
Hemopexin and Cancer


Abstract
:1. Introduction
2. Hemopexin as a Cancer Biomarker
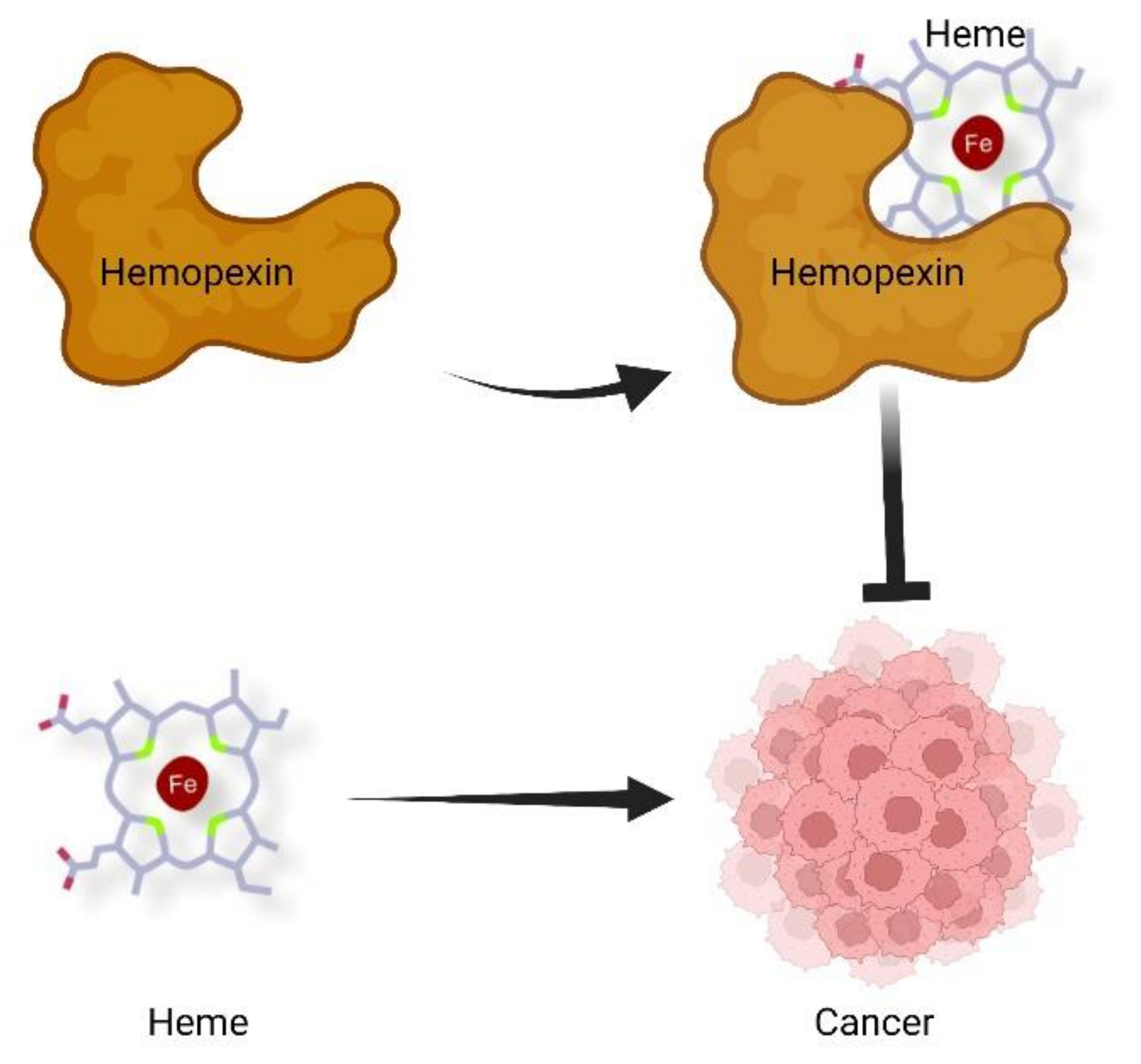
3. Hemopexin in Cancer Progression
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Immenschuh, S.; Song, D.X.; Satoh, H.; Muller-Eberhard, U. The type II hemopexin interleukin-6 response element predominates the transcriptional regulation of the hemopexin acute phase responsiveness. Biochem. Biophys. Res. Commun. 1995, 207, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Tolosano, E.; Altruda, F. Hemopexin: Structure, function, and regulation. DNA Cell Biol. 2002, 21, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Keel, S.B.; Doty, R.T.; Yang, Z.; Quigley, J.G.; Chen, J.; Knoblaugh, S.; Kingsley, P.D.; De Domenico, I.; Vaughn, M.B.; Kaplan, J.; et al. A heme export protein is required for red blood cell differentiation and iron homeostasis. Science 2008, 319, 825–828. [Google Scholar] [CrossRef] [Green Version]
- Tolosano, E.; Fagoonee, S.; Morello, N.; Vinchi, F.; Fiorito, V. Heme scavenging and the other facets of hemopexin. Antioxid. Redox Signal. 2010, 12, 305–320. [Google Scholar] [CrossRef]
- Fiorito, V.; Chiabrando, D.; Petrillo, S.; Bertino, F.; Tolosano, E. The Multifaceted Role of Heme in Cancer. Front. Oncol. 2020, 9, 1540. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Philips, J.D.; Doty, R.T.; Giraudi, P.; Ostrow, J.D.; Tiribelli, C.; Smith, A.; Abkowitz, J.L. Kinetics and specificity of feline leukemia virus subgroup C receptor (FLVCR) export function and its dependence on hemopexin. J. Biol. Chem. 2010, 285, 28874–28882. [Google Scholar] [CrossRef] [Green Version]
- Comunale, M.A.; Lowman, M.; Long, R.E.; Krakover, J.; Philip, R.; Seeholzer, S.; Evans, A.A.; Hann, H.W.; Block, T.M.; Mehta, A.S. Proteomic analysis of serum associated fucosylated glycoproteins in the development of primary hepatocellular carcinoma. J. Proteome Res. 2006, 5, 308–315. [Google Scholar] [CrossRef]
- Lin, Y.; Goedegebuure, P.S.; Tan, M.C.; Gross, J.; Malone, J.P.; Feng, S.; Larson, J.; Phommaly, C.; Trinkaus, K.; Townsend, R.R.; et al. Proteins associated with disease and clinical course in pancreas cancer: A proteomic analysis of plasma in surgical patients. J. Proteome Res. 2006, 5, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, T.M.; Hawke, D.H.; Liu, Y.; Krishnamurthy, S.; Fritsche, H.; Hunt, K.K.; Kuerer, H.M. Proteomic analysis of nipple aspirate fluid from women with early-stage breast cancer using isotope-coded affinity tags and tandem mass spectrometry reveals differential expression of vitamin D binding protein. BMC Cancer 2006, 6, 68. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Piñeiro, A.M.; Blanco-Prieto, S.; Sánchez-Otero, N.; Rodríguez-Berrocal, F.J.; de la Cadena, M.P. On the identification of biomarkers for non-small cell lung cancer in serum and pleural effusion. J. Proteom. 2010, 73, 1511–1522. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.; Huang, X.; Shen, Y.; Shen, J.; Ying, K. Differential proteome profiling of pleural effusions from lung cancer and benign inflammatory disease patients. Biochim. Biophys. Acta 2012, 1824, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, J.; Bowden, P.; Thavarajah, T.; Florentinus-Mefailoski, A.; Chen, Z.Z.; Tucholska, M.; Norzin, T.; Ho, M.T.; Phan, M.; Mohamed, N.; et al. The plasma peptides of ovarian cancer. Clin. Proteom. 2018, 15, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Takadate, T.; Mizuma, M.; Shima, H.; Suzuki, T.; Tachibana, T.; Shimura, M.; Hata, T.; Iseki, M.; Kawaguchi, K.; et al. Stromal expression of hemopexin is associated with lymph-node metastasis in pancreatic ductal adenocarcinoma. PLoS ONE 2020, 15, e0235904. [Google Scholar] [CrossRef]
- Canesin, G.; Di Ruscio, A.; Li, M.; Ummarino, S.; Hedblom, A.; Choudhury, R.; Krzyzanowska, A.; Csizmadia, E.; Palominos, M.; Stiehm, A.; et al. Scavenging of Labile Heme by Hemopexin Is a Key Checkpoint in Cancer Growth and Metastases. Cell Rep. 2020, 32, 108181. [Google Scholar] [CrossRef]
- Fiorito, V.; Allocco, A.L.; Petrillo, S.; Gazzano, E.; Torretta, S.; Marchi, S.; Destefanis, F.; Pacelli, C.; Audrito, V.; Provero, P.; et al. The heme synthesis-export system regulates the tricarboxylic acid cycle flux and oxidative phosphorylation. Cell Rep. 2021, 35, 109252. [Google Scholar] [CrossRef]
- Shen, J.; Sheng, X.; Chang, Z.; Wu, Q.; Wang, S.; Xuan, Z.; Li, D.; Wu, Y.; Shang, Y.; Kong, X.; et al. Iron metabolism regulates p53 signaling through direct heme-p53 interaction and modulation of p53 localization, stability, and function. Cell Rep. 2014, 7, 180–193. [Google Scholar] [CrossRef] [Green Version]
- Lignitto, L.; LeBoeuf, S.E.; Homer, H.; Jiang, S.; Askenazi, M.; Karakousi, T.R.; Pass, H.I.; Bhutkar, A.J.; Tsirigos, A.; Ueberheide, B.; et al. Nrf2 Activation Promotes Lung Cancer Metastasis by Inhibiting the Degradation of Bach1. Cell 2019, 178, 316–329.e318. [Google Scholar] [CrossRef]
- Wiel, C.; Le Gal, K.; Ibrahim, M.X.; Jahangir, C.A.; Kashif, M.; Yao, H.; Ziegler, D.V.; Xu, X.; Ghosh, T.; Mondal, T.; et al. BACH1 Stabilization by Antioxidants Stimulates Lung Cancer Metastasis. Cell 2019, 178, 330–345.e322. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yesilkanal, A.E.; Wynne, J.P.; Frankenberger, C.; Liu, J.; Yan, J.; Elbaz, M.; Rabe, D.C.; Rustandy, F.D.; Tiwari, P.; et al. Effective breast cancer combination therapy targeting BACH1 and mitochondrial metabolism. Nature 2019, 568, 254–258. [Google Scholar] [CrossRef]
- Costa da Silva, M.; Breckwoldt, M.O.; Vinchi, F.; Correia, M.P.; Stojanovic, A.; Thielmann, C.M.; Meister, M.; Muley, T.; Warth, A.; Platten, M.; et al. Iron Induces Anti-tumor Activity in Tumor-Associated Macrophages. Front. Immunol. 2017, 8, 1479. [Google Scholar] [CrossRef] [Green Version]
- Kayama, H.; Kohyama, M.; Okuzaki, D.; Motooka, D.; Barman, S.; Okumura, R.; Muneta, M.; Hoshino, K.; Sasaki, I.; Ise, W.; et al. Heme ameliorates dextran sodium sulfate-induced colitis through providing intestinal macrophages with noninflammatory profiles. Proc. Natl. Acad. Sci. USA 2018, 115, 8418–8423. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Tumor | Study Design | Sample | Observations | References |
|---|---|---|---|---|
| Hepatocellular carcinoma | Patients vs. healthy individuals | Serum | Presence of fucosylated Hx in patients | [7] |
| Pancreatic tumor | Surgical patients before/after tumor resection | Plasma | Positive correlation between Hx levels and tumor burden | [8] |
| Breast carcinoma | Tumor bearing breasts vs. disease free breasts | Nipple aspirate fluid | Hx over-expression in tumor-bearing breasts | [9] |
| Non-small cell lung cancer | Cancer patients vs. pneumonia patients | Serum | Higher Hx levels in cancer patients | [10] |
| Non-small cell lung cancer | Cancer patients vs. tuberculosis patients | Pleuraleffusion | Higher Hx levels in cancerpatients | [10] |
| Lung adenocarcinoma | Cancer patients vs. tuberculosis or pneumonia patients | Pleural effusion | Higher Hx levels in cancer patients | [11] |
| Ovarian cancer | Cancer patients vs. controls | Plasma | Higher Hx levels in ovarian cancer patients | [12] |
| Pancreatic ductal adenocarcinoma | LN+ PDAC vs. LN- PDAC | Stromal fibroblasts, cancer cell cytosol, lymphocytes, macrophages | Higher Hx levels in LN+ PDAC | [13] |
| Pancreatic ductal adenocarcinoma | LN+ PDAC | Tumor stroma | Positive correlation between Hx levels and High venous/lymphatic invasion | [13] |
| Prostate cancer | Patients vs. healthy individuals | Plasma | Lower Hx levels in cancer patients | [14] |
| Prostate cancer | Tumor vs. benign tissues | Stroma | Lower Hx levels in tumor stroma | [14] |
| Prostate cancer | Normal tissues vs. tumor-adjacent tissues vs. primary prostate tumors vs. PCa metastasis | Tissues | Higher Hx mRNA levels in metastasis | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiorito, V.; Tolosano, E. Hemopexin and Cancer. Int. J. Mol. Sci. 2022, 23, 997. https://doi.org/10.3390/ijms23020997
Fiorito V, Tolosano E. Hemopexin and Cancer. International Journal of Molecular Sciences. 2022; 23(2):997. https://doi.org/10.3390/ijms23020997
Chicago/Turabian StyleFiorito, Veronica, and Emanuela Tolosano. 2022. "Hemopexin and Cancer" International Journal of Molecular Sciences 23, no. 2: 997. https://doi.org/10.3390/ijms23020997
APA StyleFiorito, V., & Tolosano, E. (2022). Hemopexin and Cancer. International Journal of Molecular Sciences, 23(2), 997. https://doi.org/10.3390/ijms23020997