
Relationships between Intraocular Pressure, Effective Filtration Area, and Morphological Changes in the Trabecular Meshwork of Steroid-Induced Ocular Hypertensive Mouse Eyes


Abstract
:1. Introduction
2. Results
2.1. DEX Treatment Elevates IOP
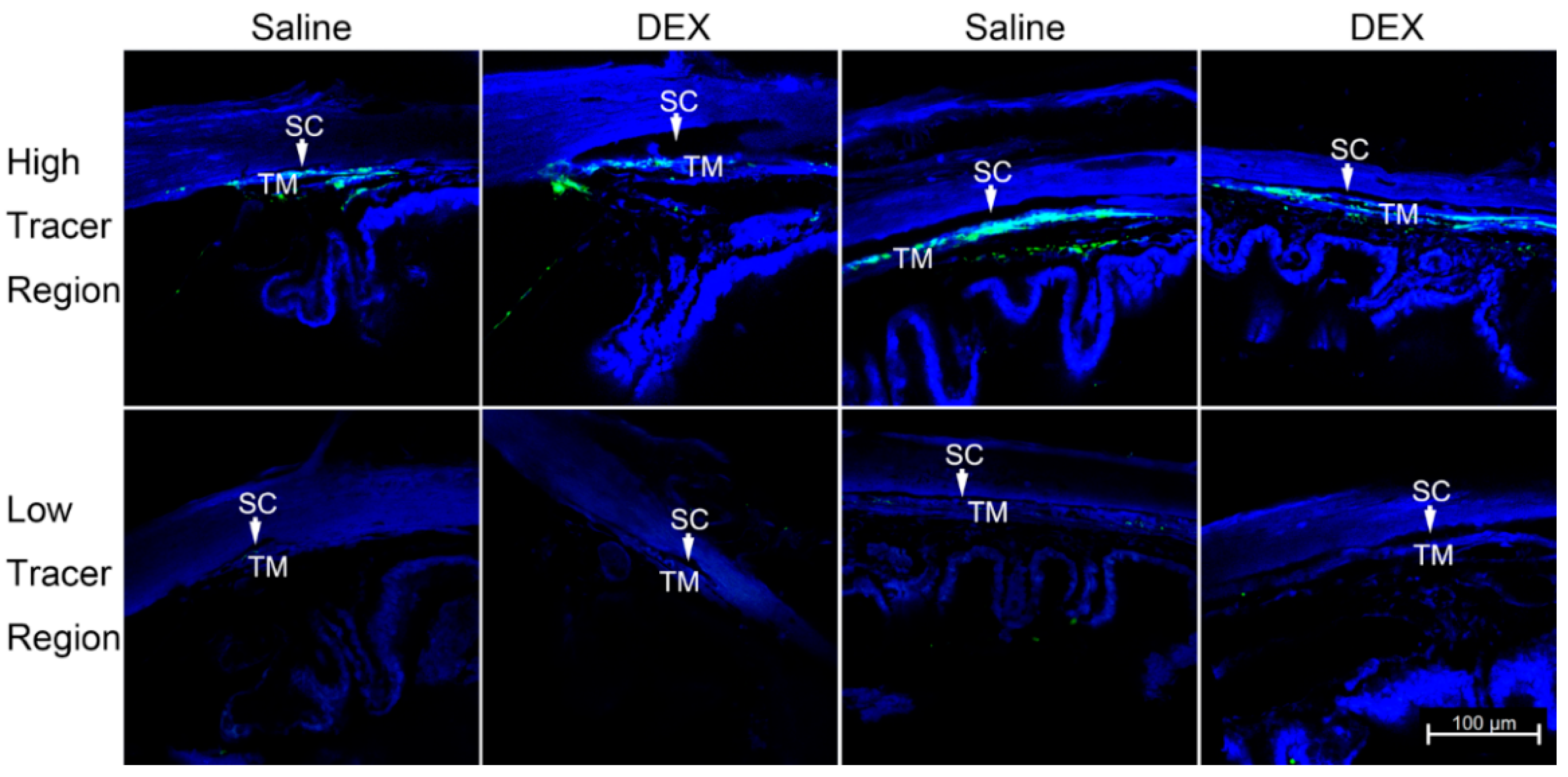
2.2. DEX Treatment Reduces PEFL
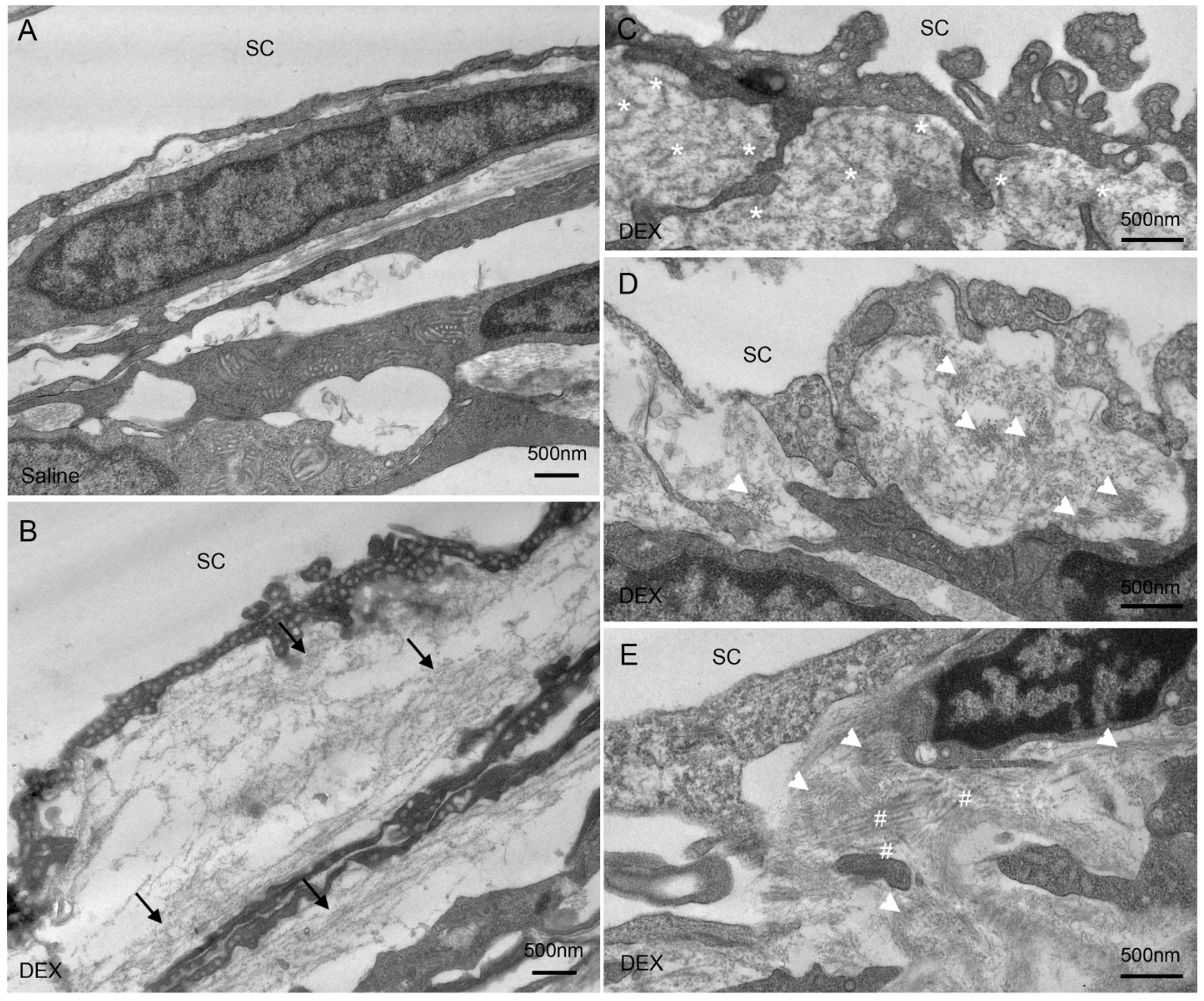
2.3. DEX Treatment-Induced Changes of Extracellular Matrix (ECM) in the JCT
2.4. DEX Treatment Decreased JCT Thickness
2.5. DEX Treatment Increased PBML of the IW of SC
2.6. Overall JCT Thickness and Overall PBML
2.7. Effects of DEX Treatment on Body Weight (BW)
3. Discussion
3.1. Correlations between IOP, TM PEFL, and PBML
3.2. Systemic Effects of Steroids Were Observed in Mice
3.3. Steroid-Induced Ultrastructural Changes in the TM Compared to Previous Studies
3.4. Mechanisms of Steroid-Induced IOP Elevation
4. Materials and Methods
4.1. Animal Husbandry
4.2. Eye Drops Administration
4.3. IOP Measurement
4.4. Tracer Injection
4.5. Confocal Microscopy
4.6. PEFL Analysis
4.7. Light and Electron Microscopy
4.8. Measurements of the JCT Thickness
4.9. Immunohistochemistry
4.10. Measurements of PBML of the IW of SC
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McLEAN, J.M.; Gordon, D.M.; Koteen, H. Clinical Experiences with ACTH and Cortisone in Ocular Diseases. Trans.-Am. Acad. Ophthalmol. Otolaryngol. Am. Acad. Ophthalmol. Otolaryngol. 1951, 55, 565–572. [Google Scholar]
- Grant, W.M. Experimental Aqueous Perfusion in Enucleated Human Eyes. Arch. Ophthalmol. 1963, 69, 783–801. [Google Scholar] [CrossRef]
- McMonnies, C.W. Intraocular Pressure Spikes in Keratectasia, Axial Myopia, and Glaucoma. Optom. Vis. Sci. 2008, 85, 1018–1026. [Google Scholar] [CrossRef]
- Johnson, D.; Gottanka, J.; Flügel, C.; Hoffmann, F.; Futa, R.; Lütjen-Drecoll, E. Ultrastructural Changes in the Trabecular Meshwork of Human Eyes Treated with Corticosteroids. Arch. Ophthalmol. 1997, 115, 375–383. [Google Scholar] [CrossRef]
- Zhou, L.; Li, Y.; Yue, B.Y. Glucocorticoid Effects on Extracellular Matrix Proteins and Integrins in Bovine Trabecular Meshwork Cells in Relation to Glaucoma. Int. J. Mol. Med. 1998, 1, 339–346. [Google Scholar] [CrossRef]
- Rönkkö, S.; Rekonen, P.; Kaarniranta, K.; Puustjärvi, T.; Teräsvirta, M.; Uusitalo, H. Matrix Metalloproteinases and Their Inhibitors in the Chamber Angle of Normal Eyes and Patients with Primary Open-Angle Glaucoma and Exfoliation Glaucoma. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Tamm, E.R.; Fuchshofer, R. What Increases Outflow Resistance in Primary Open-Angle Glaucoma? Surv. Ophthalmol. 2007, 52 (Suppl. 2), S101–S104. [Google Scholar] [CrossRef] [PubMed]
- Tektas, O.-Y.; Lütjen-Drecoll, E. Structural Changes of the Trabecular Meshwork in Different Kinds of Glaucoma. Exp. Eye Res. 2009, 88, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Vranka, J.A.; Kelley, M.J.; Acott, T.S.; Keller, K.E. Extracellular Matrix in the Trabecular Meshwork: Intraocular Pressure Regulation and Dysregulation in Glaucoma. Exp. Eye Res. 2015, 133, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Godel, V.; Feiler-Ofry, V.; Stein, R. Systemic Steroids and Ocular Fluid Dynamics. I. Analysis of the Sample as a Whole. Influence of Dosage and Duration of Therapy. Acta Ophthalmol. 1972, 50, 655–663. [Google Scholar] [CrossRef]
- Armaly, M.F.; Becker, B. Intraocular Pressure Response to Topical Corticosteroids. Fed. Proc. 1965, 24, 1274–1278. [Google Scholar]
- Armaly, M.F. Effect of corticosteroids on intraocular pressure and fluid dynamics. II. the effect of dexamethasone in the glaucomatous eye. Arch. Ophthalmol. 1963, 70, 492–499. [Google Scholar] [CrossRef]
- Lewis, J.M.; Priddy, T.; Judd, J.; Gordon, M.O.; Kass, M.A.; Kolker, A.E.; Becker, B. Intraocular Pressure Response to Topical Dexamethasone as a Predictor for the Development of Primary Open-Angle Glaucoma. Am. J. Ophthalmol. 1988, 106, 607–612. [Google Scholar] [CrossRef]
- Kitazawa, Y.; Horie, T. The Prognosis of Corticosteroid-Responsive Individuals. Arch. Ophthalmol. 1981, 99, 819–823. [Google Scholar] [CrossRef]
- Ticho, U.; Lahav, M.; Berkowitz, S.; Yoffe, P. Ocular Changes in Rabbits with Corticosteroid-Induced Ocular Hypertension. Br. J. Ophthalmol. 1979, 63, 646–650. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Gao, H.; Liu, H.; Sun, X. Metabolomics of Rabbit Aqueous Humor after Administration of Glucocorticosteroid. Curr. Eye Res. 2011, 36, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Bhattacherjee, P.; Paterson, C.A.; Spellman, J.M.; Graff, G.; Yanni, J.M. Pharmacological Validation of a Feline Model of Steroid-Induced Ocular Hypertension. Arch. Ophthalmol. 1999, 117, 361–364. [Google Scholar] [CrossRef] [Green Version]
- Gelatt, K.N.; Mackay, E.O. The Ocular Hypertensive Effects of Topical 0.1% Dexamethasone in Beagles with Inherited Glaucoma. J. Ocul. Pharmacol. Ther. 1998, 14, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Gerometta, R.; Podos, S.M.; Candia, O.A.; Wu, B.; Malgor, L.A.; Mittag, T.; Danias, J. Steroid-Induced Ocular Hypertension in Normal Cattle. Arch. Ophthalmol. 2004, 122, 1492–1497. [Google Scholar] [CrossRef] [Green Version]
- Gerometta, R.; Podos, S.M.; Danias, J.; Candia, O.A. Steroid-Induced Ocular Hypertension in Normal Sheep. Investig. Ophthalmol. Vis. Sci. 2009, 50, 669–673. [Google Scholar] [CrossRef] [Green Version]
- Fingert, J.H.; Clark, A.F.; Craig, J.E.; Alward, W.L.; Snibson, G.R.; McLaughlin, M.; Tuttle, L.; Mackey, D.A.; Sheffield, V.C.; Stone, E.M. Evaluation of the Myocilin (MYOC) Glaucoma Gene in Monkey and Human Steroid-Induced Ocular Hypertension. Investig. Ophthalmol. Vis. Sci. 2001, 42, 145–152. [Google Scholar]
- Sawaguchi, K.; Nakamura, Y.; Nakamura, Y.; Sakai, H.; Sawaguchi, S. Myocilin Gene Expression in the Trabecular Meshwork of Rats in a Steroid-Induced Ocular Hypertension Model. Ophthalmic Res. 2005, 37, 235–242. [Google Scholar] [CrossRef]
- Razali, N.; Agarwal, R.; Agarwal, P.; Kapitonova, M.Y.; Kannan Kutty, M.; Smirnov, A.; Salmah Bakar, N.; Ismail, N.M. Anterior and Posterior Segment Changes in Rat Eyes with Chronic Steroid Administration and Their Responsiveness to Antiglaucoma Drugs. Eur. J. Pharmacol. 2015, 749, 73–80. [Google Scholar] [CrossRef]
- Overby, D.R.; Bertrand, J.; Tektas, O.-Y.; Boussommier-Calleja, A.; Schicht, M.; Ethier, C.R.; Woodward, D.F.; Stamer, W.D.; Lütjen-Drecoll, E. Ultrastructural Changes Associated with Dexamethasone-Induced Ocular Hypertension in Mice. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4922–4933. [Google Scholar] [CrossRef] [Green Version]
- Patel, G.C.; Phan, T.N.; Maddineni, P.; Kasetti, R.B.; Millar, J.C.; Clark, A.F.; Zode, G.S. Dexamethasone-Induced Ocular Hypertension in Mice: Effects of Myocilin and Route of Administration. Am. J. Pathol. 2017, 187, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Whitlock, N.A.; McKnight, B.; Corcoran, K.N.; Rodriguez, L.A.; Rice, D.S. Increased Intraocular Pressure in Mice Treated with Dexamethasone. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6496–6503. [Google Scholar] [CrossRef] [Green Version]
- Zode, G.S.; Sharma, A.B.; Lin, X.; Searby, C.C.; Bugge, K.; Kim, G.H.; Clark, A.F.; Sheffield, V.C. Ocular-Specific ER Stress Reduction Rescues Glaucoma in Murine Glucocorticoid-Induced Glaucoma. J. Clin. Investig. 2014, 124, 1956–1965. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Lee, C.; Agrahari, V.; Wang, K.; Navarro, I.; Sherwood, J.M.; Crews, K.; Farsiu, S.; Gonzalez, P.; Lin, C.-W.; et al. In Vivo Measurement of Trabecular Meshwork Stiffness in a Corticosteroid-Induced Ocular Hypertensive Mouse Model. Proc. Natl. Acad. Sci. USA 2019, 116, 1714–1722. [Google Scholar] [CrossRef] [Green Version]
- Candia, O.A.; Gerometta, R.; Millar, J.C.; Podos, S.M. Suppression of Corticosteroid-Induced Ocular Hypertension in Sheep by Anecortave. Arch. Ophthalmol. 2010, 128, 338–343. [Google Scholar] [CrossRef] [Green Version]
- Danias, J.; Gerometta, R.; Ge, Y.; Ren, L.; Panagis, L.; Mittag, T.W.; Candia, O.A.; Podos, S.M. Gene Expression Changes in Steroid-Induced IOP Elevation in Bovine Trabecular Meshwork. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8636–8645. [Google Scholar] [CrossRef] [Green Version]
- François, J.; Benozzi, G.; Victoria-Troncoso, V.; Bohyn, W. Ultrastructural and Morphometric Study of Corticosteroid Glaucoma in Rabbits. Ophthalmic Res. 1984, 16, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Gerometta, R.; Kumar, S.; Shah, S.; Alvarez, L.; Candia, O.; Danias, J. Reduction of Steroid-Induced Intraocular Pressure Elevation in Sheep by Tissue Plasminogen Activator. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7903–7909. [Google Scholar] [CrossRef] [Green Version]
- Kasetti, R.B.; Maddineni, P.; Millar, J.C.; Clark, A.F.; Zode, G.S. Increased Synthesis and Deposition of Extracellular Matrix Proteins Leads to Endoplasmic Reticulum Stress in the Trabecular Meshwork. Sci. Rep. 2017, 7, 14951. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.; Tovar-Vidales, T.; Yorio, T.; Wordinger, R.J.; Clark, A.F. Perfusion-Cultured Bovine Anterior Segments as an Ex Vivo Model for Studying Glucocorticoid-Induced Ocular Hypertension and Glaucoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8068–8075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinzato, M.; Yamashiro, Y.; Miyara, N.; Iwamatsu, A.; Takeuchi, K.; Umikawa, M.; Bayarjargal, M.; Kariya, K.; Sawaguchi, S. Proteomic Analysis of the Trabecular Meshwork of Rats in a Steroid-Induced Ocular Hypertension Model: Downregulation of Type I Collagen C-Propeptides. Ophthalmic Res. 2007, 39, 330–337. [Google Scholar] [CrossRef]
- Tektas, O.-Y.; Hammer, C.M.; Danias, J.; Candia, O.; Gerometta, R.; Podos, S.M.; Lütjen-Drecoll, E. Morphologic Changes in the Outflow Pathways of Bovine Eyes Treated with Corticosteroids. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4060–4066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäepea, O.; Bill, A. The Pressures in the Episcleral Veins, Schlemm’s Canal and the Trabecular Meshwork in Monkeys: Effects of Changes in Intraocular Pressure. Exp. Eye Res. 1989, 49, 645–663. [Google Scholar] [CrossRef]
- Mäepea, O.; Bill, A. Pressures in the Juxtacanalicular Tissue and Schlemm’s Canal in Monkeys. Exp. Eye Res. 1992, 54, 879–883. [Google Scholar] [CrossRef]
- Vahabikashi, A.; Gelman, A.; Dong, B.; Gong, L.; Cha, E.D.K.; Schimmel, M.; Tamm, E.R.; Perkumas, K.; Stamer, W.D.; Sun, C.; et al. Increased Stiffness and Flow Resistance of the Inner Wall of Schlemm’s Canal in Glaucomatous Human Eyes. Proc. Natl. Acad. Sci. USA 2019, 116, 26555–26563. [Google Scholar] [CrossRef]
- Battista, S.A.; Lu, Z.; Hofmann, S.; Freddo, T.; Overby, D.R.; Gong, H. Reduction of the Available Area for Aqueous Humor Outflow and Increase in Meshwork Herniations into Collector Channels following Acute IOP Elevation in Bovine Eyes. Investig. Ophthalmol. Vis. Sci. 2008, 49, 5346–5352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Overby, D.R.; Scott, P.A.; Freddo, T.F.; Gong, H. The Mechanism of Increasing Outflow Facility by Rho-Kinase Inhibition with Y-27632 in Bovine Eyes. Exp. Eye Res. 2008, 86, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, P.A.; Lu, Z.; Liu, Y.; Gong, H. Relationships between Increased Aqueous Outflow Facility during Washout with the Changes in Hydrodynamic Pattern and Morphology in Bovine Aqueous Outflow Pathways. Exp. Eye Res. 2009, 89, 942–949. [Google Scholar] [CrossRef]
- Zhu, J.; Ye, W.; Gong, H. Development of a Novel Two Color Tracer Perfusion Technique for the Hydrodynamic Study of Aqueous Outflow in Bovine Eyes. Chin. Med. J. 2010, 123, 599–605. [Google Scholar] [PubMed]
- Lu, Z.; Zhang, Y.; Freddo, T.F.; Gong, H. Similar Hydrodynamic and Morphological Changes in the Aqueous Humor Outflow Pathway after Washout and Y27632 Treatment in Monkey Eyes. Exp. Eye Res. 2011, 93, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-Y.C.; Liu, Y.; Lu, Z.; Ren, R.; Gong, H. Effects of Y27632 on Aqueous Humor Outflow Facility with Changes in Hydrodynamic Pattern and Morphology in Human Eyes. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5859–5870. [Google Scholar] [CrossRef] [PubMed]
- Cha, E.D.K.; Xu, J.; Gong, L.; Gong, H. Variations in Active Outflow Along the Trabecular Outflow Pathway. Exp. Eye Res. 2016, 146, 354–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Mukherjee, D.; Navarro, I.; Ashpole, N.E.; Sherwood, J.M.; Chang, J.; Overby, D.R.; Yuan, F.; Gonzalez, P.; Kopczynski, C.C.; et al. Visualization of Conventional Outflow Tissue Responses to Netarsudil in Living Mouse Eyes. Eur. J. Pharmacol. 2016, 787, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Braakman, S.T.; Read, A.T.; Chan, D.W.-H.; Ethier, C.R.; Overby, D.R. Colocalization of Outflow Segmentation and Pores Along the Inner Wall of Schlemm’s Canal. Exp. Eye Res. 2015, 130, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.Y.H.; Folz, S.J.; Laryea, S.N.; Overby, D.R. Multi-Scale Analysis of Segmental Outflow Patterns in Human Trabecular Meshwork with Changing Intraocular Pressure. J. Ocul. Pharmacol. Ther. 2014, 30, 213–223. [Google Scholar] [CrossRef]
- Keller, K.E.; Bradley, J.M.; Vranka, J.A.; Acott, T.S. Segmental Versican Expression in the Trabecular Meshwork and Involvement in Outflow Facility. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5049–5057. [Google Scholar] [CrossRef] [Green Version]
- Vranka, J.A.; Staverosky, J.A.; Raghunathan, V.; Acott, T.S. Elevated Pressure Influences Relative Distribution of Segmental Regions of the Trabecular Meshwork. Exp. Eye Res. 2020, 190, 107888. [Google Scholar] [CrossRef]
- Swaminathan, S.S.; Oh, D.-J.; Kang, M.H.; Ren, R.; Jin, R.; Gong, H.; Rhee, D.J. Secreted Protein Acidic and Rich in Cysteine (SPARC)-Null Mice Exhibit More Uniform Outflow. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2035–2047. [Google Scholar] [CrossRef]
- Elena, P.-P.; Cimbolini, N.; Antonelli, S.; Bouzin, M.; Feraille, L.; Viaud, K.; Nicolas, M.-F.; Laurent, S.; Blot, C. Comparison of the Effect of Dexamethasone and Prednisolone Acetate in a Rat Model of Glucocorticoid-Induced Ocular Hypertension Following Topical Delivery. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2001. [Google Scholar]
- Smith, R.B.; Leano, N. Intraocular Pressure Following Pancuronium. Can. Anaesth. Soc. J. 1973, 20, 742–746. [Google Scholar] [CrossRef] [Green Version]
- Ivankovic, A.D.; Lowe, H.J. The Influence of Methoxyflurane and Neuroleptanesthesia on Intraocular Pressure in Man. Anesth. Analg. 1969, 48, 933–938. [Google Scholar] [CrossRef]
- Tang, L.Q.; Krupin, T.; Milner, M.; Woods, D.; Miller, K.; Carré, D.A.; Civan, M.M. Halogenated Inhalation Anesthetic Agents Decrease Transepithelial Electrical Measurements across the Isolated Iris-Ciliary Body. Investig. Ophthalmol. Vis. Sci. 1991, 32, 1912–1915. [Google Scholar]
- Ding, C.; Wang, P.; Tian, N. Effect of General Anesthetics on IOP in Elevated IOP Mouse Model. Exp. Eye Res. 2011, 92, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Cepurna, W.O.; Johnson, E.C.; Morrison, J.C. Effect of General Anesthetics on IOP in Rats with Experimental Aqueous Outflow Obstruction. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3415–3419. [Google Scholar]
- Bill, A. The Aqueous Humor Drainage Mechanism in the Cynomolgus Monkey (Macaca Irus) with Evidence for Unconventional Routes. Investig. Ophthalmol. 1965, 4, 911–919. [Google Scholar]
- Becker, B.; Neufeld, A.H. Pressure Dependence of Uveoscleral Outflow. J. Glaucoma 2002, 11, 545. [Google Scholar] [CrossRef]
- Kaufman, P. Some Thoughts on the Pressure Dependence of Uveoscleral Flow. J. Glaucoma 2003, 12, 89. [Google Scholar] [CrossRef]
- Bill, A. Some Thoughts on the Pressure Dependence of Uveoscleral Flow. J. Glaucoma 2003, 12, 88–89. [Google Scholar] [CrossRef]
- Aihara, M.; Lindsey, J.D.; Weinreb, R.N. Aqueous Humor Dynamics in Mice. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5168–5173. [Google Scholar] [CrossRef]
- Lei, Y.; Overby, D.R.; Boussommier-Calleja, A.; Stamer, W.D.; Ethier, C.R. Outflow Physiology of the Mouse Eye: Pressure Dependence and Washout. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1865–1871. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Li, G.; Le, T.D.; Kopczynski, C.; Stamer, W.D.; Gong, H. Netarsudil Increases Outflow Facility in Human Eyes Through Multiple Mechanisms. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6197–6209. [Google Scholar] [CrossRef]
- Steely, H.T.; Browder, S.L.; Julian, M.B.; Miggans, S.T.; Wilson, K.L.; Clark, A.F. The Effects of Dexamethasone on Fibronectin Expression in Cultured Human Trabecular Meshwork Cells. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2242–2250. [Google Scholar]
- Yemanyi, F.; Baidouri, H.; Burns, A.R.; Raghunathan, V. Dexamethasone and Glucocorticoid-Induced Matrix Temporally Modulate Key Integrins, Caveolins, Contractility, and Stiffness in Human Trabecular Meshwork Cells. Investig. Ophthalmol. Vis. Sci. 2020, 61, 16. [Google Scholar] [CrossRef]
- Rohen, J.W.; Linnér, E.; Witmer, R. Electron Microscopic Studies on the Trabecular Meshwork in Two Cases of Corticosteroid-Glaucoma. Exp. Eye Res. 1973, 17, 19–31. [Google Scholar] [CrossRef]
- Furuyoshi, N.; Furuyoshi, M.; Futa, R.; Gottanka, J.; Lütjen-Drecoll, E. Ultrastructural Changes in the Trabecular Meshwork of Juvenile Glaucoma. Ophthalmologica 1997, 211, 140–146. [Google Scholar] [CrossRef]
- Tektas, O.-Y.; Heinz, C.; Heiligenhaus, A.; Hammer, C.M.; Luetjen-Drecoll, E. Morphological Changes of Trabeculectomy Specimens in Different Kinds of Uveitic Glaucoma. Curr. Eye Res. 2011, 36, 442–448. [Google Scholar] [CrossRef]
- Tawara, A.; Tou, N.; Kubota, T.; Harada, Y.; Yokota, K. Immunohistochemical Evaluation of the Extracellular Matrix in Trabecular Meshwork in Steroid-Induced Glaucoma. Graefes Arch. Clin. Exp. Ophthalmol. 2008, 246, 1021–1028. [Google Scholar] [CrossRef]
- Gottanka, J.; Johnson, D.H.; Martus, P.; Lütjen-Drecoll, E. Severity of Optic Nerve Damage in Eyes with POAG Is Correlated with Changes in the Trabecular Meshwork. J. Glaucoma 1997, 6, 123–132. [Google Scholar] [CrossRef]
- Lütjen-Drecoll, E.; Shimizu, T.; Rohrbach, M.; Rohen, J.W. Quantitative Analysis of “plaque Material” in the Inner- and Outer Wall of Schlemm’s Canal in Normal- and Glaucomatous Eyes. Exp. Eye Res. 1986, 42, 443–455. [Google Scholar] [CrossRef]
- Grierson, I.; Lee, W.R.; Abraham, S.; Howes, R.C. Associations between the Cells of the Walls of Schlemm’s Canal. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 1978, 208, 33–47. [Google Scholar] [CrossRef]
- Gong, H.; Tripathi, R.C.; Tripathi, B.J. Morphology of the Aqueous Outflow Pathway. Microsc. Res. Tech. 1996, 33, 336–367. [Google Scholar] [CrossRef]
- Gong, H.; Ruberti, J.; Overby, D.; Johnson, M.; Freddo, T.F. A New View of the Human Trabecular Meshwork Using Quick-Freeze, Deep-Etch Electron Microscopy. Exp. Eye Res. 2002, 75, 347–358. [Google Scholar] [CrossRef]
- Leo, J.C.L.; Guo, C.; Woon, C.T.; Aw, S.E.; Lin, V.C.L. Glucocorticoid and Mineralocorticoid Cross-Talk with Progesterone Receptor to Induce Focal Adhesion and Growth Inhibition in Breast Cancer Cells. Endocrinology 2004, 145, 1314–1321. [Google Scholar] [CrossRef] [Green Version]
- Kasetti, R.B.; Maddineni, P.; Patel, P.; Searby, C.; Sheffield, V.C.; Zode, G.S. Transforming Growth Factor Β2 (TGFβ2) Signaling Plays a Key Role in Glucocorticoid-Induced Ocular Hypertension. J. Biol. Chem. 2018, 293, 9854–9868. [Google Scholar] [CrossRef] [Green Version]
- Fuchshofer, R.; Birke, M.; Welge-Lussen, U.; Kook, D.; Lütjen-Drecoll, E. Transforming Growth Factor-Beta 2 Modulated Extracellular Matrix Component Expression in Cultured Human Optic Nerve Head Astrocytes. Investig. Ophthalmol. Vis. Sci. 2005, 46, 568–578. [Google Scholar] [CrossRef] [Green Version]
- Zode, G.S.; Sethi, A.; Brun-Zinkernagel, A.-M.; Chang, I.-F.; Clark, A.F.; Wordinger, R.J. Transforming Growth Factor-Β2 Increases Extracellular Matrix Proteins in Optic Nerve Head Cells via Activation of the Smad Signaling Pathway. Mol. Vis. 2011, 17, 1745–1758. [Google Scholar]
- Paulsson, M. Basement Membrane Proteins: Structure, Assembly, and Cellular Interactions. Crit. Rev. Biochem. Mol. Biol. 1992, 27, 93–127. [Google Scholar] [CrossRef]
- Kalluri, R. Basement Membranes: Structure, Assembly and Role in Tumour Angiogenesis. Nat. Rev. Cancer 2003, 3, 422–433. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Week 0 | Week 1 | Week 2 | Week 3 | Week 4 | Week 5 |
|---|---|---|---|---|---|---|
| Saline | 14.1 ± 0.4 | 14.0 ± 0.3 | 13.5 ± 0.4 | 13.4 ± 0.3 | 13.6 ± 0.3 | 13.8 ± 0.3 |
| DEX | 13.8 ± 0.4 | 16.5 ± 0.7 | 16.8 ± 0.4 | 17.7 ± 0.5 | 17.8 ± 0.4 | 17.9 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, R.; Humphrey, A.A.; Swain, D.L.; Gong, H. Relationships between Intraocular Pressure, Effective Filtration Area, and Morphological Changes in the Trabecular Meshwork of Steroid-Induced Ocular Hypertensive Mouse Eyes. Int. J. Mol. Sci. 2022, 23, 854. https://doi.org/10.3390/ijms23020854
Ren R, Humphrey AA, Swain DL, Gong H. Relationships between Intraocular Pressure, Effective Filtration Area, and Morphological Changes in the Trabecular Meshwork of Steroid-Induced Ocular Hypertensive Mouse Eyes. International Journal of Molecular Sciences. 2022; 23(2):854. https://doi.org/10.3390/ijms23020854
Chicago/Turabian StyleRen, Ruiyi, Anne A. Humphrey, David L. Swain, and Haiyan Gong. 2022. "Relationships between Intraocular Pressure, Effective Filtration Area, and Morphological Changes in the Trabecular Meshwork of Steroid-Induced Ocular Hypertensive Mouse Eyes" International Journal of Molecular Sciences 23, no. 2: 854. https://doi.org/10.3390/ijms23020854