Abstract
Periodontitis is a chronic non-communicable disease caused by dysbiotic changes that affect the subgingival microbiota. During periodontitis, neutrophils play a central role in the initial recognition of bacteria, and their number increases with the appearance of the first signs of periodontal inflammation. Recent evidence has led to the proposition that neutrophils can also functionally polarize, determining selective activity patterns related to different diseases. Two well-defined neutrophil phenotypes have been described, the pro-inflammatory N1 subset and the suppressor N2 subset. To date, it has not been established whether these different neutrophil subtypes play a role in the pathogenesis of periodontitis. Thus, this scoping review aimed to determine whether there was evidence to suggest that the neutrophils present in periodontal tissues can be associated with certain phenotypes. The research question, population, concept, and context sought to identify original articles, in humans, that detected the presence of neutrophils in the periodontal tissues of people affected by periodontitis. Based on the search strategy, we found 3658 studies. After removing the papers with abstracts not related to the outcome measures and eligibility criteria, 16 articles were included for qualitative analysis. Several studies identified the presence of different neutrophil subsets, specifically, the naive, pro- and para-inflammatory, hyper-reactive and hyper-active, and high- and low-responder phenotypes. The existing evidence demonstrates the presence of pro-inflammatory, hyper-reactive and high-responder neutrophils in periodontal tissues affected with periodontitis. There is no evidence demonstrating the presence of the N1 or N2 phenotypes in periodontal tissues during periodontitis. However, the existence of pro-inflammatory phenotypes, which increase NETosis and degranulation, and increase the production of pro-inflammatory cytokines, could be suggestive of the N1 phenotypes.
1. Introduction
Neutrophils are polymorphonuclear cells derived from bone marrow granulocyte precursors [1]. Neutrophils have a relatively short lifespan in the peripheral blood circulation, approximately 4 to 5 h. However, a longer lifespan is evidenced when they infiltrate the connective tissues, where they recognize microbiota- or pathogen-associated molecular patterns (MAMPs or PAMPs). Neutrophils are between 60 to 70% of the total polymorphonuclear cells [2]. Neutrophils are the first cell population to be recruited at inflamed sites, and can migrate by chemotaxis from the peripheral blood circulation to the tissues. In the connective tissues, they fulfill their main role: phagocytosis [3,4]. In recent years, in tumor-related pathologies, two subtypes of neutrophils have been defined, with phenotypic and functional differences: neutrophil type 1, or anti-tumorigenic N1, and N2, or pro-tumorigenic neutrophils [5,6]. Additionally, these same phenotypes have been identified as pro-inflammatory (N1) and immune modulatory (N2), based on their abilities to degranulate, release cytokines, and migrate.
The TAN differentiate in the presence of interferon (IFN) type I toward the N1 subset, which is characterized by increasing adhesion, transmigration, phagocytosis, oxidative burst, degranulation, and neutrophil extracellular trap (NET) release, also known as NETosis, as well as cluster of differentiation (CD)-177 over-expression [5]. Otherwise, in the presence of high concentrations of transforming growth factor-β (TGF-β), neutrophils polarize into a modulatory phenotype, fulfilling an antagonistic role to the N1 phenotype, and N2, which also increases the expression of Olfactomedin 4 (OLFM4) [7]. Thus, the phenotypic shift from N1 to N2 may therefore suggest antagonistic signaling pathways between TGF-β and IFN type 1, with a different role during health or cancer [8]. Recently, some experimental studies in rodents have detected a similar distribution of neutrophil subsets in both heart and brain affected with inflammatory diseases.
Additionally, after a heart or brain infarction, neutrophils infiltrate the tissues rapidly after an ischemic event [9]. It was observed that during the first 24 h after ischemia, neutrophil infiltration is essential for the inflammatory response [10,11]. Additionally, it was observed that N2 neutrophils increase over time, and the decrease in the N1/N2 ratio is directly associated with the resolution of inflammation [9]. In fact, in animals depleted of neutrophils and affected by brain ischemia, it was observed that the size of the lesion is smaller compared to animals with neutrophils [12]. In general terms, both in the heart and in the brain after ischemia, the pro-inflammatory milieu triggers the polarization of N1 neutrophils [9,10,11,12,13,14]. The N1 phenotypes increase the pro-inflammatory response by secreting a higher amount of IL-1β, IL-6, TNF-α, and IFN-γ, by producing reactive oxygen species (ROS), and ultimately through NETosis [3,4,15].
Although the TAN1 and TAN2 response has been studied extensively, the possible role of neutrophil phenotypes in other pathologies has only recently been explored in animal models. To date, it has not been identified in chronic inflammatory diseases such as periodontal diseases. Therefore, this scoping review aims to identify whether there is evidence to suggest the presence of these neutrophil subsets in periodontal tissues affected by periodontitis.
2. Results
2.1. Search Results
Initially, 3658 studies were identified. After removing the papers with abstracts not related to the primary, secondary or tertiary outcome measures, a total of 147 potentially eligible abstracts were selected. When applying the eligibility criteria, 101 articles were selected, and 46 were excluded. After eliminating the duplicates, a full-text analysis of the studies was performed, and, finally, 14 articles were included for qualitative analysis (Figure 1). The studies included in the selection corresponded to four descriptive [16,17,18,19], four cohort [20,21,22,23], and six case–control [24,25,26,27,28,29] studies. The full data summaries are presented in Table 1.
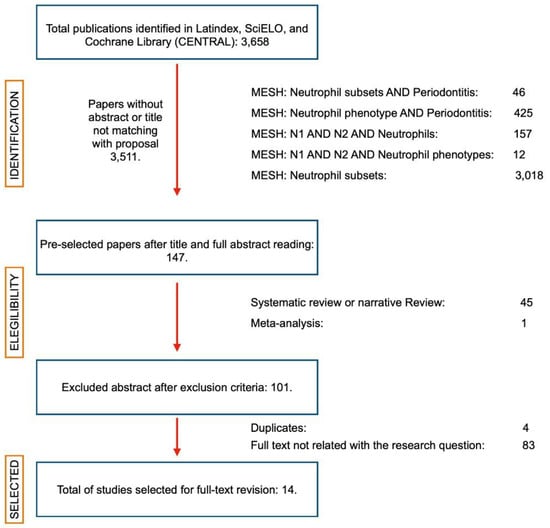
Figure 1.
PRISMA flow diagram. Summary of the article search. From a universe of 3658 articles, 147 passed the first review of abstract and title. Subsequently, systematic reviews with or without meta-analysis and narrative reviews were eliminated until 101 articles remained, which were reviewed in full-text format. Finally, 14 articles were selected to prepare this systematic review. (Created with http://biorender.com, accessed on March 2020).

Table 1.
Data extracted after individual analysis. Population, concept, and context individualization.
The selected articles correspond to studies in humans characterizing the presence of neutrophils in periodontal tissues based on their functions. Table 1 summarizes the main findings described. In addition, Table 2 shows the main findings for each study, indicating the results that answer our research question and gaps in the research.
Table 2.
Contribution of the selected studies to the outcome.
2.2. Inclusion of Sources of Evidence
In Table 2, the relevance of each selected source to the objectives of this scoping review is identified.
2.3. Review Findings
2.3.1. Phenotypic Characterization of Infiltrating Neutrophils in Periodontitis
During periodontitis, there is an increase in neutrophils in the periodontal tissues compared to the absence of periodontitis [20,26]. Neutrophils are characterized by CD10+, CD11b+, CD15+, CD16+, CD18+, CD55+, CD62L+, CD63+, CD64+, CD66+, CD138+, CD177+, and elastase+ [20,23,26]. In addition, they exhibit increased chemotaxis, phagocytosis, degranulation, production of cytokines, MPO and ROS, and NETosis [27,28,29]. In fact, in periodontitis-affected tissues, NET degradation is lower than in healthy subjects [26]. Additionally, 2.4% of oral neutrophils produce the receptor activator of the nuclear factor-κB ligand (RANKL) [30]. Additionally, in patients with refractory periodontitis, neutrophil ROS production is correlated with increased periodontal index (PI), bleeding on probing (BOP), pocket deep (PD), and clinical attachment level (CAL) [18]. This ROS production is independent of the chemotaxis capability and could be induced by local IFN-γ production [17,25].
When analyzing the neutrophil phenotypes described, there is no clear consensus. Some studies mention hyper-reactive or hyperactive phenotypes [16], naïve, pro-inflammatory, and para-inflammatory subsets [21,27]. These phenotypes can be described based on their capability to produce ROS [16,18], their euchromatin/heterochromatin ratio, and their higher or lesser phagocytic activity [21,27]. Interestingly, when patients are treated and a follow-up of 3, 6, or 12 months is carried out, it is observed that the CD16b+CD66+CD11b+CD62L− population decreases, and has a positive correlation with the BOP index [23]. Curiously, CD177+ cells predominate in gingival crevicular fluid (GFC) during periodontitis, while the CD177- population predominates under healthy conditions. Curiously, CD177+ cells have a higher apoptotic activity than CD177− cells [29].
Finally, when comparing circulating with oral neutrophils, the oral neutrophils present a unique transcriptome, characterized by increased IL-17, IFN-γ, RANKL, CXCL10, and elastase expression [19,30,31].
2.3.2. Para- and Pro-Inflammatory Neutrophils in Periodontitis
Periodontal tissues can be affected by different causes that induce inflammation. Periodontitis has been recognized as the main chronic inflammatory disease, but it is not the only one. There may be a form of periodontitis resulting from trauma, or there may be an adaptation phenomenon [32]. Thus, when periodontitis occurs due to dysbiosis of the microbiota, we speak of pro-inflammatory phenomena, and when periodontitis is due to an adaptive process, we speak of para-inflammatory phenomena [33,34]. This is noteworthy to mention, because the cause of the inflammation defines the type of inflammatory response, and the pro- and para-inflammatory phenotypes have been described in periodontal tissues [21]. In this context, a para-inflammatory and pro-inflammatory phenotype characterized by low granulation, light cytoplasm, and a large amount of euchromatin was detected [21]. Additionally, in healthy periodontal tissues, two different populations of oral neutrophils have been observed—para-inflammatory 1 and para-inflammatory 2—and both of them have a similar profile to naïve blood neutrophils and a lower state of activation [21].
2.3.3. Degranulation and NETosis
Considering degranulation and NETosis as an important part of neutrophil function, it is curious that only one study has evaluated NETosis [16]. The N1 phenotypes produce NET to induce the pro-inflammatory response, and aggNET enhances the N1 function. During periodontitis, the infiltration of neutrophils with high capability of triggering NETosis subsets is essential for determining the greater or lesser recruitment of phagocytes, which influences the increase or decrease in inflammatory signs. Additionally, it is not unusual to find a positive correlation between the presence of NET-producer neutrophils with the PI, and BOP index, or even CAL loss.
3. Methods
3.1. Review Question
Is there evidence characterizing the existence or the possible role of the N1 and N2 neutrophil phenotypes in periodontal tissues affected by periodontitis?
3.2. Participants, Concept, and Context
We include studies conducted in humans, in which the population is clearly identified. In particular, studies were selected where samples of adults between 20 and 80 years of age were used, regardless of the distribution by sex, socioeconomic level or ethnicity (participants). Additionally, the studies described both the periodontal diagnosis and the clinical criteria used to diagnose (concept). We selected the studies that analyzed or characterized the presence of neutrophils in the periodontal tissues of healthy people or those affected by periodontitis (concept), and the origin of each participant (context).
3.3. Source of Evidence
The present scoping review was carried out following the indications of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines. The protocol related to the data search, selection, extraction, and analysis was discussed and established by all of the authors in a dynamic drive and without further modifications. There were no language, year, or publication status restrictions for inclusion, and when eligible studies in languages other than English, Spanish, French or Portuguese were detected, a qualified translator was consulted. The selected articles were included in a drive for full text analysis, and the articles not selected were included in a table indicating the reasons for their non-inclusion or exclusion.
We included primary research studies, randomized or non-randomized clinical trials, and case–control and cohort studies. We excluded systematic reviews with or without meta-analysis from the analysis, as they are not a primary source of information. However, we left open the option to include any source of evidence directly associated with the research question and that met the eligibility criteria.
3.4. Search Strategy
An electronic search was performed in the Latindex, SciELO, Cochrane Library (CENTRAL) and Medline via Pubmed databases, from 30 January 2021 to 30 June 2022. Two reviewers (DS-E and JD-Z) performed the electronic database survey independently and in duplicate. The search strategy was performed using Mesh terms “neutrophil”, “bacteria”, “host–pathogens interactions”, “periodontitis”, “subsets”, and “phenotypes” in the Medline database and adapted for the other databases. The complete search strategy is provided in Supplementary file S1.
3.5. Source of Evidence Screening and Selection
Evidence screening and selection were performed independently and in duplicate, by assessing titles and abstracts to determine their inclusion. Furthermore, the selected full-texts were analyzed to determine whether they met the inclusion criteria. In case of disagreement, the article selection was discussed until a consensus was reached on whether the article could be included or excluded. If excluded, the motifs were recorded in the PRISMA flow chart (Figure 1). In cases where consensus was not reached, a third reviewer made the decision (SM-R).
3.6. Data Extraction
For each paper included, the following information was extracted: first author and year of publication, study objective, population (number, sex, age), concept (periodontal diagnosis criteria, interventions or phenomena of interest, samples, analysis methods, and results), and context (group definition and origin) (Table 1).
3.7. Outcome Measures
To assess the characterization of the neutrophils in the gingival tissues, the primary outcome was to determine the population of neutrophils in the periodontal tissues of patients with or without periodontal disease. As a secondary outcome, articles that isolated neutrophils from peripheral blood samples and evaluated phenotypes were selected. As a tertiary outcome, studies that included a complete phenotype or cytokine analysis in patients with inflammatory diseases were chosen.
4. Discussion
Periodontitis is one of the most prevalent pathologies in the oral cavity, which affects the supporting tissues of the teeth: periodontal ligament, root cement and alveolar bone; and its characteristic is the progressive destruction of these tissues with the possibility of a subsequent tooth loss [35,36]. Additionally, it is considered a non-communicable chronic disease, characterized by a high concentration of pro-inflammatory mediators in the affected periodontal tissue, which can spread to the peripheral circulation and induce a chronic low-grade inflammatory phenotype (CLIP) response [36]. The main cause of periodontitis is the dysbiosis of the subgingival microbiota, which begins with an alteration in the amount of bacteria [34,37].
Generally, periodontitis begins with a pro-inflammatory phenomenon, characterized by the presence of bacteria in the gingiva-dental sulcus and their recognition by Langerhans cells pattern recognition receptors (PRR) within the epithelium [38,39,40]. The Langerhans cells secrete IL-8, which will spread into the connective tissue creating a concentration gradient [38,39,40]. Furthermore, neutrophils can recognize the chemoattractant signal through their CD177 and migrate to the thickness of the epithelium [29,39,40]. Once neutrophils reach the site of inflammation and are activated, they facilitate microbial removal through several different processes, including phagocytosis, generation of myeloperoxidase (MPO) and ROS, release of granular content, cytokine production, and NETosis [24,41,42,43].
Different studies have evaluated the frequency of resident neutrophils in the periodontal tissues during health or periodontal disease, which vary from 0 to 100% of the leukocyte infiltrate [44,45]. Neutrophils are cells detected with high frequency in oral cavity, saliva, gingival sulcus, and inside pocket epithelium. Recently, 141 cluster of differentiation (CD) from neutrophils were described in healthy and inflammatory periodontal conditions [46]. Of them, CD11b, CD14, CD15, CD16, and CD62L have been used for its phenotypic characterization [20,23,27,46], CD11b/CD18, CD11c/CD18, CD21, and CD35 for phagocytosis evaluation [23,27,47,48], CD63 (primary granules), CD15 and CD66b (secondary granules), CD11b (tertiary granules), and CD13, CD14, CD18 and CD45 (quaternary granules) for degranulation analysis [15,20,23], and CD177 for maturation [49]. Additionally, two markers have been detected in some neutrophils: OLFM4 within granules, and CD177 in plasma membrane [7,50].
Other characteristics that have been observed in neutrophils are the degranulation capacity and death induced by NETosis. Neutrophils produce NET under different conditions, and it is made up of elastase, histones, MPO, and cathepsin-G, among other proteins [43]. In the presence of bacterial invasion, NETosis causes the release of IL-1β in the surrounding neutrophils and increase the production of chemokines such as CXCR2, CXCL8, CXCL1, CXCL3, and CXCL3, which produce a higher infiltrate of neutrophils favoring inflammation by degranulation [51]. Additionally, the neutrophils trigger the aggregation of NET, which initiates the anti-inflammatory process by trapping dead cells, bacteria, or phagocytes, facilitating the macrophages phagocytosis [52]. Interestingly, the NET production depends directly of the C-type lectin receptor Dectin-1, which is a size microorganism sensor [53]. The activation of Dectin-1 activates elastase, and elastase translocates to the nuclei and promotes histone degradation [43]. Once the concentration of neutrophils in the affected site of bacterial invasion exceeds a certain threshold, the NET begins to aggregate, building the aggNET [52]. AggNETs sequester DNA and proteins from neutrophil granules, and degrade pro-inflammatory cytokines through DNA-bound serine proteases. Indeed, as aggNET increases, more control exists over the pro-inflammatory phenomenon [43,52].
Several studies have determined the pro-inflammatory cytokines profile during periodontitis. Today, it has been widely recognized the role of IL-1β, IL-6, IL-8, IL-10, IL-12, IL-35, TNF-α, IFN-γ, and TGF-β in the balance of periodontal health or disease [34,54,55,56,57,58,59]. In this sense, an increase in granulocytes-colony stimulant factor (G-CSF), IL-6, IL-35, and TGF-β could trigger the differentiation towards N2 phenotypes, the increase of IL-12 and IFN-β, the differentiation towards N1 subsets, and the increase of IFN-γ and TNF-α could trigger the N2 to N1 subset conversion [5,60,61]. Recently, it has been proposed that under bacterial infection, the axis N1-M1 (pro-inflammatory macrophages) has a high phagocytic capacity, and the N2-M2 (modulatory macrophages) has a low ability for bacterial clearance [62,63]. Particularly, macrophages can differentiate toward a M1 phenotype that will promote the inflammatory and destructive response of damaged tissue. Otherwise, the M2 differentiate later and play a role in the resolution of inflammation and tissue regeneration [64]. After the first priming, neutrophils produce IL-8 and CXCL1 that allow adhesion, migration and chemoattraction, and represent a positive feedback loop [65,66]. Both subsets of neutrophils express similar levels of co-stimulatory molecules CD80, CD863, CD86, and HLA-DR, and cytokines, participating in antigen presentation [67]. Thus, N1 neutrophils could secrete IL-12, IFN-γ, and TNF-α, and polarize macrophages and dendritic cells (DCs) toward the pro-inflammatory subsets M1 or DC1, respectively. Conversely, against less pathogenic bacteria, neutrophils will produce IL-10 and TGF-β, which could induce M2- and DC2-modulatory subsets [34,57,59].
Aboodi and Johnstone described the high- and low-responder phenotypes based on ROS production, corresponding to the N1 and N2 subsets, respectively [18,24]. Additionally, the presence of neutrophils with CD15, CD16, CD62L, and CXCL10 markers corresponds to the N1 subset, and neutrophils with higher levels of CD11b and lower levels of CD62L correspond to the N2 subset [20,23,27]. In the presence of pathogenic mechanisms, neutrophils are primed by TLR2, TLR4, CD14, or even NOD1 [68]. Both subsets produce divergent amounts of CXCR4, which allows neutrophils to migrate to lymph nodes and present antigens to T lymphocytes. N1 neutrophils produce lower levels of CXCR4, while N2 neutrophils produce higher levels of CXCR4 than the N1 subset [69]. In this sense, the pro-inflammatory response triggered by N1 neutrophils could occur in situ after priming, and the modulatory response triggered by N2 probably occurs in the lymph-nodes. Nevertheless, plausible mechanisms for resolvign the gap in the knowledge regarding the capability of N2 to modulate the adaptive response in periodontitis is still unresolved.
Although we can identify that there is evidence in animals that neutrophils N1 and N2 have been described outside of a tumor environment, there are no studies that have characterized them during periodontal disease or in periodontal tissues [9]. A first approach for neutrophil subsets was related to their high or low density. High-density neutrophils (HDN) are Ly6GHIGHCD11bHIGH, mature, with segmented nuclei, cytotoxic, with a high capacity of migration, phagocytosis, and oxidative burst, non-suppressive, pro-inflammatory, and anti-tumorigenic [70,71,72]. Conversely, low-density neutrophils (LDN) are Ly6GHIGHCD11bLOW, mature, with segmented nuclei, non-cytotoxic, with a reduced migration, phagocytosis, and ROS production, suppressive, anti-inflammatory, and pro-tumorigenic. Additionally, HDN were named as belonging to a pro-inflammatory or anti-tumor phenotype, or N1, while LDN were denoted as belonging to an anti-inflammatory or pro-tumor phenotype, or N2 [70,72].
HDN subsets are characterized by Toll-like receptor (TLR)-2, TLR4, TLR5, TLR8, CD11bLOWCD49HIGHCD177HIGH expression and IL-12 production [73]. HDN subsets also produce higher amounts of TNF, CXCL10, and ROS, express higher levels of ICAM-1, CCL3, and CD95, and lower levels of CXC4, vascular endothelial growth factor (VEGF), and IL-8 [5,74,75,76]. Conversely, LDN neutrophils express TLR2, TLR4, TLR7, TLR9, and CD11bHIGHCD49LOWCD177LOW, and produce IL-10 [73]. LDN has a long lifespan and produces large amounts of arginase, which in turn inactivates T-cells, produce higher levels of CCL2, CXCR2, CXCR4, VEGF, IL-8, IL-10, and TGF-β1, and produce low levels of ICAM-1 and CCL3 [5,75,77]. LDN neutrophils also express MPO and produce ROS, but at a lower rate than HDN [76]. The greater or lesser extent of ROS production can be explained by its pro-inflammatory or homeostatic role. In fact, Aboodi [18] demonstrated the presence of two responder phenotypes, according to ROS production. HDN cells produce an oxidative burst, which contributes to local degranulation and bacterial killing, while LDN decreases ROS in order to produce genotoxicity in other immune cells [76]. When comparing published data, it is possible to identify the presence of different neutrophil phenotypes; however, whether they are N1 or N2 subsets can only be speculated (Figure 2). Curiously, in systemic inflammation, a third phenotype has been described, characterized by being CD62LOWCD11bHIGHCD11cHIGH, and by its strong capacity to inhibit T lymphocyte phenotypes by direct contact through Mac1 integrin and ROS production [53,78]. Additionally, two studies analyzed a population of neutrophils, detecting a third circulating phenotype, which could be the granulocytic myeloid-derived suppressor cells (gMSCD) [16,23,72].
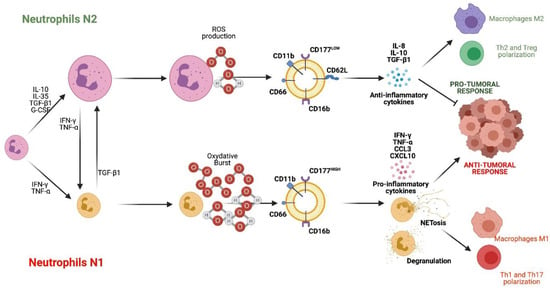
Figure 2.
Neutrophil subsets N1 and N2. N2 neutrophils are differentiated in the presence of IL-10, IL-35, TGF-β1, and G-CSF. N2 cells are characterized by producing ROS, IL-10 and TGF-β1, expressing CD11b, CD16b, CD62L, CD66 and CD177LOW, and having a pro-tumor and anti-inflammatory function. On the other hand, N1 neutrophils differentiate in the presence of IFN-γ and TNF-α. They are characterized by producing oxidative burst, IFN-γ, TNF-α, CCL3, CXCL4 and expressing CD16b, CD11b, CD66 and CD177HIGH, and fulfill an anti-tumor and pro-inflammatory role. References: [16,18,23,24,72,76]. (Created with http://biorender.com).
In general terms, there are no human studies demonstrating the presence of the N1 or N2 phenotypes of neutrophils in periodontal tissues. The closest approximations are related to the increase in pro-inflammatory functions in people with periodontitis, which in itself does not imply that there is an N1 phenotype. Thus, the absence of evidence does not allow us to demonstrate that these phenotypes exist in the periodontium, nor does it allow us to identify a possible role. Thus, in vitro and experimental studies are required to demonstrate the role of the two neutrophil phenotypes during periodontitis.
5. Future Research Directions
The evidence of neutrophil subsets in periodontitis is an unexplored field. Even though several studies have determined different neutrophil roles, the role of the N1 or N2 subsets in periodontal health or disease still cannot be concluded. It is necessary to determine the pathways by which oral bacteria may be able to induce N1 or N2 response in in vitro or experimental models. Additionally, it is necessary to determine the frequency of detection of these neutrophil phenotypes in periodontal health or disease.
6. Conclusions
The presence of neutrophils increases in tissues affected by periodontitis compared to healthy tissues. These neutrophils have a pro-inflammatory phenotype characterized by increased phagocytosis, degranulation, production of pro-inflammatory cytokines, and NETosis. However, neither study defines detected neutrophils as being either N1 or N2.
7. Implication of the Findings for Research
This scoping review demonstrates that there is a gap in knowledge. In this context, new research hypotheses can be developed that must be resolved in the future. Additionally, Figure 3 represents a hypothesis regarding the role of both subsets during periodontitis, which must be proven in the future.
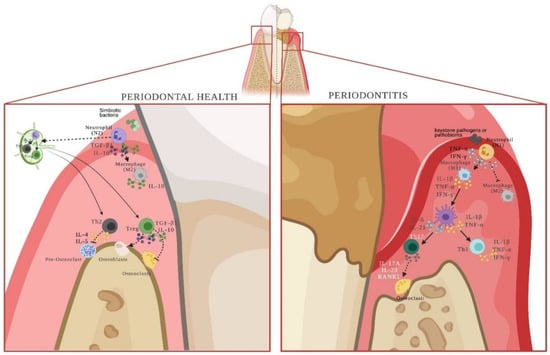
Figure 3.
N1 and N2 neutrophils and their role during periodontal health or disease. Neutrophils, by recognizing symbiotic bacteria, have the ability to differentiate into the N2 phenotype. Cytokines produced by N2 cells will polarize macrophages towards M2-anti-inflammatory subsets. In addition, they will migrate to the regional lymph-node and present the antigen to naïve lymphocytes to allow their differentiation into the Th2 and Treg phenotypes, which will naturally maintain tissue homeostasis. Conversely, in the presence of keystone bacteria or pathobionts, neutrophils polarize towards the N1 phenotype, which secretes cytokines that allow the differentiation of pro-inflammatory M1 macrophages and pro-inflammatory dendritic cells. Together, they present antigens to T lymphocytes at the affected periodontal site, differentiating them towards Th1 and Th17 effector phenotypes. Thus, the response initiated by N2 neutrophils is a regional response, and the one triggered by N1 neutrophils is localized. References: [5,34,57,59,60,61,62,63,64,65,66,69]. (Created with http://biorender.com).
8. Implication of the Findings for Practice
Understanding the role of neutrophils as the first line of immune response will make it possible to design therapeutic alternatives for the treatment of pro-inflammatory diseases in order to avoid N1 polarization or induce N2 polarization. Specifically, periodontitis is caused by keystone pathogens that trigger a pro-inflammatory response. In this context, determining what type of neutrophil response each bacterium induces would be important for assessing the microbiological or immunological susceptibility of each individual.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232012068/s1, Supplementary file S1: 1Search strategies
Author Contributions
Conceptualization, V.M.M.-A., J.D.-Z., L.D.S.-E. and S.M.-R.; methodology, V.M.M.-A., J.D.-Z., L.D.S.-E. and S.M.-R.; software, V.M.M.-A., J.D.-Z. and L.D.S.-E.; validation, L.D.S.-E., S.M.-R., R.V., B.A.C.-Á., V.M.M.-A. and J.D.-Z.; formal analysis, L.D.S.-E., S.M.-R., R.V., B.A.C.-Á., V.M.M.-A. and J.D.-Z.; investigation, L.D.S.-E., S.M.-R. and J.D.-Z.; resources, S.M.-R., B.A.C.-Á., V.M.M.-A. and J.D.-Z.; data curation, L.D.S.-E., V.M.M.-A. and J.D.-Z.; writing—original draft preparation, L.D.S.-E., S.M.-R., R.V., B.A.C.-Á., V.M.M.-A. and J.D.-Z.; writing—review and editing, L.D.S.-E., S.M.-R., R.V., B.A.C.-Á., V.M.M.-A. and J.D.-Z.; visualization, L.D.S.-E., S.M.-R., R.V., B.A.C.-Á., V.M.M.-A. and J.D.-Z.; supervision, J.D.-Z.; project administration, V.M.M.-A. and J.D.-Z.; funding acquisition, S.M.-R., V.M.M.-A., J.D.-Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by RDP-IADR 2019-2023, Chilean government Grant FONDECYT Nº1220999.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The study did not report any data.
Acknowledgments
All the author have consented to the acknowledgment.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Ost, M.; Singh, A.; Peschel, A.; Mehling, R.; Rieber, N.; Hartl, D. Myeloid-Derived suppresor cells in bacterial infections. Front. Cell. Infect. Microbiol. 2016, 6, 37. [Google Scholar] [CrossRef]
- Pillay, J.; den Braber, I.; BVrisekoop, N.; Kwast, L.M.; de Boer, R.J.; Borghans, J.A.M.; Tesselaar, K.; Koenderman, L. In vivo labeling with 2H2O reveals a human lifespan of 5.4 days. Blood 2010, 116, 625–627. [Google Scholar] [CrossRef]
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef]
- Imtiyaz, H.Z.; Simon, M.C. Hypoxia-inducible factors as essential regulators of inflammation. Curr. Top. Microbiol. Immunol. 2010, 345, 20. [Google Scholar]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-beta: "N1" versus "N2" TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar]
- Fridlender, Z.G.; Albelda, S.M. Tumor-associated neutrophils: Friend or foe? Carcinogenesis 2012, 33, 949–955. [Google Scholar] [CrossRef]
- Goldschmeding, R.; van Dalen, C.M.; Faber, N.; Calafat, J.; Huizinga, T.W.; van der Schoot, C.E.; Clement, L.T.; von dem Borne, A.E. Further characterization of the NB 1 antigen as a variably expressed 56–62 kD GPI- linked glycoprotein of plasma membranes and specific granules of neu- trophils. Br. J. Haematol. 1992, 81, 336–345. [Google Scholar] [CrossRef]
- Masucci, M.T.; Minopoli, M.; Carriero, V. Tumor Associated Neutrophils. Their role in tumorigenesis, metastasis, prognosis and therapy. Front. Oncol. 2019, 9, 1146. [Google Scholar] [CrossRef]
- Ma, Y.; Yabluchanskiy, A.; Iyer, R.P.; Cannon, P.L.; Flynn, E.R.; Jung, M.; Henry, J.; Cates, C.A.; Deleon-Pennell, K.Y.; Lindsey, M.L. Temporal neutrophil polarization following myocardial infarction. Cardiovasc. Res. 2016, 110, 51–60. [Google Scholar] [CrossRef]
- Perez de Puig, I.; Miro-Mur, F.; Ferrer-Ferrer, M.; Gelpi, E.; Pedragosa, J.; Justicia, C.; Urra, X.; Chamorro, A.; Planas, A.M. Neutrophil recruitment to the brain in mouse and human ischemic stroke. Acta Neuropathol. 2015, 129, 239–257. [Google Scholar] [CrossRef]
- Cai, W.; Liu, S.; Hu, M.; Huang, F.; Zhu, Q.; Qiu, W.Q.; Hu, X.; Colello, J.; Zheng, S.G.; Lu, Z. Functional dynamics of neutrophils after ischemic stroke. Transl. Stroke Res. 2020, 11, 108–121. [Google Scholar] [CrossRef]
- Cuartero, M.I.; Ballesteros, I.; Moraga, A.; Nombela, F.; Vivancos, J.; Hamilton, J.A.; Corbí, A.L.; Lizacoain, I.; Moro, M.A. N2 neutrophils, novel players in brain inflammation after stroke: Modulation by the PPARgamma agonist rosiglitazone. Stroke 2013, 44, 3498–3508. [Google Scholar] [CrossRef]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Theilgaard-Mönch, K.; Knudsen, S.; Follin, P.; Borregaard, N. The transcriptional activation program of human neutrophils in skin lesions supports their important role in wound healing. J. Immunol. 2004, 172, 7684–7693. [Google Scholar] [CrossRef]
- Naegelen, I.; Beaume, N.; Plançon, S.; Schenten, V.; Tschirhart, E.J.; Bréchard, S. Regulation of neutrophil degranulation and cytokine secretion: A novel model approach based on linar fitting. J. Immunol. Res. 2015, 2015, 817038. [Google Scholar] [CrossRef]
- Matthews, J.B.; Wright, H.J.; Roberts, A.; Cooper, P.R.; Chapple, I. Hyperactivity and reactivity of peripheral blood neutrophils in chronic periodontitis. Clin. Exp. Immunol. 2006, 147, 255–264. [Google Scholar] [CrossRef]
- Yagi, M.; Kantarçi, A.; Iwata, T.; Omori, K.; Ayilavarapu, S.; Ito, K.; Hasturk, H.; van Dyke, T.E. PDK1 regulates chemotaxis in human neutrophils. J. Dent. Res. 2009, 88, 1119–1124. [Google Scholar] [CrossRef]
- Aboodi, G.M.; Goldberg, M.B.; Glogauer, M. Refractory periodontitis population characterized by a hyperactive oral neutrophil phenotype. J. Periodontol. 2011, 82, 726–733. [Google Scholar] [CrossRef]
- Lakschevitz, F.S.; Aboodi, G.M.; Glogauer, M. Oral neutrophils display a site-specific phenotype characterized by expression of T-cell receptors. J. Periodontol. 2013, 84, 1493–1503. [Google Scholar] [CrossRef]
- Dutzan, N.; Konkel, J.E.; Greenwell-Wild, T.; Moutsopoulos, N.M. Characterization of the human immune cell network at the gingival barrier. Mucosal Immunol. 2016, 9, 1163–1172. [Google Scholar]
- Borenstein, A.; Fine, N.; Hassanpour, S.; Sun, C.; Oveisi, M.; Tenenbaum, H.C.; Glogauer, M. Morphological characterization of para- and proinflammatory neutrophil phenotypes using transmission electron microscopy. J. Periodontal Res. 2018, 53, 972–982. [Google Scholar] [CrossRef]
- Papantonopoulos, G.; Delatola, C.; Takahashi, K.; Laine, M.L.; Loos, B.G. Hidden noise in immunologic parameters might explain rapid progression in aerly-onset peeriodontitis. PLoS ONE 2019, 14, e0224615. [Google Scholar] [CrossRef]
- Medara, N.; Lenzo, J.C.; Walsh, K.A.; Reynolds, E.C.; O’Brien-Simpson, N.M.; Darby, I.B. Peripheral neutrophil phenotypes during management of periodontitis. J. Periodontal Res. 2020, 56, 58–68. [Google Scholar]
- Johnstone, A.M.; Koh, A.; Goldberg, M.B.; Glogauer, M. A hyperactive neutrophil phenotype in patients with refractory periodontitis. J. Periodontol. 2007, 78, 1788–1794. [Google Scholar] [CrossRef]
- Wright, H.J.; Matthews, J.B.; Chapple, I.; Ling-Mountford, N.; Cooper, P. Periodontitis associates with type 1 IFN signature in peripheral blood neutrophils. J. Immunol. 2008, 181, 5775–5784. [Google Scholar] [CrossRef]
- Thorbert-Mros, S.; Larsson, L.; Berglundh, T. Cellular composition of long-standing gingivitis and periodontitis lesions. J. Periodontal Res. 2014, 50, 535–543. [Google Scholar] [CrossRef]
- Fine, N.; Hassanpour, S.; Borenstein, A.; Sima, C.; Oveisi, M.; Scholey, J.; Cherney, D.; Glogauer, M. Distinct oral neutrophil subsets define health and periodontal disease states. J. Dent. Res. 2016, 95, 931–938. [Google Scholar] [CrossRef]
- White, P.; Sakellari, D.; Roberts, H.; Risafi, I.; Ling, M.; Cooper, P.; Milward, M.; Chapple, I. Peripheral blood neutrophil extracellular trap production and degradation in chronic periodontitis. J. Clin. Periodontol. 2016, 43, 1041–1049. [Google Scholar] [CrossRef]
- Rudin, A.D.; Amirbeagi, F.; Davidsson, L.; Khamzeh, A.; Thorbert-Mros, S.; Thulin, P.; Welin, A.; Björkman, L.; Christenson, K.; Bylund, J. The neutrophil subset defined by CD177 expression is preferentially recruited to gingival crevicular fluid in periodontitis. J. Leukoc. Biol. 2020, 109, 349–362. [Google Scholar]
- Moonen, C.G.J.; de Vries, T.J.; Rijkschroeff, P.; Poubelle, P.E.; Nicu, E.A.; Loos, B.G. The possible role of neutrophils in the induction of osteoclastogenesis. J. Immunol. Res. 2019, 2019, 8672604. [Google Scholar]
- Moutsopoulos, N.M.; Konkel, J.E.; Sarmadi, M.; Eskan, M.A.; Wild, T.; Dutzan, N.; Abusleme, L.; Zenobia, C.; Hosur, K.B.; Abe, T.; et al. Defective neutrophil recruitment in leukocyte adhesion deficiency type I disease causes IL-17-driven inflammatory bone loss. Sci. Transl. Med. 2014, 6, 229ra40. [Google Scholar] [CrossRef]
- Caton, J.G.; Armitage, G.; Berglundh, T.; Chapple, I.; Jepsen, S.; Kornman, K.S.; Mealey, B.L.; Papapanou, P.N.; Sanz, M.; Tonetti, M.S. A new classification scheme for periodontal and peri-implant diseases and conditions - Introduction and key changes from the 1999 classification. J. Clin. Periodontol. 2018, 45, S1–S8. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef]
- Armitage, G. Development of a Classification System for Periodontal Diseases and Conditions. Ann. Periodontol. 1999, 4, 1–110. [Google Scholar] [CrossRef]
- Papapanou, P.N.; Sanz, M.; Buduneli, N.; Dietrich, T.; Feres, M.; Fine, D.H.; Flemmig, T.F.; Garcia, R.; Giannobile, W.V.; Graziani, F.; et al. Periodontitis: Consensus report of workgroup 2 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Clin. Periodontol. 2018, 45, S162–S170. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Seguier, S.; Godeau, G.; Leborgne, M.; Pivert, G.; Brousse, N. Quantitative morphological analysis of Langerhans cells in healthy and diseased human gingiva. Arch. Oral Biol. 2000, 45, 1073–1081. [Google Scholar]
- Kinane, D.F. Causation and pathogenesis of periodontal disease. Periodontology 2001, 25, 8–20. [Google Scholar] [CrossRef]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Rev. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.; Knopf, J.; Maueröder, C.; Kienhöfer, D.; Leppkes, M.; Herrmann, M. Neutrophils and neutrophil extracellular traps orchestrate initiation and resolution of inflammation. Clin. Exp. Rheumatol. 2016, 34, 6–8. [Google Scholar]
- Zhou, G.; Yu, L.; Fang, L.; Yang, W.; Yu, T.; Miao, Y.; Chen, M.; Wu, K.; Chen, F.; Cong, Y.; et al. CD177+ neutrophils as functionally activated neutrophils negatively regulate IBD. Gut 2017, 67, 1052–1063. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A cell with many roles in inflammation or several cell types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Matsuo, K.; Lin, A.; Procter, J.L.; Clement, L.T.; Stroncek, D. Variations in the expression of granulocyte antigen NB1. Transfusion 2000, 40, 654–662. [Google Scholar] [CrossRef]
- Taniguchi, K.; Kobayashi, M.; Harada, H.; Hiraoka, A.; Tanihiro, M.; Takata, N.; Kimura, A. Human neutrophil antigen-2a expression on neutrophils from healthy adults in western Japan. Transfusion 2002, 42, 651–657. [Google Scholar] [CrossRef]
- Lakschevitz, F.S.; Hassanpour, S.; Rubin, A.; Fine, N.; Sun, C.; Glogauer, M. Identification of neutrohpil surface marker changes in health and inflammation using high-thourhput screening flow cytometry. Exp. Cell Res. 2016, 342, 200–209. [Google Scholar] [CrossRef]
- Holers, V.M. Complement and its receptors: New insights into human disease. Annu. Rev. Immunol. 2014, 32, 433–459. [Google Scholar] [CrossRef]
- van Kessel, K.P.M.; Bestebroer, J.; van Strijp, J.A.G. Neutrophil-mediated phagocytosis of Staphylococcus aureus. Front. Immunol. 2014, 5, 467. [Google Scholar] [CrossRef]
- Galli, S.J.; Borregaard, N.; Wynn, T.A. Phenotypic and functional plasticity of cells of innate immunity: Macrophages, mast cells and neutrophils. Nat. Immunol. 2012, 12, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, S.N.; Bohr, C.T.; Rorvig, S.; Glenthoj, A.; Mora-Jensen, H.; Cramer, E.P.; Jacobsen, L.C.; Larsen, M.T.; Cowland, J.B.; Tanassi, T.; et al. Olfactomedin 4 defines a subset of human neutrophils. J. Leukoc. Biol. 2012, 91, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Terkeltaub, R.; Baird, S.; Sears, P.; Santiago, R.; Boisvert, W. The murine homolog if the interleukin-8 receptor CXCR2 is essential for the occurrence of neutrophilic inflammation in the air pouch model of acute urate crystal-induced gouty synovitis. Arthritis Rheum. 1998, 41, 900–909. [Google Scholar] [CrossRef]
- Schauer, C.; Janko, C.; Munoz, L.E.; Zhao, Y.K.D.; Frey, B.; Lell, M.; Manger, B.; Rech, J.; Naschberger, E.; Holmdahl, R.; et al. Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines. Nat. Med. 2014, 20, 511–517. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, G.; Manwani, D.; Morthe, A.; Xu, C.; Faith, J.J.; Burk, R.D.; Kunisaki, Y.; Jang, J.E.; Scheiermann, C.; et al. Neutrophil ageing is regulated by the microbiome. Nature 2015, 525, 528–532. [Google Scholar] [CrossRef]
- Vernal, R.; Dutzan, N.; Chaparro, A.; Puente, J.; Valenzuela, M.; Gamonal, J. Levels of interleukin-17 in gingival crevicular fluid and in supernatants of cellular cultures of gingival tissue from patients with chronic periodontitis. J. Clin. Periodontol. 2005, 32, 383–389. [Google Scholar] [CrossRef]
- Vernal, R.; Dutzan, N.; Hernández, M.; Chandía, S.; Puente, J.; León, R.; García, L.; Del Valle, I.; Silva, A.; Gamonal, J. High expression levels of receptor activator of nuclear factor-kappa B ligand associated with human chronic periodontitis are mainly secreted by CD4+ T lymphocytes. J. Periodontol. 2006, 77, 1772–1780. [Google Scholar] [CrossRef]
- Dutzan, N.; Vernal, R.; Hernández, M.; Dezerega, A.; Rivera, O.; Silva, N.; Aguillon, J.C.; Puente, J.; Pozo, P.; Gamonal, J. Levels of interferon-gamma and transcription factor T-bet in progressive periodontal lesions in patients with chronic periodontitis. J. Periodontol. 2009, 80, 290–296. [Google Scholar] [CrossRef]
- Hernández, M.; Dutzan, N.; García-Sesnich, J.; Abusleme, L.; Dezerega, A.; Silva, N.; González, F.E.; Vernal, R.; Sorsa, T.; Gamonal, J. Host-pathogen interactions in progressive chronic periodontitis. J. Dent. Res. 2011, 90, 1164–1170. [Google Scholar] [PubMed]
- Hajishengallis, G. Immuno-microbial pathogenesis of periodontitis: Keystones, pathobionts, and the host response. Trends Immunol. 2014, 35, 3–11. [Google Scholar] [CrossRef]
- Silva, N.; Abusleme, L.; Bravo, D.; Dutzan, N.; García-Sesnich, J.; Vernal, R.; Hernández, M.; Gamonal, J. Host response mechanisms in periodontal diseases. J. Appl. Oral Sci. 2015, 23, 329–355. [Google Scholar] [CrossRef] [PubMed]
- Jablonska, J.; Lechner, S.; Westphal, K.; Lienenklaus, S.; Weiss, S. Neutrophils responsive to endogenous IFN-B regulate tumor angiogenesis and growth in a mouse tumor model. J. Clin. Investig. 2010, 120, 1151–1164. [Google Scholar]
- Zou, J.M.; Qin, J.; Li, Y.C.; Wang, Y.; Li, D.; Shu, Y.; Luo, C.; Wang, S.S.; Chi, G.; Guo, F.; et al. IL-35 induces N2 phenotype of neutrophils to promote tumor growth. Oncotarget 2017, 8, 33501–33514. [Google Scholar] [CrossRef]
- Shigematsu, K.; Asai, A.; Kobayashi, M.; Herndon, D.N.; Suzuki, F. Enterococcus faecalis translocation in mice with severe burn injury: A pathogenic role of CCL2 and alternatively activated macrophages (M2aMo abd M2cMc). J. Leukoc. Biol. 2009, 86, 999–1005. [Google Scholar] [CrossRef]
- Neely, C.J.; Kartchner, L.B.; Mendoza, A.E.; Linz, B.M.; Frelinger, J.A.; Wolfgang, M.C.; Maile, R.; Cairns, B.A. Flagellin treatment prevents increased susceptibility to systemic bacterial infection after injury by inhibiting anti-inflammatory IL-10+IL-12- neutrophil polarization. PLoS ONE 2014, 9, e85623–e85633. [Google Scholar] [CrossRef]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M1/M2 macrophages and the Th1/Th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef]
- Gasperini, S.; Calzetti, F.; Russo, M.P.; de Gironcoli, M.; Cassatella, M.A. Regulation of GRO alpha production in human granulocytes. J. Inflamm. 1995, 45, 143–151. [Google Scholar]
- van Gisbergen, K.P.J.M.; Geijtenbeek, T.B.H.; van Kooyk, Y. Close encounters of neutrofils and DCs. Trends Immunol. 2005, 26, 626–631. [Google Scholar] [CrossRef]
- Shaul, M.E.; Levy, L.; Sun, J.; Mishalian, I.; Singhal, S.; Kapoor, V.; Horng, W.; Fridlender, G.; Albelda, S.M.; Fridlender, Z.G. Tumor-associated neutrophils display a distinct N1 profile following TGFB modulation: A transcriptomics analysis of pro- vs. antitumor TANs. Oncoimmunology 2016, 5, e1232221. [Google Scholar] [CrossRef]
- Hergott, C.B.; Roche, A.M.; Tamashiro, E.; Clarke, T.B.; Bailey, A.G.; Laughlin, A.; Bushman, F.D.; Weiser, J.N. Peptidoglycan from the gut microbiota governs the lifespan of circulatinf phagocytes at homeostasis. Blood 2016, 127, 2460–2471. [Google Scholar] [CrossRef]
- Gorlino, C.V.; Ranocchia, R.P.; Harman, M.F.; García, I.A.; Crespo, M.I.; Morón, G.; Maletto, B.A.; Pistoresi-Palencia, M.C. Neutrophils exhibit differential requirements for homing molecules in their lymphatic and blood trafficking into draining lymph nodes. J. Immunol. 2014, 193, 1966–1974. [Google Scholar]
- Granot, Z.; Henke, E.; Comen, E.A.; King, T.A.; Norton, L.; Benezra, R. Tumor entrained neutrophils inhibit seeding in the premetastasic lung. Cancer Cell 2011, 20, 300–314. [Google Scholar] [CrossRef]
- Rayner, B.S.; Love, D.T.; Hawkins, C.L. Comparative reactivity of myeloperoxidase-derived oxidants with mammalian cells. Free. Radic. Biol. Med. 2014, 71, 240–255. [Google Scholar] [CrossRef]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef]
- Tsuda, Y.; Takahashi, H.; Kobayashi, M.; Hanafusa, T.; Herndon, D.N.; Suzuki, F. Three different neutrophil subsets exhibited in mice with different susceptibilities to infection by methicillin-resistant Staphylococcus aureus. Immunity 2004, 21, 215–226. [Google Scholar] [CrossRef]
- Strieter, R.M.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Polverini, P.J. Interferon gamma-inducible protein 10 (IP-10), a member of the C-X-C- chemokine family, is an inhibitor of angiogenesis. Biochem. Biophys. Res. Commun. 1995, 210, 51–57. [Google Scholar]
- Zhang, X.; Shi, H.; Jiang, P.; Qian, H.; Xu, W. Tumor-derived exosomes induce N2 polarization of neutrophils to promote gastric cancer cell migration. Mol. Cancer 2018, 17, 146. [Google Scholar]
- Ohms, M.; Möller, S.; Laskay, T. An attempt to polarize human neutrophils toward N1 and N2 phenotypes in vitro. Front. Immunol. 2020, 11, 532. [Google Scholar] [CrossRef]
- Scaverlandi, M.V.; Peinetti, N.; Leimgruber, C.; Cuello Rubio, M.M.; Nicola, J.P.; Menezes, G.B.; Maldonado, C.A.; Quintar, A.A. Inefficient N2-Like neutrophils are promoted by androgens during infection. Front. Immunol. 2018, 9, 1980. [Google Scholar]
- Pillay, J.; Tak, T.; Kamp, V.M.; Koenderman, L. Immune suppression by neutrophils and granulocytic myeloid-derived suppressor cells: Similarities and differences. Cell. Mol. Life Sci. 2013, 70, 3813–3827. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).