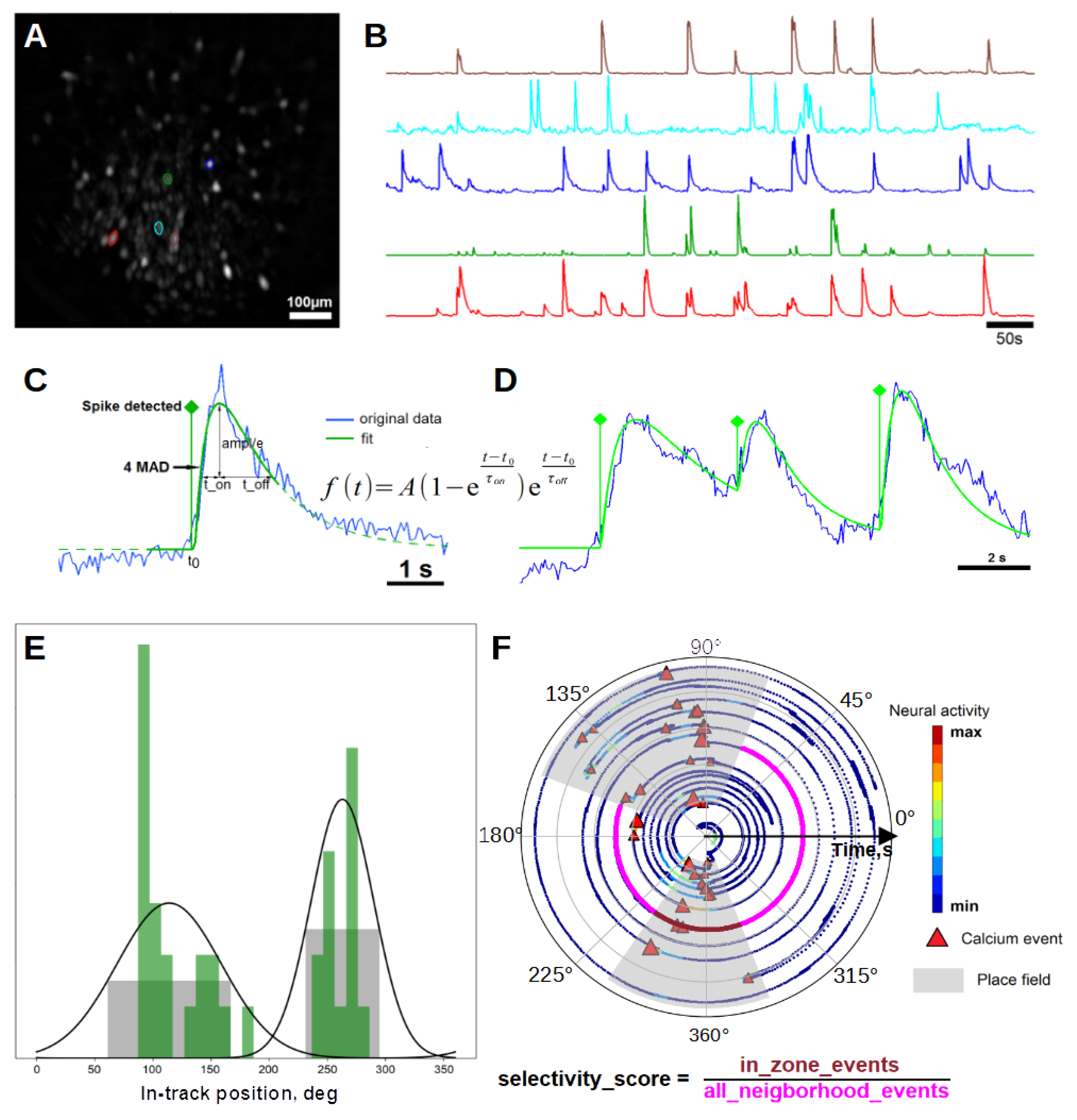
2.3. Neural and Behavioral Data Processing
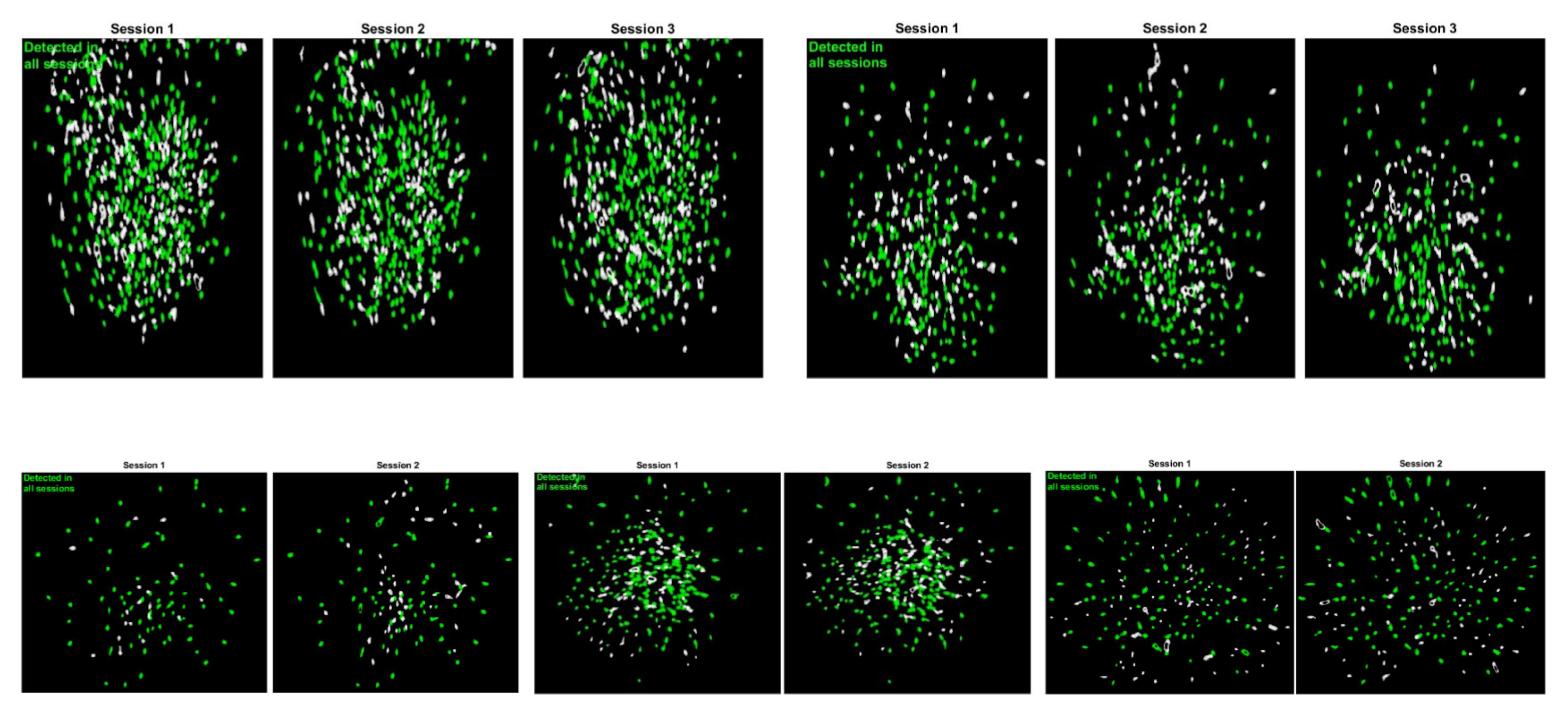
2.4. Place Cell Detection
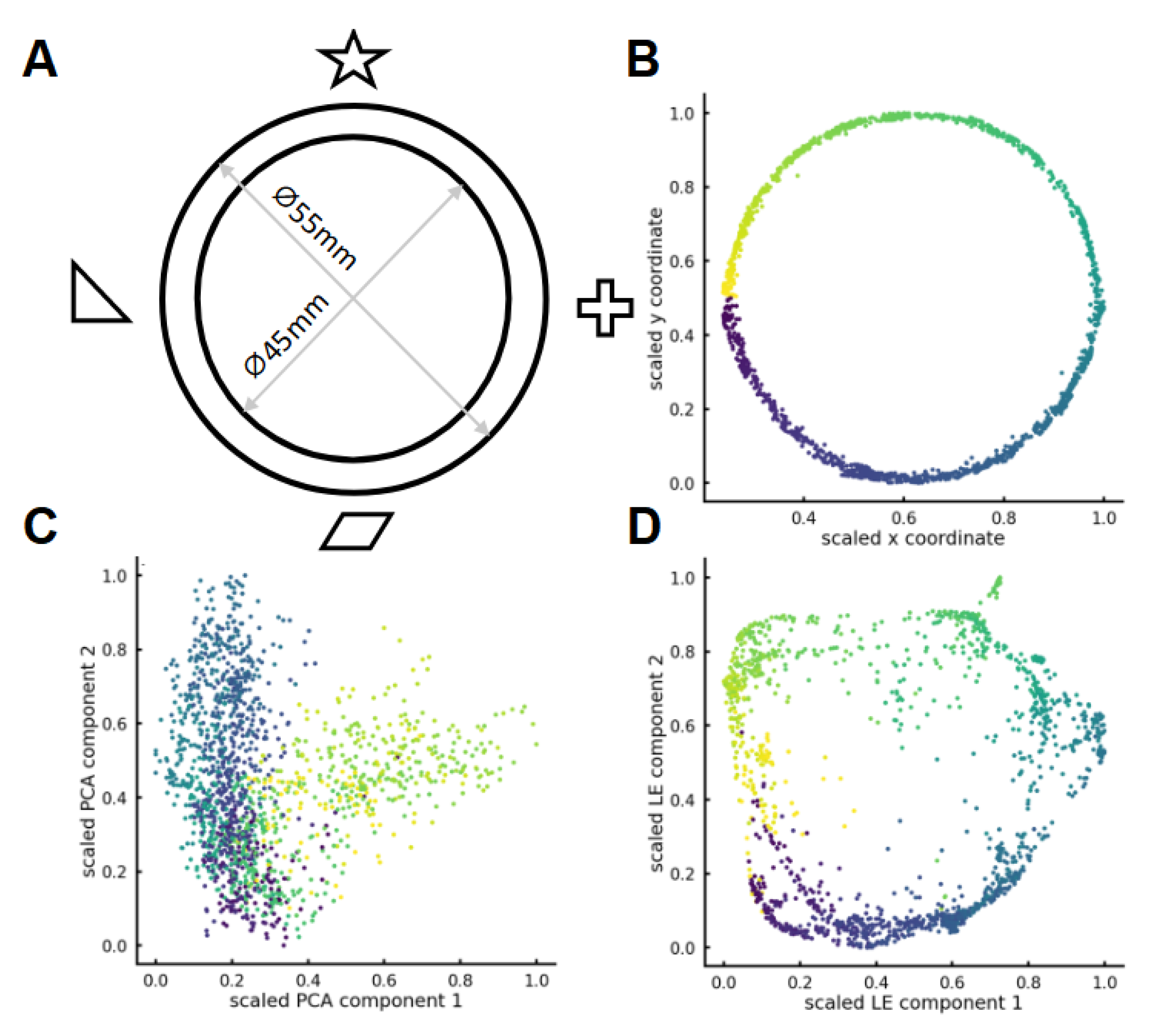
2.5. Dimensionality Reduction
3. Results
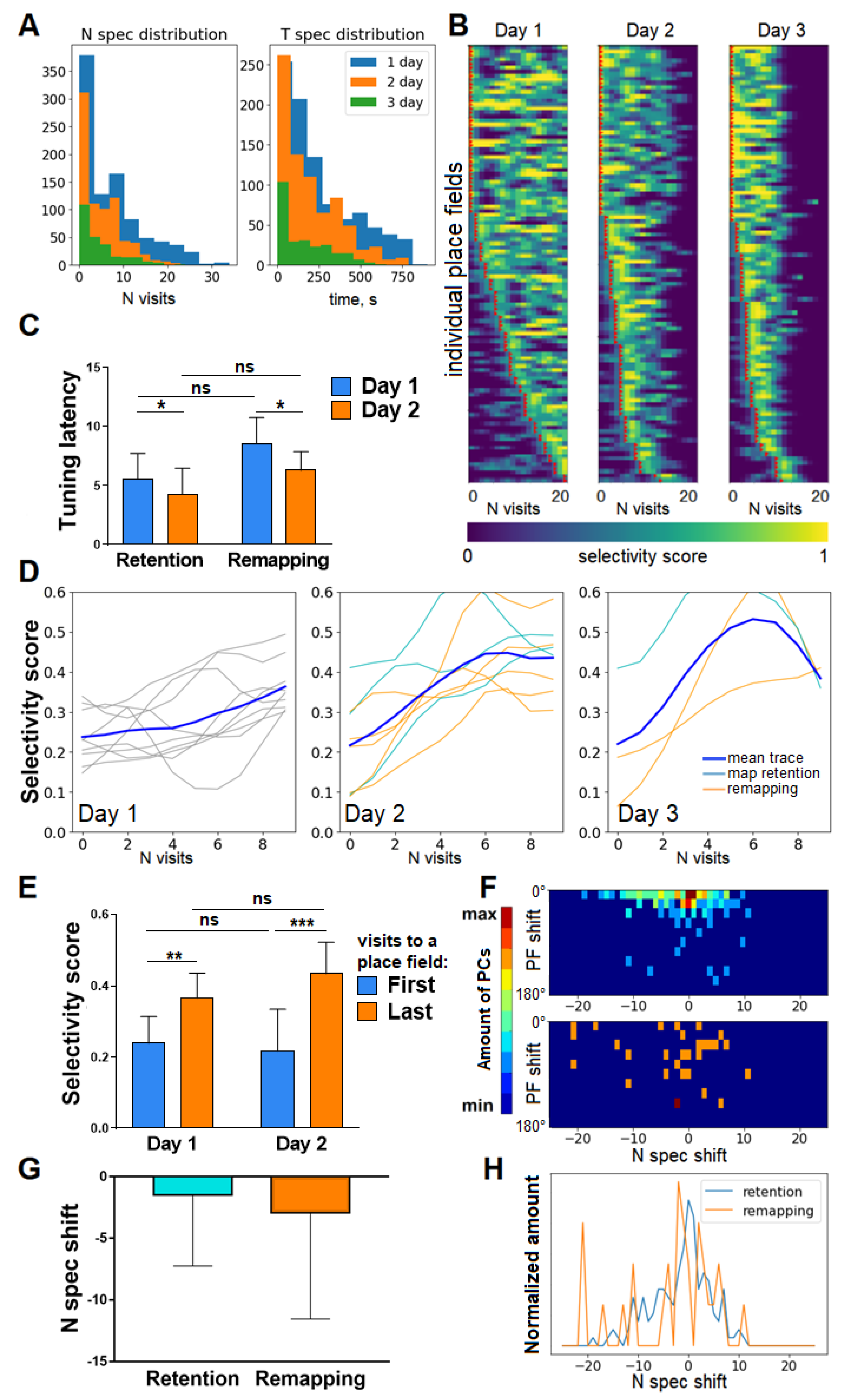
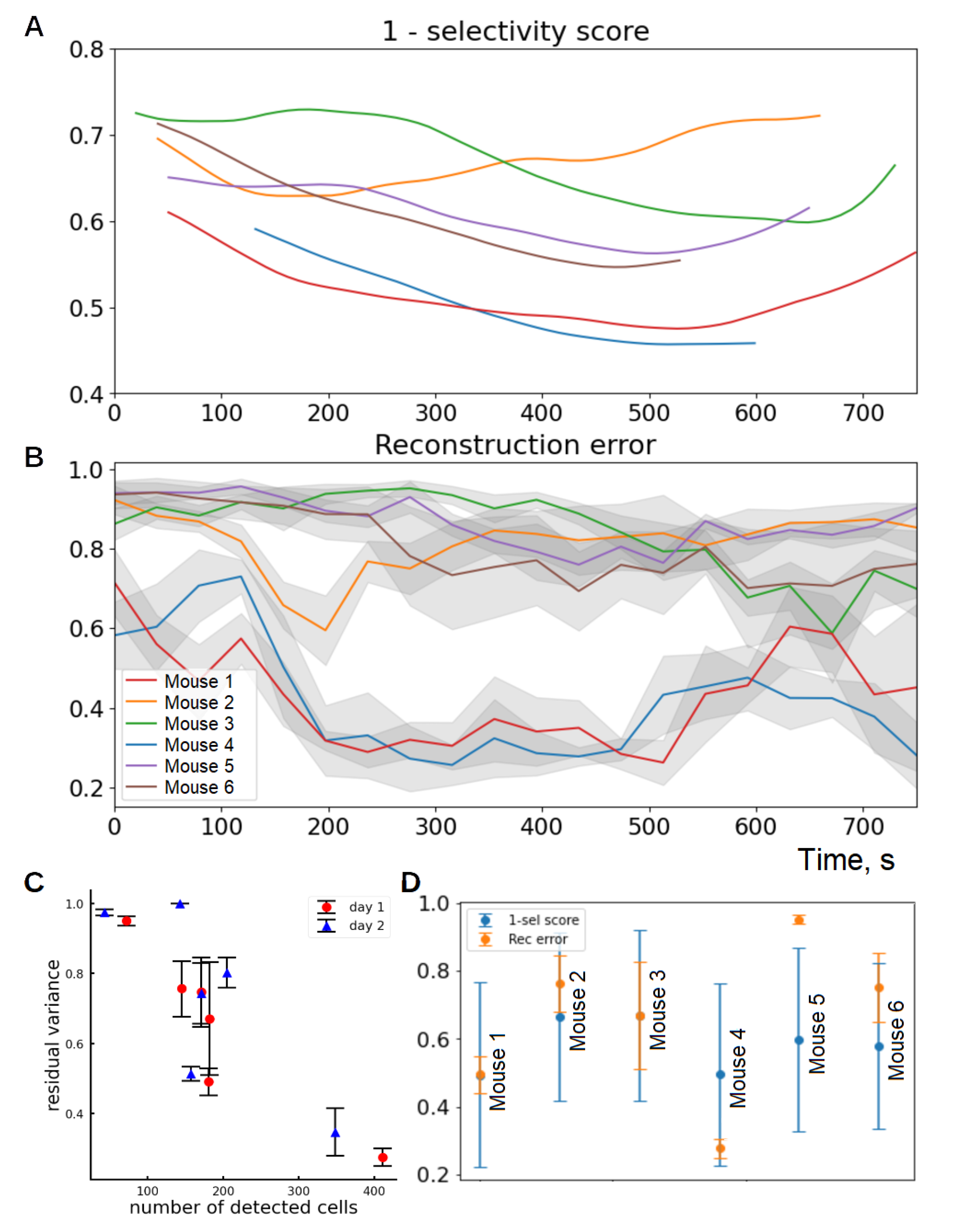
3.1. Selectivity Score and Tuning Latency
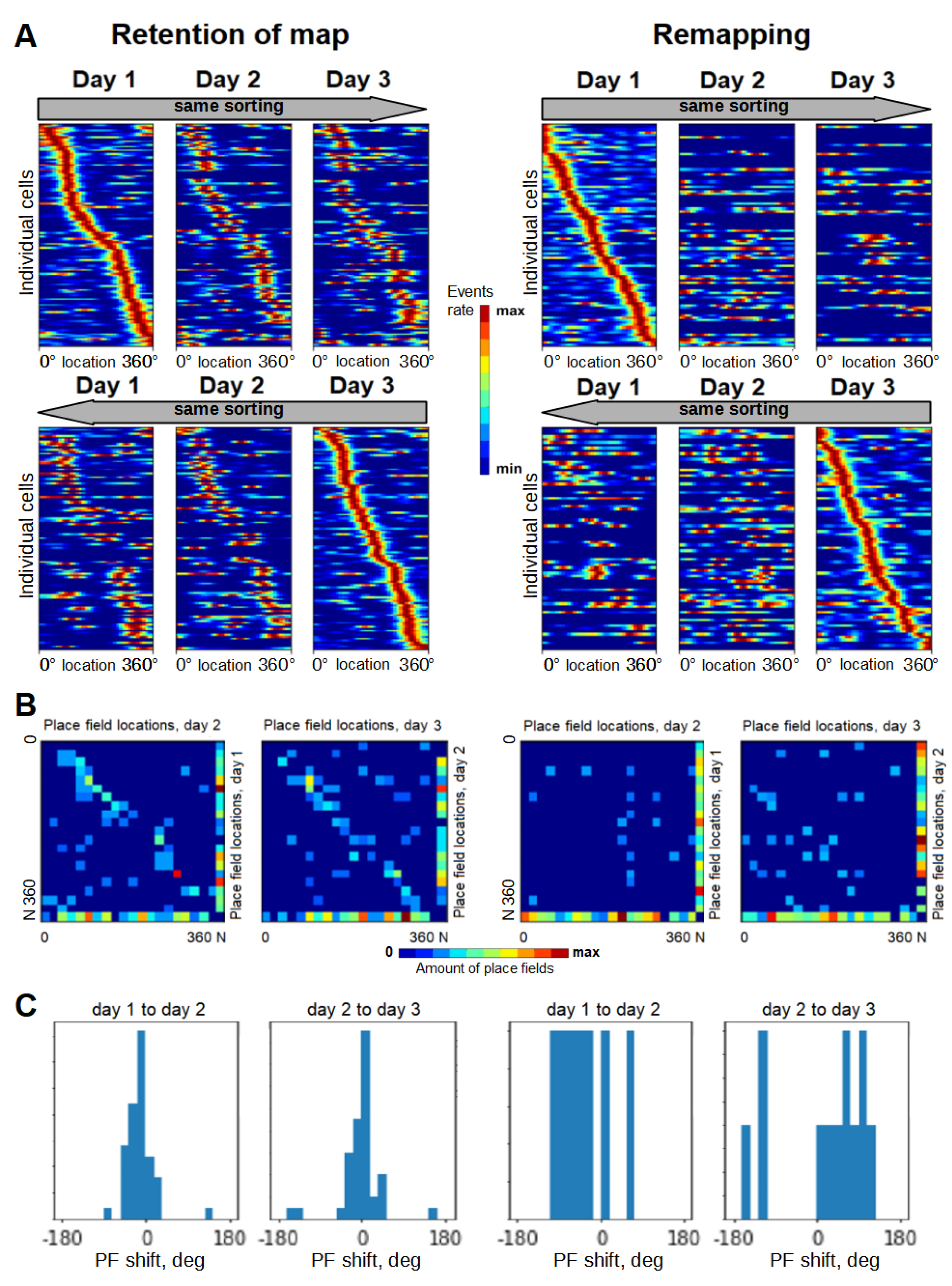
3.2. Selectivity Score Dynamics within Session and across Days
3.3. Nonlinear Dimensionality Reduction Reveals Track Geometry from Multidimensional Place Cell Activity
3.4. Dependence of Geometry Coding Quality on Population Size
3.5. Representation Quality Dynamics over Time
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Extended Data Tables and Figures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse | Session | Calcium Sensor | Cells | Place Cells | Place Fields | 1PF Cells | N Spec | T Spec | First Visit sel.sc. | Last Visit sel.sc. |
|---|---|---|---|---|---|---|---|---|---|---|
| Mouse 1 | 1 | GCaMP6s | 207 | 59 | 63 | 55 | 8.63 | 217.8 | 0.23 | 0.38 |
| 2 | 235 | 24 | 28 | 21 | 8.79 | 219.2 | 0.10 | 0.30 | ||
| 3 | 233 | 30 | 34 | 26 | 5.59 | 177.8 | 0.06 | 0.38 | ||
| Mouse 2 | 1 | GCaMP6s | 263 | 90 | 101 | 79 | 10.10 | 237.3 | 0.20 | 0.30 |
| 2 | 297 | 88 | 102 | 74 | 5.69 | 146.5 | 0.23 | 0.38 | ||
| 3 | 228 | 80 | 92 | 68 | 7.37 | 196.4 | 0.19 | 0.41 | ||
| Mouse 3 | 1 | GCaMP6s | 320 | 121 | 135 | 107 | 4.83 | 170.4 | 0.34 | 0.45 |
| 2 | 315 | 106 | 114 | 98 | 2.21 | 152.2 | 0.41 | 0.44 | ||
| 3 | 301 | 113 | 127 | 99 | 2.14 | 127.0 | 0.41 | 0.36 | ||
| Mouse 4 | 1 | GCaMP6s | 562 | 257 | 298 | 218 | 3.90 | 256.4 | 0.32 | 0.49 |
| 2 | 487 | 247 | 294 | 202 | 4.07 | 246.6 | 0.30 | 0.49 | ||
| Mouse 5 | 1 | NCaMP7 | 283 | 74 | 82 | 66 | 7.99 | 252.1 | 0.20 | 0.35 |
| 2 | 317 | 86 | 96 | 77 | 6.58 | 186.0 | 0.09 | 0.46 | ||
| Mouse 6 | 1 | NCaMP7 | 200 | 26 | 30 | 22 | 6.63 | 245.1 | 0.15 | 0.33 |
| 2 | 189 | 11 | 13 | 9 | 4.62 | 208.8 | 0.09 | 0.58 | ||
| Mouse 7 | 1 | GCaMP7f | 292 | 130 | 174 | 87 | 11.26 | 305.6 | 0.16 | 0.31 |
| 2 | 239 | 72 | 81 | 63 | 6.70 | 153.9 | 0.21 | 0.35 | ||
| Mouse 8 | 1 | FGCaMP7 | 95 | 10 | 10 | 10 | 6.10 | 144.4 | 0.31 | 0.31 |
| 2 | 123 | 35 | 38 | 32 | 5.68 | 102.4 | 0.30 | 0.47 | ||
| Mouse 9 | 1 | NCaMP7 | 103 | 12 | 19 | 7 | 7.26 | 271.5 | 0.23 | 0.35 |
| Mouse | Sessions Compared | Cells Matched | Retention p-Value | Retention of Map |
|---|---|---|---|---|
| Mouse 1 | 1 vs. 2 | 157 | 0.886 | no |
| 2 vs. 3 | 179 | 0.657 | no | |
| 1 vs. 3 | 153 | 0.839 | no | |
| Mouse 2 | 1 vs. 2 | 127 | 0.839 | no |
| 2 vs. 3 | 138 | 0.552 | no | |
| 1 vs. 3 | 116 | 0.006 | yes | |
| Mouse 3 | 1 vs. 2 | 229 | <0.001 | yes |
| 2 vs. 3 | 217 | <0.001 | yes | |
| 1 vs. 3 | 116 | <0.001 | yes | |
| Mouse 4 | 1 vs. 2 | 396 | <0.001 | yes |
| Mouse 5 | 1 vs. 2 | 208 | <0.001 | yes |
| Mouse 6 | 1 vs. 2 | 120 | 0.456 | no |
| Mouse 7 | 1 vs. 2 | 69 | 0.457 | no |
| Mouse 8 | 1 vs. 2 | 77 | 0.237 | no |


References
- O’Keefe, J.; Dostrovsky, J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971, 4, 171–175. [Google Scholar] [CrossRef]
- O’Keefe, J. Place units in the hippocampus of the freely moving rat. Exp. Neurol. 1976, 51, 78–109. [Google Scholar] [CrossRef]
- Ziv, Y.; Burns, L.; Cocker, E.; Hamel, E.; Ghosh, K.; Kitch, L.; El Gamal, A.; Schnitzer, M. Long-term dynamics of CA1 hippocampal place codes. Nat. Neurosci. 2013, 16, 264–266. [Google Scholar] [CrossRef]
- Rubin, A.; Geva, N.; Sheintuch, L.; Ziv, Y. Hippocampal ensemble dynamics timestamp events in long-term memory. Elife 2015, 4, e12247. [Google Scholar] [CrossRef]
- Kinsky, N.; Sullivan, D.; Mau, W.; Hasselmo, M.; Eichenbaum, H. Hippocampal Place Fields Maintain a Coherent and Flexible Map across Long Timescales. Curr. Biol. 2018, 28, 3578–3588. [Google Scholar] [CrossRef] [Green Version]
- Barnes, C.; Suster, M.; Shen, J.; McNaughton, B. Multistability of cognitive maps in the hippocampus of old rats. Nature 1997, 388, 272–275. [Google Scholar] [CrossRef]
- Sheintuch, L.; Geva, N.; Baumer, H.; Rechavi, Y.; Rubin, A.; Ziv, Y. Multiple Maps of the Same Spatial Context Can Stably Coexist in the Mouse Hippocampus. Curr. Biol. 2020, 30, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Madar, A.; Sheffield, M. Distinct place cell dynamics in CA1 and CA3 encode experience in new environments. Nat. Commun. 2021, 12, 2977. [Google Scholar] [CrossRef]
- Mehta, M.; Barnes, C.; McNaughton, B. Experience-dependent, asymmetric expansion of hippocampal place fields. Proc. Natl. Acad. Sci. USA 1997, 94, 8918–8921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, Y.; Igata, H.; Ikegaya, Y.; Sasaki, T. The Integration of Goal-Directed Signals onto Spatial Maps of Hippocampal Place Cells. Cell Rep. 2019, 27, 1516–1527. [Google Scholar] [CrossRef] [Green Version]
- Hok, V.; Lenck-Santini, P.; Roux, S.; Save, E.; Muller, R.; Poucet, B. Goal-related activity in hippocampal place cells. J. Neurosci. 2007, 27, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Burns, L.; Cocker, E.; Nimmerjahn, A.; Ziv, Y.; Gamal, A.; Schnitzer, M. Miniaturized integration of a fluorescence microscope. Nat. Methods 2011, 8, 871–878. [Google Scholar] [CrossRef] [Green Version]
- Barykina, N.; Sotskov, V.; Gruzdeva, A.; Wu, Y.; Portugues, R.; Subach, O.; Chefanova, E.; Plusnin, V.; Ivashkina, O.; Anokhin, K.; et al. FGCaMP7, an Improved Version of Fungi-Based Ratiometric Calcium Indicator for In Vivo Visualization of Neuronal Activity. Int. J. Mol. Sci. 2020, 21, 3012. [Google Scholar] [CrossRef]
- Subach, O.; Sotskov, V.; Plusnin, V.; Gruzdeva, A.; Barykina, N.; Ivashkina, O.; Anokhin, K.; Nikolaeva, A.; Korzhenevskiy, D.; Vlaskina, A.; et al. Novel Genetically Encoded Bright Positive Calcium Indicator NCaMP7 Based on the mNeonGreen Fluorescent Protein. Int. J. Mol. Sci. 2020, 21, 1644. [Google Scholar] [CrossRef] [Green Version]
- Gallego, J.; Perich, M.; Miller, L.; Solla, S. Neural Manifolds for the Control of Movement. Neuron 2017, 94, 978–984. [Google Scholar] [CrossRef]
- Gallego, J.; Perich, M.; Naufel, S.; Ethier, C.; Solla, S.; Miller, L. Cortical population activity within a preserved neural manifold underlies multiple motor behaviors. Nat. Commun. 2018, 9, 4233. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Cunningham, J.; Santhanam, G.; Ryu, S.; Shenoy, K.; Sahani, M. Gaussian-process factor analysis for low-dimensional single-trial analysis of neural population activity. J. Neurophysiol. 2009, 102, 614–635. [Google Scholar] [CrossRef]
- Barykina, N.; Subach, O.; Doronin, D.; Sotskov, V.; Roshchina, M.; Kunitsyna, T.; Malyshev, A.; Smirnov, I.; Azieva, A.; Sokolov, I.; et al. A new design for a green calcium indicator with a smaller size and a reduced number of calcium-binding sites. Sci. Rep. 2016, 6, 34447. [Google Scholar] [CrossRef]
- Barykina, N.; Doronin, D.; Subach, O.; Sotskov, V.; Plusnin, V.; Ivleva, O.; Gruzdeva, A.; Kunitsyna, T.; Ivashkina, O.; Lazutkin, A.; et al. NTnC-like genetically encoded calcium indicator with a positive and enhanced response and fast kinetics. Sci. Rep. 2018, 8, 15233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pnevmatikakis, E.; Giovannucci, A. NoRMCorre: An online algorithm for piecewise rigid motion correction of calcium imaging data. J. Neurosci. Methods 2017, 291, 83–94. [Google Scholar] [CrossRef]
- Lu, J.; Li, C.; Singh-Alvarado, J.; Zhou, Z.; Fröhlich, F.; Mooney, R.; Wang, F. MIN1PIPE: A Miniscope 1-Photon-Based Calcium Imaging Signal Extraction Pipeline. Cell Rep. 2018, 23, 3673–3684. [Google Scholar] [CrossRef]
- Sheintuch, L.; Rubin, A.; Brande-Eilat, N.; Geva, N.; Sadeh, N.; Pinchasof, O.; Ziv, Y. Tracking the Same Neurons across Multiple Days in Ca2+ Imaging Data. Cell Rep. 2017, 21, 1102–1115. [Google Scholar] [CrossRef] [Green Version]
- Lopes, G.; Bonacchi, N.; Frazão, J.; Neto, J.; Atallah, B.; Soares, S.; Moreira, L.; Matias, S.; Itskov, P.; Correia, P.; et al. Bonsai: An event-based framework for processing and controlling data streams. Front. Neuroinform. 2015, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Grijseels, D.; Shaw, K.; Barry, C.; Hall, C. Choice of method of place cell classification determines the population of cells identified. PLoS Comput. Biol. 2021, 17, e1008835. [Google Scholar] [CrossRef]
- Markus, E.; Barnes, C.; McNaughton, B.; Gladden, V.; Skaggs, W. Spatial information content and reliability of hippocampal CA1 neurons: Effects of visual input. Hippocampus 1994, 4, 410–421. [Google Scholar] [CrossRef]
- Belkin, M.; Niyogi, P. Laplacian eigenmaps for dimensionality reduction and data representation. Neural Comput. 2003, 15, 1373–1396. [Google Scholar] [CrossRef] [Green Version]
- Altman, N.; Krzywinski, M. The curse(s) of dimensionality. Nat. Methods 2018, 15, 399–400. [Google Scholar] [CrossRef]
- Chung, F. Spectral Graph Theory; American Mathematical Society: Philadelphia, PA, USA, 1997. [Google Scholar]
- Tenenbaum, J.; De Silva, V.; Langford, J. A global geometric framework for nonlinear dimensionality reduction. Science 2000, 290, 2319–2323. [Google Scholar] [CrossRef]
- Sotskov, V.; Plusnin, V.; Pospelov, N.; Anokhin, K. The Rapid Formation of CA1 Hippocampal Cognitive Map in Mice Exploring a Novel Environment. In Advances in Cognitive Research, Artificial Intelligence and Neuroinformatics. Intercognsci 2020. Advances in Intelligent Systems and Computing; Velichkovsky, B., Balaban, P., Ushakov, V., Eds.; Springer: Cham, Switzerland, 2021; pp. 452–457. [Google Scholar]
- Meshulam, L.; Gauthier, J.L.; Brody, C.D.; Tank, D.; Bialek, W. Collective Behavior of Place and Non-place Neurons in the Hippocampal Network. Neuron 2017, 96, 1178–1191. [Google Scholar] [CrossRef] [Green Version]
- Muller, R.; Kubie, J.; Ranck, J. Spatial firing patterns of hippocampal complex-spike cells in a fixed environment. J. Neurosci. 1987, 7, 1935–1950. [Google Scholar] [CrossRef] [PubMed]
- Latuske, P.; Kornienko, O.; Kohler, L.; Allen, K. Hippocampal Remapping and Its Entorhinal Origin. Front. Behav. Neurosci. 2018, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Q.; LeDuke, D.O.; Chua, K.; McDonald, R.J.; Sutherland, R.J. Relocating cued goals induces population remapping in CA1 related to memory performance in a two-platform water task in rats. Hippocampus 2018, 28, 431–440. [Google Scholar] [CrossRef]
- Mankin, E.A.; Sparks, F.T.; Slayyeh, B.; Sutherland, R.J.; Leutgeb, S.; Leutgeb, J.K. Neuronal code for extended time in the hippocampus. Proc. Natl. Acad. Sci. USA 2012, 109, 19462–19467. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, M.; Frank, L. Network dynamics underlying the formation of sparse, informative representations in the hippocampus. J. Neurosci. 2008, 28, 14271–14281. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Guo, W.; Zhang, J.; Newman, J.; Sun, S.; Wilson, M. Behavioral clusters revealed by end-to-end decoding from microendoscopic imaging. bioRxiv 2021. [Google Scholar] [CrossRef]
- Muller, R.; Bostock, E.; Taube, J.; Kubie, J. On the directional firing properties of hippocampal place cells. J. Neurosci. 1994, 14, 7235–7251. [Google Scholar] [CrossRef]
- McNaughton, B.; Barnes, C.; O’Keefe, J. The contributions of position, direction, and velocity to single unit activity in the hippocampus of freely-moving rats. Exp. Brain Res. 1983, 52, 41–49. [Google Scholar] [CrossRef]
- Scaplen, K.M.; Gulati, A.A.; Heimer-McGinn, V.L.; Burwell, R.D. Objects and landmarks: Hippocampal place cells respond differently to manipulations of visual cues depending on size, perspective, and experience. Hippocampus 2014, 24, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Kentros, C.; Agnihotri, N.; Streater, S.; Hawkins, R.; Kandel, E. Increased attention to spatial context increases both place field stability and spatial memory. Neuron 2004, 42, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Frank, L.; Brown, E.; Wilson, M. Trajectory encoding in the hippocampus and entorhinal cortex. Neuron 2000, 27, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Dragoi, G.; Tonegawa, S. Preplay of future place cell sequences by hippocampal cellular assemblies. Nature 2011, 469, 397–401. [Google Scholar] [CrossRef]
- Dragoi, G.; Tonegawa, S. Distinct preplay of multiple novel spatial experiences in the rat. Proc. Natl. Acad. Sci. USA 2013, 110, 9100–9105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosmark, A.; Buzsáki, G. Diversity in neural firing dynamics supports both rigid and learned hippocampal sequences. Science 2016, 351, 1440–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengio, Y.; Paiement, J.-F.; Vincent, P.; Delalleau, O.; Le Roux, N.; Ouimet, M. Out-of-sample extensions for LLE, Isomap, MDS, Eigenmaps, and Spectral Clustering. In Proceedings of the 16th International Conference on Neural Information Processing Systems (NIPS’03), Whistler, BC, Canada, 9–11 December 2003; MIT Press: Cambridge, MA, USA, 2003; pp. 177–184. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotskov, V.P.; Pospelov, N.A.; Plusnin, V.V.; Anokhin, K.V. Calcium Imaging Reveals Fast Tuning Dynamics of Hippocampal Place Cells and CA1 Population Activity during Free Exploration Task in Mice. Int. J. Mol. Sci. 2022, 23, 638. https://doi.org/10.3390/ijms23020638
Sotskov VP, Pospelov NA, Plusnin VV, Anokhin KV. Calcium Imaging Reveals Fast Tuning Dynamics of Hippocampal Place Cells and CA1 Population Activity during Free Exploration Task in Mice. International Journal of Molecular Sciences. 2022; 23(2):638. https://doi.org/10.3390/ijms23020638
Chicago/Turabian StyleSotskov, Vladimir P., Nikita A. Pospelov, Viktor V. Plusnin, and Konstantin V. Anokhin. 2022. "Calcium Imaging Reveals Fast Tuning Dynamics of Hippocampal Place Cells and CA1 Population Activity during Free Exploration Task in Mice" International Journal of Molecular Sciences 23, no. 2: 638. https://doi.org/10.3390/ijms23020638
APA StyleSotskov, V. P., Pospelov, N. A., Plusnin, V. V., & Anokhin, K. V. (2022). Calcium Imaging Reveals Fast Tuning Dynamics of Hippocampal Place Cells and CA1 Population Activity during Free Exploration Task in Mice. International Journal of Molecular Sciences, 23(2), 638. https://doi.org/10.3390/ijms23020638