
Frequency and Spectrum of Mutations Induced by Gamma Rays Revealed by Phenotype Screening and Whole-Genome Re-Sequencing in Arabidopsis thaliana


Abstract
:1. Introduction
2. Results
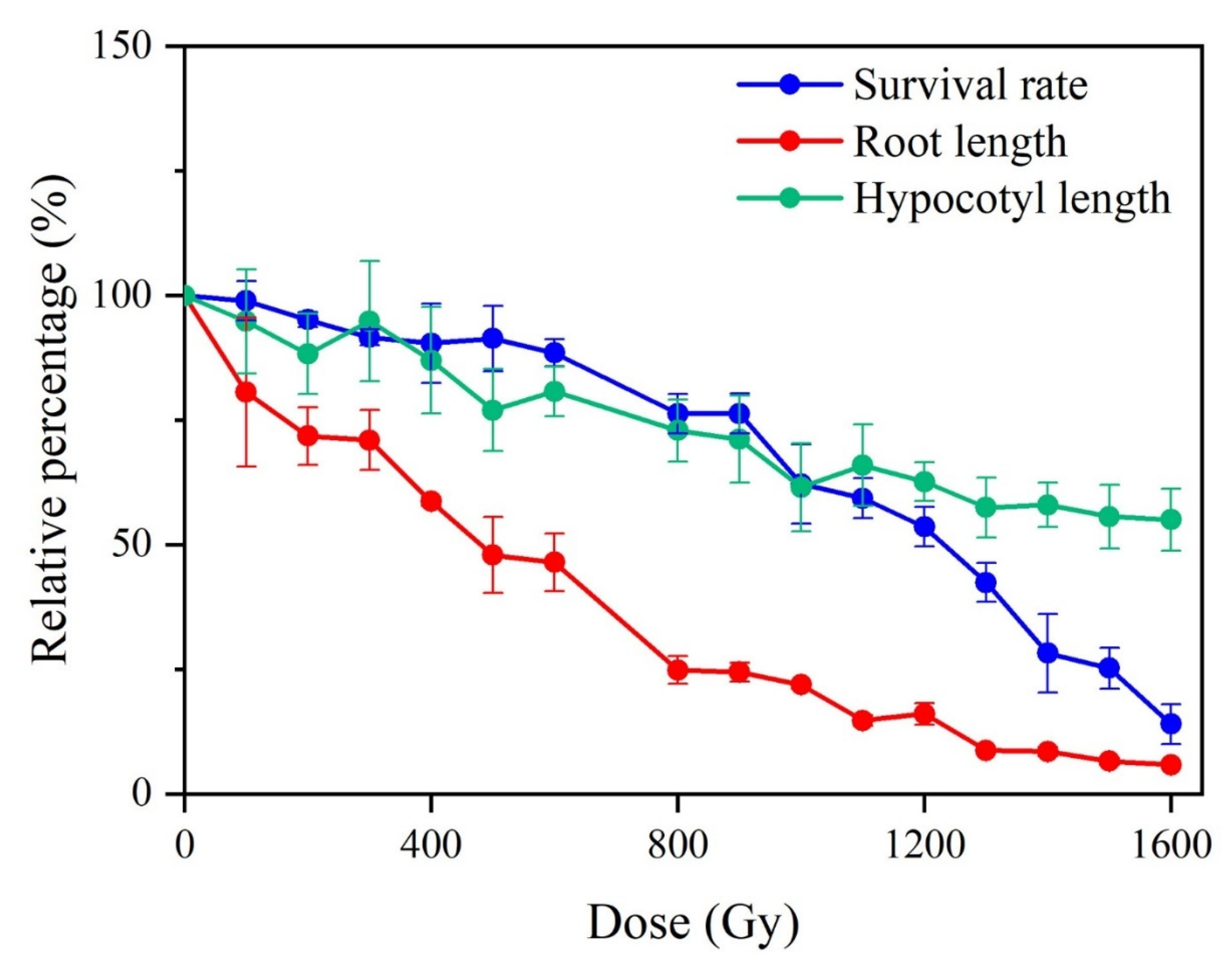
2.1. Biological Effects of Gamma Rays on M1 Plants
2.2. Spectrum and Frequency of Phenotype Mutations Identified in M2 Generation
2.3. Type and Frequency of Genomic Mutations Induced by Gamma Rays in M3 Generation
2.4. Distribution of Mutations on Genome
2.5. Characteristics of the SBSs and Small InDels Induced by Gamma Rays
2.6. Gene Affected by Gamma Rays
3. Discussion
3.1. Biological and Phenotypic Effects of Gamma Rays Irradiation on Arabidopsis thaliana
3.2. Genomic Mutation Profile of Mutations Induced by Gamma Rays Irradiation
3.3. Molecular Nature of Mutations Induced by Gamma Rays
3.4. Density of Affected Genes
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Growth Condition
5.2. Gamma Rays Irradiation, Plant Growth, Biological Effects on M1 Plants
5.3. Biological Effects on M1 Plants
5.4. Mutation Screening
5.5. Whole-Genome Re-Sequencing and Mutation Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qamar, Z.U.; Hameed, A.; Ashraf, M.; Rizwan, M.; Akhtar, M. Development and molecular characterization of low phytate basmati rice through induced mutagenesis, hybridization, backcross, and marker assisted breeding. Front. Plant Sci. 2019, 10, 1525. [Google Scholar] [CrossRef] [PubMed]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of ionizing radiation on biological molecules--mechanisms of damage and emerging methods of detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef] [PubMed]
- Pannunzio, N.R.; Watanabe, G.; Lieber, M.R. Nonhomologous DNA end-joining for repair of DNA double-strand breaks. J. Biol. Chem. 2018, 293, 10512–10523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.F.; Nagasawa, H.; Little, J.B.; Kato, T.A.; Shih, H.Y.; Xie, X.J.; Wilson, P.F.; Brogan, J.R.; Kurimasa, A.; Chen, D.J.; et al. Differential radiosensitivity phenotypes of DNA-PKcs mutations affecting NHEJ and HRR systems following irradiation with gamma-rays or very low fluences of alpha particles. PLoS ONE 2014, 9, e93579. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Ryu, T.H.; Lee, S.S.; Lee, S.; Chung, B.Y. Ionizing radiation manifesting DNA damage response in plants: An overview of DNA damage signaling and repair mechanisms in plants. Plant Sci. 2019, 278, 44–53. [Google Scholar] [CrossRef]
- Huefner, N.D.; Yoshiyama, K.; Friesner, J.D.; Conklin, P.A.; Britt, A.B. Genomic stability in response to high versus low linear energy transfer radiation in Arabidopsis thaliana. Front. Plant Sci. 2014, 5, 206. [Google Scholar] [CrossRef] [Green Version]
- Decottignies, A. Alternative end-joining mechanisms: A historical perspective. Front. Genet. 2013, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, R.; Nozawa, S.; Hase, Y.; Narumi, I.; Hidema, J.; Sakamoto, A.N. Mutational effects of gamma-rays and carbon ion beams on Arabidopsis. seedlings. J. Radiat. Res. 2013, 54, 1050–1056. [Google Scholar] [CrossRef]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional cloning of rice semidwarfing gene, sd-1: Rice “green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 2002, 9, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Shirasawa, K.; Hirakawa, H.; Nunome, T.; Tabata, S.; Isobe, S. Genome-wide survey of artificial mutations induced by ethyl methanesulfonate and gamma rays in tomato. Plant Biotechnol. J. 2016, 14, 51–60. [Google Scholar] [CrossRef]
- Xiao, X.O.; Lin, W.; Li, K.; Feng, X.; Jin, H.; Zou, H. Genome-wide analysis of artificial mutations induced by ethyl methanesulfonate in the eggplant (Solanum. melongena. L.). Genes 2019, 10, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Feng, H.; Du, Y.; Luo, S.; Li, W.; Yu, L.; Feng, Z.; Cui, T.; Zhou, L. Genetic polymorphisms in mutagenesis progeny of Arabidopsis thaliana irradiated by carbon-ion beams and gamma-rays irradiations. Int. J. Radiat. Biol. 2020, 96, 267–275. [Google Scholar] [CrossRef]
- Jo, Y.D.; Kim, S.H.; Hwang, J.E.; Kim, Y.S.; Kang, H.S.; Kim, S.W.; Kwon, S.J.; Ryu, J.; Kim, J.B.; Kang, S.Y. Construction of mutation populations by gamma-ray and carbon beam irradiation in chili pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2016, 57, 606–614. [Google Scholar] [CrossRef]
- Ulukapi, K.; Ozmen, S.F. Study of the effect of irradiation (Co-60) on M-1 plants of common bean (Phaseolus. vulgaris. L.) cultivars and determined of proper doses for mutation breeding. J. Radiat. Res. Appl. Sci. 2018, 11, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Lyu, J.I.; Hong, M.J.; Kim, D.G.; Kim, J.M.; Kim, J.B.; Eom, S.H.; Ha, B.K.; Kwon, S.J. Utility of TRAP markers to determine indel mutation frequencies induced by gamma-ray irradiation of faba bean (Vicia faba L.) seeds. Int. J. Radiat. Biol. 2019, 95, 1160–1171. [Google Scholar] [CrossRef]
- Singh, P.K.; Sadhukhan, R. EMS and gamma radiation induced mutation in grasspea (Lathyrus. sativus. L.). Legume Res. 2019, 42, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kim, S.W.; Ahn, J.W.; Ryu, J.; Kwon, S.J.; Kang, B.C.; Kim, J.B. Frequency, spectrum, and stability of leaf mutants induced by diverse gamma-ray treatments in two cymbidium hybrids. Plants 2020, 9, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setia, M.K.; Bala, M.; Singh, S. Induction of novel inflorescence traits in Chrysanthemum through (60)Co gamma irradiation. Int. J. Radiat. Biol. 2020, 96, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Kalpande, H.V.; Surashe, S.M.; Badignnavar, A.; More, A.; Ganapathi, T.R. Induced variability and assessment of mutagenic effectiveness and efficiency in sorghum genotypes [Sorghum bicolor (L.) Moench]. Int. J. Radiat. Biol. 2021, 1–23. [Google Scholar] [CrossRef]
- Naito, K.; Kusaba, M.; Shikazono, N.; Takano, T.; Tanaka, A.; Tanisaka, T.; Nishimura, M. Transmissible and nontransmissible mutations induced by irradiating Arabidopsis. thaliana. pollen with gamma-rays and carbon ions. Genetics 2005, 169, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Tavdumadze, K.R.; Topua, V.A. The effect of various doses of Co60 gamma-rays on the growth and development of Nicotiana tabacum L. Radiobiologiia 1967, 7, 475–476. [Google Scholar]
- Lee, M.B.; Kim, D.Y.; Jeon, W.B.; Hong, M.J.; Lee, Y.J.; Bold, O.; Jang, J.H.; Kim, Y.J.; Kang, S.Y.; Kim, D.S.; et al. Effect of gamma radiation on growth and lignin content in Brachypodium distachyon. J. Crop Sci. Biotechnol. 2013, 16, 105–110. [Google Scholar] [CrossRef]
- von Well, E.; Fossey, A.; Booyse, M. Efficiency of energy conversion and growth of gamma irradiated embryos and young seedlings of Triticum monococcum L. cultivar Einkorn. J. Radiat. Res. Appl. Sci. 2018, 11, 75–82. [Google Scholar] [CrossRef]
- Lee, J.-W.; Jo, I.-H.; Kim, J.-U.; Hong, C.-E.; Bang, K.-H.; Park, Y.-D. Determination of mutagenic sensitivity to gamma rays in ginseng (Panax ginseng) dehiscent seeds, roots, and somatic embryos. Hortic. Environ. Biotechnol. 2019, 60, 721–731. [Google Scholar] [CrossRef]
- Lavanya, H.N.; Mishra, S.; Anjanappa, M.; Aghora, T.S.; Sood, M.; Rao, K.V.; Reddy, A.B. Biological response, efficiency and effectiveness of gamma irradiation as mutagen on Pusa Navbahar cluster bean. Indian J. Hortic. 2020, 77, 478–483. [Google Scholar] [CrossRef]
- Perez-Jimenez, M.; Ignacio Tallon, C.; Perez-Tornero, O. Inducing mutations in Citrus spp.: Sensitivity of different sources of plant material to gamma radiation. Appl. Radiat. Isot. 2020, 157, 109030. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, S.Y.; Ryu, J.; Jo, Y.D.; Choi, H.-I.; Kim, J.-B.; Kang, S.-Y. Suggested doses of proton ions and gamma-rays for mutation induction in 20 plant species. Int. J. Radiat. Biol. 2021, 97, 1624–1629. [Google Scholar] [CrossRef]
- Kim, S.-M.; Jo, Y.D.; Chun, J.-I.; Kim, J.-B.; Kang, J.-H. Chronic Gamma Irradiation Changes Phenotype and Gene Expression Partially Transmitted to Next-Generation Tomato Seedlings. Agronomy 2021, 11, 1638. [Google Scholar] [CrossRef]
- Guo, W.; Comai, L.; Henry, I.M. Chromoanagenesis from radiation-induced genome damage in Populus. PLoS Genet. 2021, 17, e1009735. [Google Scholar] [CrossRef]
- Henry, I.M.; Zinkgraf, M.S.; Groover, A.T.; Comai, L. A system for dosage-based functional genomics in poplar. Plant Cell. 2015, 27, 2370–2383. [Google Scholar] [CrossRef] [Green Version]
- Hase, Y.; Satoh, K.; Seito, H.; Oono, Y. Genetic consequences of acute/chronic gamma and carbon ion irradiation of Arabidopsis. thaliana. Front. Plant. Sci. 2020, 11, 336. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.G.; Lee, S.C.; Lee, J.K.; Lee, J.W.; Kim, J.H.; Choi, S.Y.; Kim, J.B.; Choi, H.I.; Jang, C.S. Resequencing of a core rice mutant population induced by gamma-ray irradiation and its application in a genome-wide association study. J. Plant Biol. 2020, 63, 463–472. [Google Scholar] [CrossRef]
- Jo, D.Y.; Kim, J.-B. Frequency and spectrum of radiation-induced mutations revealed by whole-genome sequencing analyses of plants. Quantum Beam Sci. 2019, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Zhang, X.Q.; Wang, Y.; Wu, D.; Bellgard, M.I.; Xu, Y.; Shu, X.; Zhou, G.; Li, C. Characterization of genome-wide variations induced by gamma-ray radiation in barley using RNA-Seq. BMC Genom. 2019, 20, 783. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Luo, W.; Zhang, J.; Yan, X.; Du, Y.; Zhou, L.; Li, W.; Wang, H.; Chen, Z.; Guo, T. Genome-wide comparisons of mutations induced by carbon-ion beam and gamma-rays irradiation in rice via resequencing multiple mutants. Front. Plant Sci. 2019, 10, 1514. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Li, S.; Huang, J.; Fan, L.; Shu, Q. Identification and characterization of gamma-ray-induced mutations in rice cytoplasmic genomes by whole-genome sequencing. Cytogenet. Genome Res. 2020, 160, 100–109. [Google Scholar] [CrossRef]
- Lee, M.B.; Kim, D.Y.; Seo, Y.W. Identification of candidate genes for the seed coat colour change in a Brachypodium distachyon mutant induced by gamma radiation using whole-genome re-sequencing. Genome 2017, 60, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Jo, Y.D.; Kang, H.S.; Choi, H.-I.; Kim, J.-B. Genotype-by-Sequencing Analysis of Mutations and Recombination in Pepper Progeny of Gamma-Irradiated Gametophytes. Plants 2021, 10, 144. [Google Scholar] [CrossRef]
- Datta, S.; Jankowicz-Cieslak, J.; Nielen, S.; Ingelbrecht, I.; Till, B.J. Induction and recovery of copy number variation in banana through gamma irradiation and low-coverage whole-genome sequencing. Plant Biotechnol. J. 2018, 16, 1644–1653. [Google Scholar] [CrossRef] [Green Version]
- Hase, Y.; Satoh, K.; Kitamura, S.; Oono, Y. Physiological status of plant tissue affects the frequency and types of mutations induced by carbon-ion irradiation in Arabidopsis. Sci. Rep. 2018, 8, 1394. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shimizu, A.; Nishio, T.; Tsutsumi, N.; Kato, H. Comparison and characterization of mutations induced by gamma-ray and carbon-ion irradiation in rice (Oryza sativa L.) using whole-genome resequencing. G3-Genes Genomes Genet. 2019, 9, 3743–3751. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.G.; Kim, J.H.; Jang, C.S. A major QTL and a candidate gene for heading date in an early maturing rice mutant induced by gamma ray irradiation. Genes Genom. 2016, 38, 747–756. [Google Scholar] [CrossRef]
- Fiaz, S.; Ahmar, S.; Saeed, S.; Riaz, A.; Mora-Poblete, F.; Jung, K.-H. Evolution and Application of Genome Editing Techniques for Achieving Food and Nutritional Security. Int. J. Mol. Sci. 2021, 22, 5585. [Google Scholar] [CrossRef]
- Hyten, D.L.; Song, Q.; Zhu, Y.; Choi, I.Y.; Nelson, R.L.; Costa, J.M.; Specht, J.E.; Shoemaker, R.C.; Cregan, P.B. Impacts of genetic bottlenecks on soybean genome diversity. Proc. Natl. Acad. Sci. USA 2006, 103, 16666–16671. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.Y.; Ramachandran, S. Natural and artificial mutants as valuable resources for functional genomics and molecular breeding. Int. J. Biol. Sci. 2010, 6, 228–251. [Google Scholar] [CrossRef]
- Zhao, S.; Zheng, F.; He, W.; Wu, H.; Pan, S.; Lam, H.M. Impacts of nucleotide fixation during soybean domestication and improvement. BMC Plant Biol. 2015, 15, 81. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Li, W.; Yu, L.; Chen, G.; Liu, Q.; Luo, S.; Shu, Q.; Zhou, L. Mutagenic effects of carbon-ion irradiation on dry Arabidopsis thaliana seeds. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2014, 759, 28–36. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sheng, L.; Xu, Y.; Li, J.; Yang, Z.; Huang, H.; Xu, L. WOX11 and 12 Are Involved in the First-Step Cell Fate Transition during de Novo Root Organogenesis in Arabidopsis. Plant Cell 2014, 26, 1081–1093. [Google Scholar] [CrossRef] [Green Version]
- Shikazono, N.; Yokota, Y.; Kitamura, S.; Suzuki, C.; Watanabe, H.; Tano, S.; Tanaka, A. Mutation rate and novel tt mutants of Arabidopsis thaliana induced by carbon ions. Genetics 2003, 163, 1449–1455. [Google Scholar] [CrossRef]
- Gowthami, R.; Vanniarajan, C.; Souframanien, J.; Veni, K.; Renganathan, V.G. Efficiency of electron beam over gamma rays to induce desirable grain-type mutation in rice (Oryza sativa L.). Int. J. Radiat. Biol. 2021, 97, 727–736. [Google Scholar] [CrossRef]
- Cho, C.; Kim, K.-H.; Choi, M.S.; Chun, J.-B.; Seo, M.-S.; Jin, M.; Son, B.-y.; Kim, D.-Y. Characterization of gamma-radiation induced salt-tolerant silage maize mutant. Korean. J. Breed. Sci. 2019, 51, 318–325. [Google Scholar] [CrossRef]
- Shimizu, A.; Yamaguchi, H.; Degi, K.; Morishita, T. Development of ‘Darumadattan’, a semidwarf lodging-resistant Tartary buckwheat cultivar, using gamma-ray irradiation. Breed. Sci. 2020, 70, 623–630. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, S.; Huang, J.; Fu, H.; Zhou, L.; Furusawa, Y.; Shu, Q. Mutagenic effect of three ion beams on rice and identification of heritable mutations by whole genome sequencing. Plants 2020, 9, 551. [Google Scholar] [CrossRef]
- Oono, Y.; Ichida, H.; Morita, R.; Nozawa, S.; Satoh, K.; Shimizu, A.; Abe, T.; Kato, H.; Hase, Y. Genome sequencing of ion-beam-induced mutants facilitates detection of candidate genes responsible for phenotypes of mutants in rice. Mutat. Res. 2020, 821, 111691. [Google Scholar] [CrossRef]
- Onda, Y.; Mochida, K. Exploring genetic diversity in plants using high-throughput sequencing techniques. Curr. Genom. 2016, 17, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Jain, R.; Chern, M.; Pham, N.T.; Martin, J.A.; Wei, T.; Schackwitz, W.S.; Lipzen, A.M.; Duong, P.Q.; Jones, K.C.; et al. The sequences of 1504 mutants in the model rice variety Kitaake facilitate rapid functional genomic studies. Plant Cell 2017, 29, 1218–1231. [Google Scholar] [CrossRef] [Green Version]
- Kazama, Y.; Ishii, K.; Hirano, T.; Wakana, T.; Yamada, M.; Ohbu, S.; Abe, T. Different mutational function of low- and high-linear energy transfer heavy-ion irradiation demonstrated by whole-genome resequencing of Arabidopsis mutants. Plant J. 2017, 92, 1020–1030. [Google Scholar] [CrossRef] [Green Version]
- Hwang, W.J.; Kim, M.Y.; Kang, Y.J.; Shim, S.; Stacey, M.G.; Stacey, G.; Lee, S.H. Genome-wide analysis of mutations in a dwarf soybean mutant induced by fast neutron bombardment. Euphytica 2015, 203, 399–408. [Google Scholar] [CrossRef]
- Du, Y.; Hase, Y.; Satoh, K.; Shikazono, N. Characterization of gamma irradiation-induced mutations in Arabidopsis. mutants deficient in non-homologous end joining. J. Radiat. Res. 2020, 61, 639–647. [Google Scholar] [CrossRef]
- Belfield, E.J.; Gan, X.; Mithani, A.; Brown, C.; Jiang, C.; Franklin, K.; Alvey, E.; Wibowo, A.; Jung, M.; Bailey, K.; et al. Genome-wide analysis of mutations in mutant lineages selected following fast-neutron irradiation mutagenesis of Arabidopsis thaliana. Genome Res. 2012, 22, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Luo, S.; Yu, L.; Cui, T.; Chen, X.; Yang, J.; Li, X.; Li, W.; Wang, J.; Zhou, L. Strategies for identification of mutations induced by carbon-ion beam irradiation in Arabidopsis thaliana by whole genome re-sequencing. Mutat. Res. 2018, 807, 21–30. [Google Scholar] [CrossRef]
- Ishii, K.; Kazama, Y.; Hirano, T.; Hamada, M.; Ono, Y.; Yamada, M.; Abe, T. AMAP: A pipeline for whole-genome mutation detection in Arabidopsis thaliana. Genes Genet. Syst. 2017, 91, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Ichida, H.; Morita, R.; Shirakawa, Y.; Hayashi, Y.; Abe, T. Targeted exome sequencing of unselected heavy-ion beam-irradiated populations reveals less-biased mutation characteristics in the rice genome. Plant J. 2019, 98, 301–314. [Google Scholar] [CrossRef]
- Hirano, T.; Kazama, Y.; Ishii, K.; Ohbu, S.; Shirakawa, Y.; Abe, T. Comprehensive identification of mutations induced by heavy-ion beam irradiation in Arabidopsis thaliana. Plant J. 2015, 82, 93–104. [Google Scholar] [CrossRef]
- McVey, M.; Lee, S.E. MMEJ repair of double-strand breaks (director’s cut): Deleted sequences and alternative endings. Trends Genet. 2008, 24, 529–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, X.; Zhang, P.; Wang, Y. The Ku-dependent non-homologous end-joining but not other repair pathway is inhibited by high linear energy transfer ionizing radiation. DNA Repair. 2008, 7, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Hase, Y.; Yoshihara, R.; Nozawa, S.; Narumi, I. Mutagenic effects of carbon ions near the range end in plants. Mutat. Res. 2012, 731, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Luo, S.; Li, X.; Yang, J.; Cui, T.; Li, W.; Yu, L.; Feng, H.; Chen, Y.; Mu, J.; et al. Identification of Substitutions and Small Insertion-Deletions Induced by Carbon-Ion Beam Irradiation in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1851. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Chern, M.; Jain, R.; Martin, J.A.; Schackwitz, W.S.; Jiang, L.; Vega-Sanchez, M.E.; Lipzen, A.M.; Barry, K.W.; Schmutz, J.; et al. Genome-wide sequencing of 41 rice (Oryza sativa L.) mutated lines reveals diverse mutations induced by fast-neutron irradiation. Mol. Plant 2016, 9, 1078–1081. [Google Scholar] [CrossRef]
- Ossowski, S.; Schneeberger, K.; Lucas-Lledo, J.I.; Warthmann, N.; Clark, R.M.; Shaw, R.G.; Weigel, D.; Lynch, M. The rate and molecular spectrum of spontaneous mutations in Arabidopsis thaliana. Science 2010, 327, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Weng, M.L.; Becker, C.; Hildebrandt, J.; Neumann, M.; Rutter, M.T.; Shaw, R.G.; Weigel, D.; Fenster, C.B. Fine-grained analysis of spontaneous mutation spectrum and frequency in Arabidopsis thaliana. Genetics 2019, 211, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Addo-Quaye, C.; Buescher, E.; Best, N.; Chaikam, V.; Baxter, I.; Dilkes, B.P. Forward genetics by sequencing EMS variation-induced inbred lines. G3 Genes Genomes Genet. 2017, 7, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Greene, E.A.; Codomo, C.A.; Taylor, N.E.; Henikoff, J.G.; Till, B.J.; Reynolds, S.H.; Enns, L.C.; Burtner, C.; Johnson, J.E.; Odden, A.R.; et al. Spectrum of chemically induced mutations from a large-scale reverse-genetic screen in Arabidopsis. Genetics 2003, 164, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Cannistraro, V.J.; Taylor, J.-S. Synergistic modulation of cyclobutane pyrimidine dimer photoproduct formation and deamination at a TmCG site over a full helical DNA turn in a nucleosome core particle. Nucleic Acids Res. 2014, 42, 13122–13133. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data, P. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Koboldt, D.C.; Zhang, Q.Y.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Conesa, A.; Garcia-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Tung, N.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w(1118); iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, X.; Sherman, B.T.; Huang, D.W.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID-WS: A stateful web service to facilitate gene/protein list analysis. Bioinformatics 2012, 28, 1805–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | No. of Mutant | Frequency (%) | Brief Description |
|---|---|---|---|
| Leaf | 267 | 1.30 | |
| Lamina | 134 | 0.65 | The ratio of length to width, the edge and margin, surface of lamina; |
| Arrangement | 12 | 0.06 | Non-radial symmetry arranged rosette leaves, number of leaves |
| Petiole | 45 | 0.22 | Changes in the length of petioles |
| Vein | 13 | 0.06 | Obvious vein |
| Color | 63 | 0.31 | Yellow, variegated or albino leaves |
| Seeds viability | 36 | 0.18 | All the 18 seeds from single M2 family did not germinate |
| Stem | 29 | 0.14 | Dwarf |
| Fertility | 10 | 0.05 | Short or small siliques |
| Premature | 75 | 0.37 | Early bolting or flowering between 5–10 days than the Lab-WT |
| Total | 417 | 2.03 |
| Line | ID | Description |
|---|---|---|
| G200 | AT1G10170 | NF-X-like 1 (NFXL1) |
| AT1G13870 | KTI12-like, chromatin associated protein (DRL1) | |
| AT1G50600 | Scarecrow-like 5 (SCL5) | |
| AT2G17480 | Seven transmembrane MLO family protein (MLO8) | |
| AT2G33200 | F-box family protein | |
| AT3G02810 | Protein kinase superfamily protein | |
| AT3G05110 | Early endosome antigen-like protein, putative (DUF3444) | |
| AT3G09100 | mRNA capping enzyme family protein | |
| AT3G09750 | Galactose oxidase/kelch repeat superfamily protein | |
| AT3G09960 | Calcineurin-like metallo-phosphoesterase superfamily protein | |
| AT3G45060 | High affinity nitrate transporter 2.6 (NRT2.6) | |
| AT3G48140 | B12D protein | |
| AT3G57430 | Tetratricopeptide repeat (TPR)-like superfamily protein (OTP84) | |
| AT4G13190 | Protein kinase superfamily protein | |
| AT4G15200 | Formin 3 (FH3) | |
| AT4G23350 | Transmembrane protein, putative (DUF239) | |
| AT5G10620 | Methyltransferase | |
| AT5G25840 | DUF1677 family protein (DUF1677) | |
| AT5G43280 | Delta (3,5), delta (2,4)-dienoyl-CoA isomerase 1 (DCI1) | |
| G240 | AT1G10720 | BSD domain-containing protein |
| AT2G30520 | Phototropic-responsive NPH3 family protein (RPT2) | |
| AT2G32410 | AXR1-like protein (AXL) | |
| AT3G59350 | Protein kinase superfamily protein | |
| AT3G60700 | Hypothetical protein (DUF1163) | |
| AT4G04640 | ATPase, F1 complex, gamma subunit protein | |
| AT2G34420 | Photosystem II light harvesting complex protein B1B2 (LHB1B2) | |
| AT4G23660 | Polyprenyltransferase 1 (PPT1) | |
| G266 | AT1G60090 | Beta glucosidase 4 (BGLU4) |
| AT5G63100 | S-adenosyl-L-methionine-dependent methyltransferases superfamily protein | |
| G287 | AT3G01530 | Myb domain protein 57 (MYB57) |
| AT4G17140 | Pleckstrin homology (PH) domain-containing protein | |
| AT4G18110 | RING/U-box superfamily protein | |
| G320 | AT1G56360 | Purple acid phosphatase 6 (PAP6) |
| AT1G77020 | DNAJ heat shock N-terminal domain-containing protein | |
| AT2G01660 | Plasmodesmata-located protein 6 (PDLP6) | |
| AT2G29790 | Maternally expressed family protein | |
| AT3G20660 | Organic cation/carnitine transporter4 (OCT4) | |
| AT3G25727 | Non-LTR retrolelement reverse transcriptase | |
| AT3G56320 | PAP/OAS1 substrate-binding domain superfamily | |
| AT3G58300 | Phospholipase-like protein (PEARLI 4) family protein | |
| AT4G04880 | Adenosine/AMP deaminase family protein | |
| AT4G08350 | Global transcription factor group A2 (GTA2) | |
| AT4G26650 | RNA-binding (RRM/RBD/RNP motifs) family protein | |
| AT4G39190 | Nucleolar-like protein | |
| AT5G05820 | Nucleotide-sugar transporter family protein | |
| AT5G41460 | Transferring glycosyl group transferase (DUF604) | |
| AT5G50070 | Plant invertase/pectin methylesterase inhibitor superfamily protein | |
| AT5G51730 | RNA-binding (RRM/RBD/RNP motifs) family protein | |
| AT5G52570 | Beta-carotene hydroxylase 2 (BETA-OHASE 2) | |
| AT5G55970 | RING/U-box superfamily protein | |
| G431 | AT1G15490 | Alpha/beta-Hydrolases superfamily protein |
| AT1G70320 | Ubiquitin-protein ligase 2 (UPL2) | |
| AT3G18080 | B-S glucosidase 44 (BGLU44) | |
| AT3G18680 | Amino acid kinase family protein | |
| AT4G08580 | Microfibrillar-associated protein-like protein | |
| AT5G22580 | Stress responsive A/B Barrel Domain-containing protein | |
| AT5G22794 | Hypothetical protein | |
| G692 | AT1G09040 | Arginine-glutamic acid dipeptide repeat protein |
| AT1G44750 | Purine permease 11 (PUP11) | |
| AT2G15730 | P-loop containing nucleoside triphosphate hydrolases superfamily protein | |
| AT3G10015 | tRNA | |
| AT3G10720 | Plant invertase/pectin methylesterase inhibitor superfamily | |
| AT3G57910 | D111/G-patch domain-containing protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Feng, Z.; Wang, J.; Jin, W.; Wang, Z.; Guo, T.; Chen, Y.; Feng, H.; Yu, L.; Li, W.; et al. Frequency and Spectrum of Mutations Induced by Gamma Rays Revealed by Phenotype Screening and Whole-Genome Re-Sequencing in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 654. https://doi.org/10.3390/ijms23020654
Du Y, Feng Z, Wang J, Jin W, Wang Z, Guo T, Chen Y, Feng H, Yu L, Li W, et al. Frequency and Spectrum of Mutations Induced by Gamma Rays Revealed by Phenotype Screening and Whole-Genome Re-Sequencing in Arabidopsis thaliana. International Journal of Molecular Sciences. 2022; 23(2):654. https://doi.org/10.3390/ijms23020654
Chicago/Turabian StyleDu, Yan, Zhuo Feng, Jie Wang, Wenjie Jin, Zhuanzi Wang, Tao Guo, Yuze Chen, Hui Feng, Lixia Yu, Wenjian Li, and et al. 2022. "Frequency and Spectrum of Mutations Induced by Gamma Rays Revealed by Phenotype Screening and Whole-Genome Re-Sequencing in Arabidopsis thaliana" International Journal of Molecular Sciences 23, no. 2: 654. https://doi.org/10.3390/ijms23020654