
IFT20: An Eclectic Regulator of Cellular Processes beyond Intraflagellar Transport

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
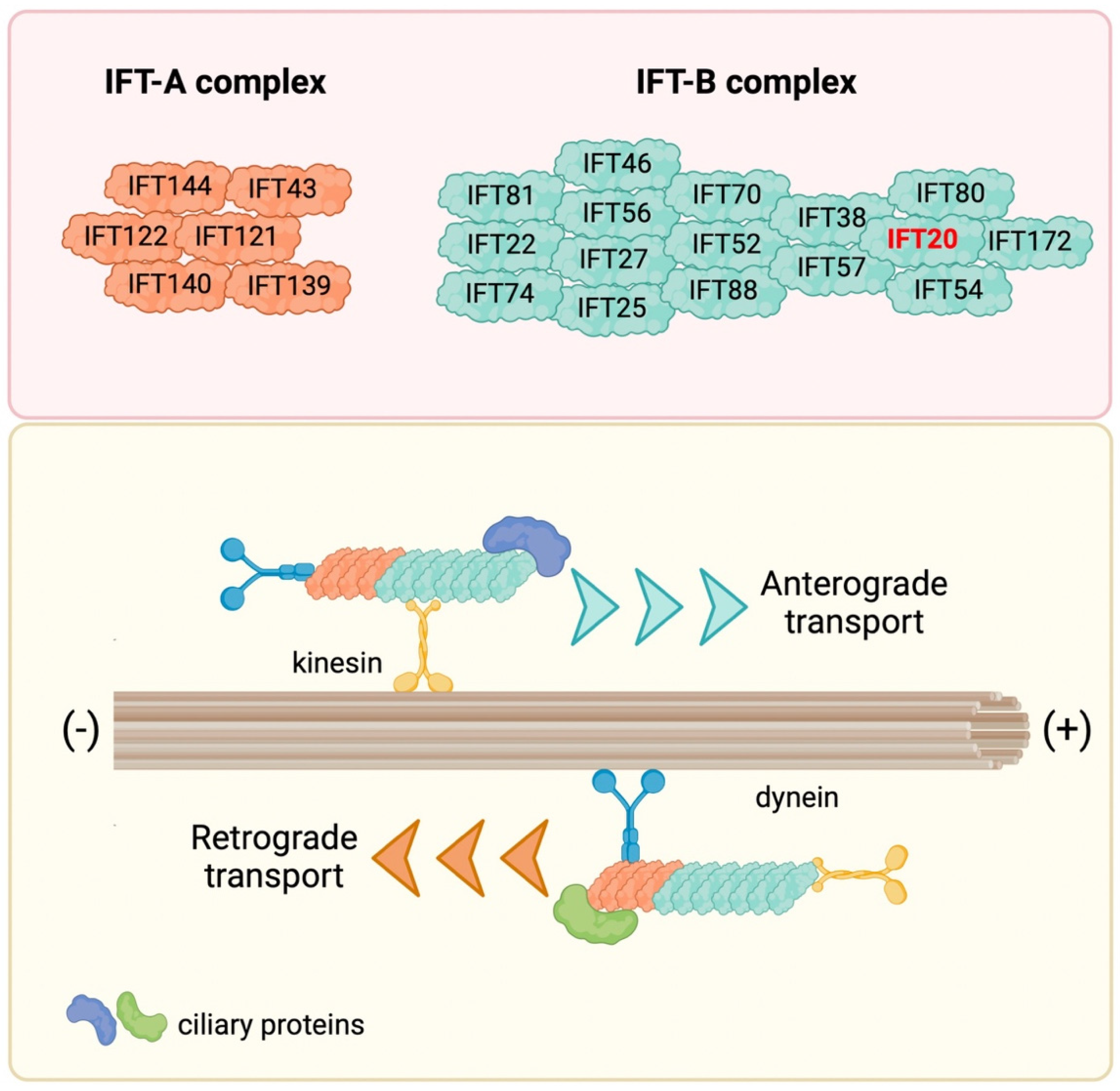
2. Molecular Dissection of IFT20: Structure and Localization
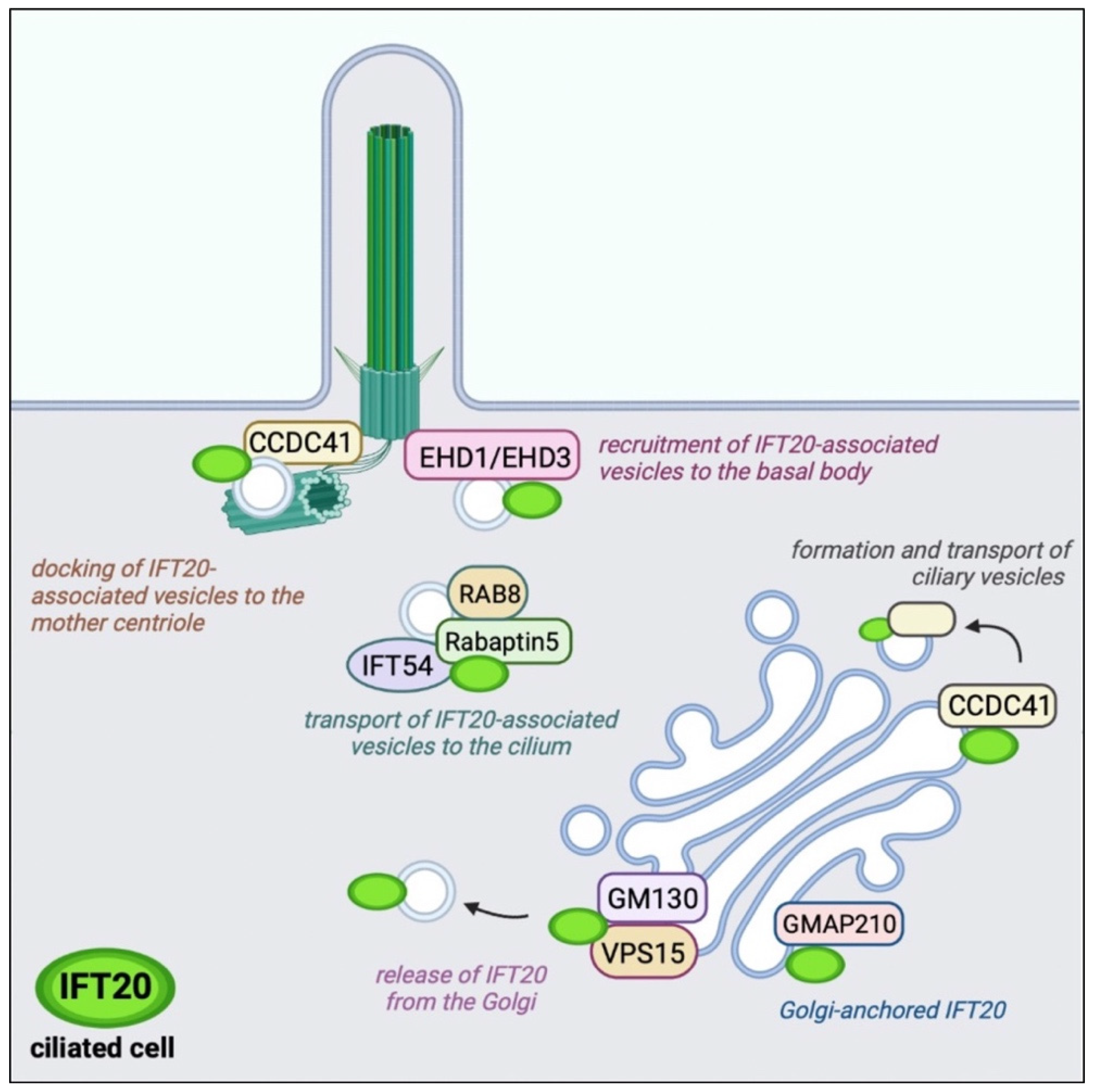
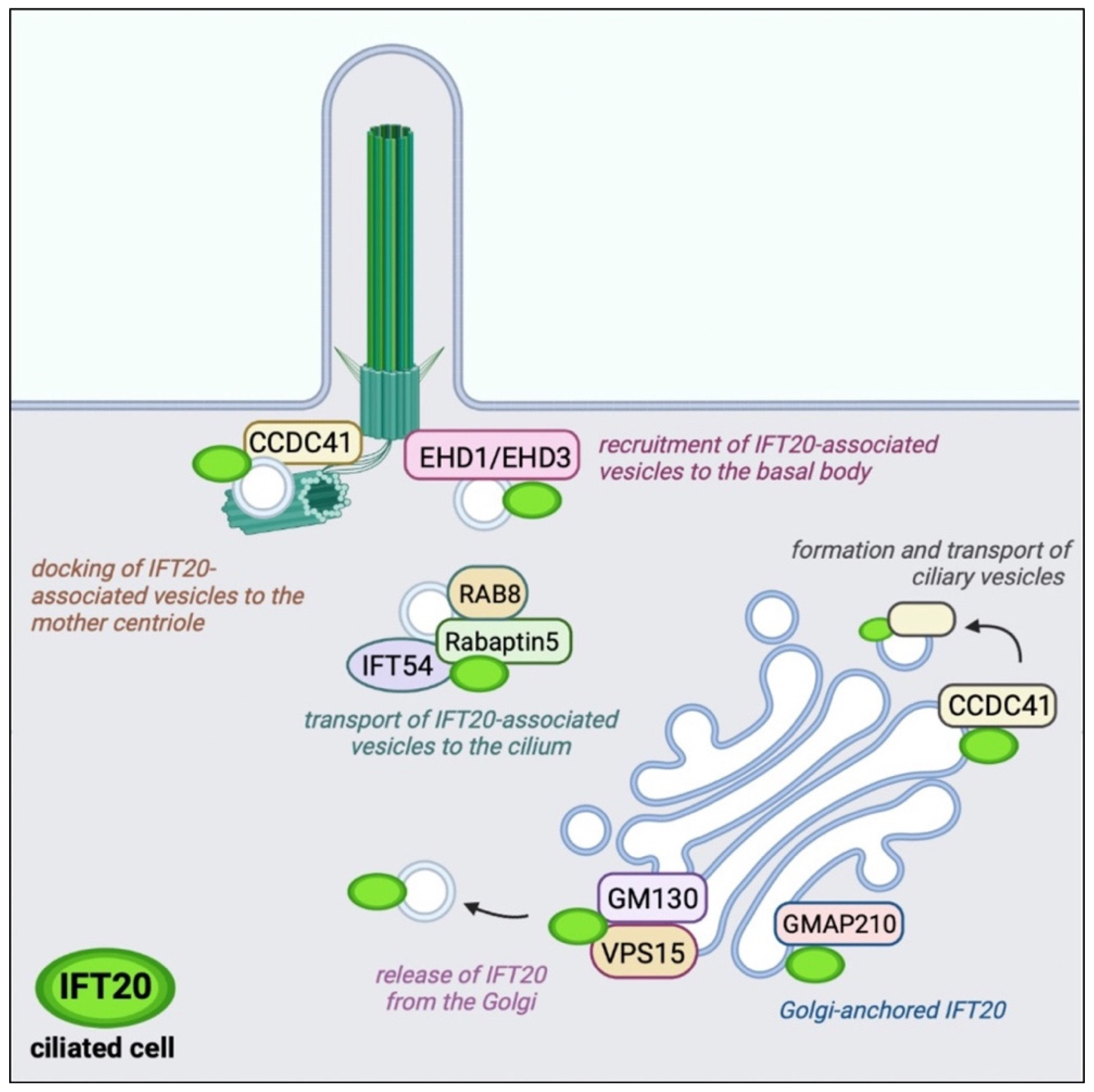
3. Ciliary Trafficking of IFT20: From the Golgi Apparatus to the PC Tip
3.1. Cilium Assembly and Maintenance
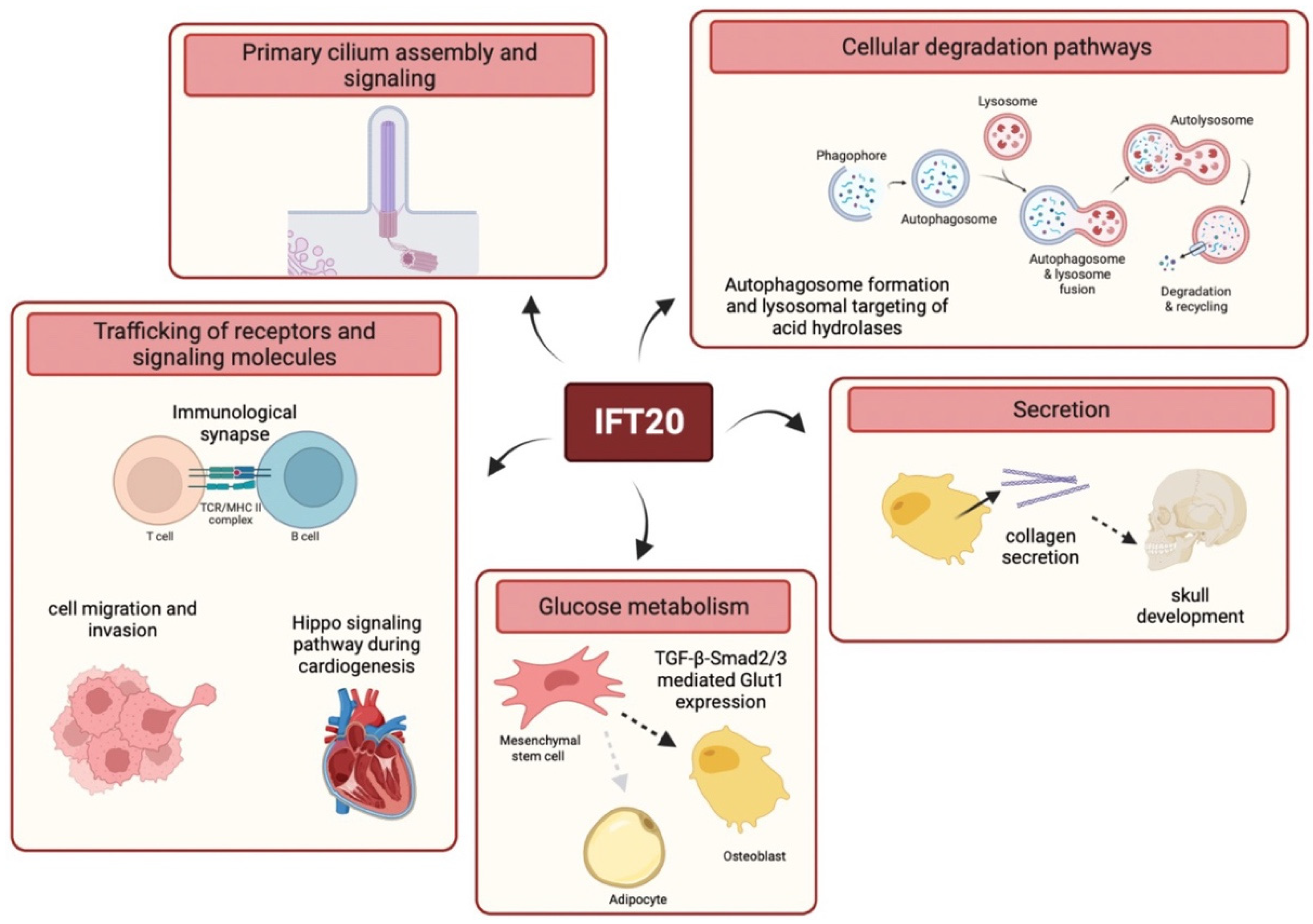
3.2. Trafficking of Receptors and Signaling Molecules
4. Cilia-Unrelated Vesicular Trafficking
4.1. Trafficking of Receptors and Signaling Molecules
4.2. Cellular Degradation Pathways
4.3. Secretion
4.4. Glucose Metabolism
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kozminski, K.G.; Johnson, K.A.; Forscher, P.; Rosenbaum, J.L. A Motility in the Eukaryotic Flagellum Unrelated to Flagellar Beating. Proc. Natl. Acad. Sci. USA 1993, 90, 5519–5523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilley, A.E.; Walters, M.S.; Shaykhiev, R.; Crystal, R.G. Cilia Dysfunction in Lung Disease. Annu. Rev. Physiol. 2015, 77, 379–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satir, P.; Pedersen, L.B.; Christensen, S.T. The Primary Cilium at a Glance. J. Cell Sci. 2010, 123, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Bornens, M. The Centrosome in Cells and Organisms. Science 2012, 335, 422–426. [Google Scholar] [CrossRef]
- Nigg, E.A.; Raff, J.W. Centrioles, Centrosomes, and Cilia in Health and Disease. Cell 2009, 139, 663–678. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.A.; Rosenbaum, J.L. Polarity of Flagellar Assembly in Chlamydomonas. J. Cell Biol. 1992, 119, 1605–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, W.F.; Rosenbaum, J.L. Intraflagellar Transport Balances Continuous Turnover of Outer Doublet Microtubules: Implications for Flagellar Length Control. J. Cell Biol. 2001, 155, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Pazour, G.J.; Agrin, N.; Leszyk, J.; Witman, G.B. Proteomic Analysis of a Eukaryotic Cilium. J. Cell Biol. 2005, 170, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, J.L.; Witman, G.B. Intraflagellar Transport. Nat. Rev. Mol. Cell Biol. 2002, 3, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Scholey, J.M. Intraflagellar Transport. Annu. Rev. Cell Dev. Biol. 2003, 19, 423–443. [Google Scholar] [CrossRef] [PubMed]
- Pigino, G.; Geimer, S.; Lanzavecchia, S.; Paccagnini, E.; Cantele, F.; Diener, D.R.; Rosenbaum, J.L.; Lupetti, P. Electron-Tomographic Analysis of Intraflagellar Transport Particle Trains in Situ. J. Cell Biol. 2009, 187, 135–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piperno, G.; Mead, K. Transport of a Novel Complex in the Cytoplasmic Matrix of Chlamydomonas Flagella. Proc. Natl. Acad. Sci. USA 1997, 94, 4457–4462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, D.G.; Diener, D.R.; Himelblau, A.L.; Beech, P.L.; Fuster, J.C.; Rosenbaum, J.L. Chlamydomonas Kinesin-II-Dependent Intraflagellar Transport (IFT): IFT Particles Contain Proteins Required for Ciliary Assembly in Caenorhabditis Elegans Sensory Neurons. J. Cell Biol. 1998, 141, 993–1008. [Google Scholar] [CrossRef]
- Hou, Y.; Qin, H.; Follit, J.A.; Pazour, G.J.; Rosenbaum, J.L.; Witman, G.B. Functional Analysis of an Individual IFT Protein: IFT46 Is Required for Transport of Outer Dynein Arms into Flagella. J. Cell Biol. 2007, 176, 653–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, J.M.; Harris, J.A.; Hyman, S.; Kner, P.; Lechtreck, K.F. Tubulin Transport by IFT Is Upregulated during Ciliary Growth by a Cilium-Autonomous Mechanism. J. Cell Biol. 2015, 208, 223–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wren, K.N.; Craft, J.M.; Tritschler, D.; Schauer, A.; Patel, D.K.; Smith, E.F.; Porter, M.E.; Kner, P.; Lechtreck, K.F. A Differential Cargo-Loading Model of Ciliary Length Regulation by IFT. Curr. Biol. 2013, 23, 2463–2471. [Google Scholar] [CrossRef] [Green Version]
- Pigino, G. Intraflagellar Transport. Curr. Biol. 2021, 31, R530–R536. [Google Scholar] [CrossRef]
- Piperno, G.; Mead, K.; Henderson, S. Inner Dynein Arms but Not Outer Dynein Arms Require the Activity of Kinesin Homologue Protein KHP1(FLA10) to Reach the Distal Part of Flagella in Chlamydomonas. J. Cell Biol. 1996, 133, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Snell, W.J. Kinesin II and Regulated Intraflagellar Transport of Chlamydomonas Aurora Protein Kinase. J. Cell Sci. 2003, 116, 2179–2186. [Google Scholar] [CrossRef] [Green Version]
- Goetz, S.C.; Anderson, K.V. The Primary Cilium: A Signalling Centre during Vertebrate Development. Nat. Rev. Genet. 2010, 11, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Keady, B.T.; Samtani, R.; Tobita, K.; Tsuchya, M.; San Agustin, J.T.; Follit, J.A.; Jonassen, J.A.; Subramanian, R.; Lo, C.W.; Pazour, G.J. IFT25 Links the Signal-Dependent Movement of Hedgehog Components to Intraflagellar Transport. Dev. Cell 2012, 22, 940–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liem, K.F.; Ashe, A.; He, M.; Satir, P.; Moran, J.; Beier, D.; Wicking, C.; Anderson, K.V. The IFT-A Complex Regulates Shh Signaling through Cilia Structure and Membrane Protein Trafficking. J. Cell Biol. 2012, 197, 789–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguether, T.; SanAgustin, J.T.; Keady, B.T.; Jonassen, J.A.; Liang, Y.; Francis, R.; Tobita, K.; Johnson, C.A.; Abdelhamed, Z.A.; Lo, C.W.; et al. IFT27 Links the BBSome to IFT for Maintenance of the Ciliary Signaling Compartment. Dev. Cell 2014, 31, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheway, G.; Nazlamova, L.; Hancock, J.T. Signaling through the Primary Cilium. Front. Cell Dev. Biol. 2018, 6, 8. [Google Scholar] [CrossRef]
- Pazour, G.J.; Witman, G.B. The Vertebrate Primary Cilium Is a Sensory Organelle. Curr. Opin. Cell Biol. 2003, 15, 105–110. [Google Scholar] [CrossRef]
- Lechtreck, K.F.; Johnson, E.C.; Sakai, T.; Cochran, D.; Ballif, B.A.; Rush, J.; Pazour, G.J.; Ikebe, M.; Witman, G.B. The Chlamydomonas Reinhardtii BBSome Is an IFT Cargo Required for Export of Specific Signaling Proteins from Flagella. J. Cell Biol. 2009, 187, 1117–1132. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.; Katoh, Y. Architecture of the IFT Ciliary Trafficking Machinery and Interplay between Its Components. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 179–196. [Google Scholar] [CrossRef]
- Christensen, S.T.; Morthorst, S.K.; Mogensen, J.B.; Pedersen, L.B. Primary Cilia and Coordination of Receptor Tyrosine Kinase (RTK) and Transforming Growth Factor β (TGF-β) Signaling. Cold Spring Harb. Perspect. Biol. 2017, 9, a028167. [Google Scholar] [CrossRef] [Green Version]
- Finetti, F.; Baldari, C.T. Compartmentalization of Signaling by Vesicular Trafficking: A Shared Building Design for the Immune Synapse and the Primary Cilium. Immunol. Rev. 2013, 251, 97–112. [Google Scholar] [CrossRef]
- De-Castro, A.R.G.; Quintas-Gonçalves, J.; Silva-Ribeiro, T.; Rodrigues, D.R.M.; De-Castro, M.J.G.; Abreu, C.M.; Dantas, T.J. The IFT20 Homolog in Caenorhabditis Elegans Is Required for Ciliogenesis and Cilia-Mediated Behavior. MicroPubl. Biol. 2021, 2021, 10. [Google Scholar] [CrossRef]
- Baker, S.A.; Freeman, K.; Luby-Phelps, K.; Pazour, G.J.; Besharse, J.C. IFT20 Links Kinesin II with a Mammalian Intraflagellar Transport Complex That Is Conserved in Motile Flagella and Sensory Cilia. J. Biol. Chem. 2003, 278, 34211–34218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omori, Y.; Zhao, C.; Saras, A.; Mukhopadhyay, S.; Kim, W.; Furukawa, T.; Sengupta, P.; Veraksa, A.; Malicki, J. Elipsa Is an Early Determinant of Ciliogenesis That Links the IFT Particle to Membrane-Associated Small GTPase Rab8. Nat. Cell Biol. 2008, 10, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Boukhalfa, A.; Roccio, F.; Dupont, N.; Codogno, P.; Morel, E. The Autophagy Protein ATG16L1 Cooperates with IFT20 and INPP5E to Regulate the Turnover of Phosphoinositides at the Primary Cilium. Cell Rep. 2021, 35, 109045. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.H.; Lin, Y.; Zhang, Z.G. Intraflagellar Transport 20: New Target for the Treatment of Ciliopathies. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118641. [Google Scholar] [CrossRef]
- Maharjan, Y.; Lee, J.N.; Kwak, S.A.; Dutta, R.K.; Park, C.; Choe, S.; Park, R. TMEM135 Regulates Primary Ciliogenesis through Modulation of Intracellular Cholesterol Distribution. EMBO Rep. 2020, 21, e48901. [Google Scholar] [CrossRef]
- Follit, J.A.; Tuft, R.A.; Fogarty, K.E.; Pazour, G.J. The Intraflagellar Transport Protein IFT20 Is Associated with the Golgi Complex and Is Required for Cilia Assembly. Mol. Biol. Cell 2006, 17, 3781–3792. [Google Scholar] [CrossRef] [Green Version]
- Finetti, F.; Patrussi, L.; Masi, G.; Onnis, A.; Galgano, D.; Lucherini, O.M.; Pazour, G.J.; Baldari, C.T. Specific Recycling Receptors Are Targeted to the Immune Synapse by the Intraflagellar Transport System. J. Cell Sci. 2014, 127, 1924–1937. [Google Scholar] [CrossRef] [Green Version]
- Nachury, M.V.; Loktev, A.V.; Zhang, Q.; Westlake, C.J.; Peränen, J.; Merdes, A.; Slusarski, D.C.; Scheller, R.H.; Bazan, J.F.; Sheffield, V.C.; et al. A Core Complex of BBS Proteins Cooperates with the GTPase Rab8 to Promote Ciliary Membrane Biogenesis. Cell 2007, 129, 1201–1213. [Google Scholar] [CrossRef] [Green Version]
- Knödler, A.; Feng, S.; Zhang, J.; Zhang, X.; Das, A.; Peränen, J.; Guo, W. Coordination of Rab8 and Rab11 in Primary Ciliogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 6346–6351. [Google Scholar] [CrossRef] [Green Version]
- Westlake, C.J.; Baye, L.M.; Nachury, M.V.; Wright, K.J.; Ervin, K.E.; Phu, L.; Chalouni, C.; Beck, J.S.; Kirkpatrick, D.S.; Slusarski, D.C.; et al. Primary Cilia Membrane Assembly Is Initiated by Rab11 and Transport Protein Particle II (TRAPPII) Complex-Dependent Trafficking of Rabin8 to the Centrosome. Proc. Natl. Acad. Sci. USA 2011, 108, 2759–2764. [Google Scholar] [CrossRef]
- Follit, J.A.; San Agustin, J.T.; Xu, F.; Jonassen, J.A.; Samtani, R.; Lo, C.W.; Pazour, G.J. The Golgin GMAP210/TRIP11 Anchors IFT20 to the Golgi Complex. PLoS Genet. 2008, 4, e1000315. [Google Scholar] [CrossRef] [PubMed]
- Joo, K.; Kim, C.G.; Lee, M.S.; Moon, H.Y.; Lee, S.H.; Kim, M.J.; Kweon, H.S.; Park, W.Y.; Kim, C.H.; Gleeson, J.G.; et al. CCDC41 Is Required for Ciliary Vesicle Docking to the Mother Centriole. Proc. Natl. Acad. Sci. USA 2013, 110, 5987–5992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, H.; Huang, K. Transport of Ciliary Membrane Proteins. Front. Cell Dev. Biol. 2020, 7, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roboti, P.; Sato, K.; Lowe, M. The Golgin GMAP-210 Is Required for Efficient Membrane Trafficking in the Early Secretory Pathway. J. Cell Sci. 2015, 128, 1595–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoetzel, C.; Bär, S.; de Craene, J.O.; Scheidecker, S.; Etard, C.; Chicher, J.; Reck, J.R.; Perrault, I.; Geoffroy, V.; Chennen, K.; et al. A Mutation in VPS15 (PIK3R4) Causes a Ciliopathy and Affects IFT20 Release from the Cis-Golgi. Nat. Commun. 2016, 7, 13586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Joo, K.; Jung, E.J.; Hong, H.; Seo, J.; Kim, J. Export of Membrane Proteins from the Golgi Complex to the Primary Cilium Requires the Kinesin Motor, KIFC1. FASEB J. 2018, 32, 957–968. [Google Scholar] [CrossRef] [Green Version]
- Moritz, O.L.; Tam, B.M.; Hurd, L.L.; Peränen, J.; Deretic, D.; Papermaster, D.S. Mutant Rab8 Impairs Docking and Fusion of Rhodopsin-Bearing Post-Golgi Membranes and Causes Cell Death of Transgenic Xenopus Rods. Mol. Biol. Cell 2001, 12, 2341–2351. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Insinna, C.; Ott, C.; Stauffer, J.; Pintado, P.A.; Rahajeng, J.; Baxa, U.; Walia, V.; Cuenca, A.; Hwang, Y.S.; et al. Early Steps in Primary Cilium Assembly Require EHD1/EHD3-Dependent Ciliary Vesicle Formation. Nat. Cell Biol. 2015, 17, 228–240. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Shao, L.; Yao, Y.; Tong, X.; Liu, H.; Yue, S.; Xie, L.; Cheng, S.Y. DGKδ Triggers Endoplasmic Reticulum Release of IFT88-Containing Vesicles Destined for the Assembly of Primary Cilia. Sci. Rep. 2017, 7, 5296. [Google Scholar] [CrossRef] [Green Version]
- Monis, W.J.; Faundez, V.; Pazour, G.J. BLOC-1 Is Required for Selective Membrane Protein Trafficking from Endosomes to Primary Cilia. J. Cell Biol. 2017, 216, 2131–2150. [Google Scholar] [CrossRef]
- Schmid, F.M.; Schou, K.B.; Vilhelm, M.J.; Holm, M.S.; Breslin, L.; Farinelli, P.; Larsen, L.A.; Andersen, J.S.; Pedersen, L.B.; Christensen, S.T. IFT20 Modulates Ciliary PDGFRα Signaling by Regulating the Stability of Cbl E3 Ubiquitin Ligases. J. Cell Biol. 2018, 217, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Mitchell, D.R.; Jékely, G. Chapter 3 How Did the Cilium Evolve. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2008; Volume 85, pp. 63–82. [Google Scholar]
- Finetti, F.; Paccani, S.R.; Riparbelli, M.G.; Giacomello, E.; Perinetti, G.; Pazour, G.J.; Rosenbaum, J.L.; Baldari, C.T. Intraflagellar Transport Is Required for Polarized Recycling of the TCR/CD3 Complex to the Immune Synapse. Nat. Cell Biol. 2009, 11, 1332–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedmak, T.; Wolfrum, U. Intraflagellar Transport Molecules in Ciliary and Nonciliary Cells of the Retina. J. Cell Biol. 2010, 189, 171–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Begum, S.; Ezratty, E.J. An IFT20 Mechanotrafficking Axis Is Required for Integrin Recycling, Focal Adhesion Dynamics, and Polarized Cell Migration. Mol. Biol. Cell 2020, 31, 1917–1930. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, F.; Long, H.; Lin, Y.; Liao, J.; Xia, H.; Huang, K. IFT20 Mediates the Transport of Cell Migration Regulators From the Trans-Golgi Network to the Plasma Membrane in Breast Cancer Cells. Front. Cell Dev. Biol. 2021, 9, 632198. [Google Scholar] [CrossRef]
- Liu, H.; Rhodes, M.; Wiest, D.L.; Vignali, D.A. On the Dynamics of TCR:CD3 Complex Cell Surface Expression and Downmodulation. Immunity 2000, 13, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kremer, K.N.; Dominguez, D.; Tadi, M.; Hedin, K.E. G 13 and Rho Mediate Endosomal Trafficking of CXCR4 into Rab11+ Vesicles upon Stromal Cell-Derived Factor-1 Stimulation. J. Immunol. 2011, 186, 951–958. [Google Scholar] [CrossRef] [Green Version]
- Mayle, K.M.; Le, A.M.; Kamei, D.T. The Intracellular Trafficking Pathway of Transferrin. Biochim. Biophys. Acta 2012, 1820, 264–281. [Google Scholar] [CrossRef] [Green Version]
- Grant, B.D.; Donaldson, J.G. Pathways and Mechanisms of Endocytic Recycling. Nat. Rev. Mol. Cell Biol. 2009, 10, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Onnis, A.; Baldari, C.T. Orchestration of Immunological Synapse Assembly by Vesicular Trafficking. Orchestration of Immunological Synapse Assembly by Vesicular Trafficking. Front. Cell Dev. Biol. 2019, 7, 110. [Google Scholar] [CrossRef]
- Vivar, O.I.; Masi, G.; Carpier, J.; Magalhaes, J.G.; Galgano, D.; Pazour, G.J.; Amigorena, S.; Hivroz, C.; Baldari, C.T. IFT20 Controls LAT Recruitment to the Immune Synapse and T-Cell Activation in Vivo. Proc. Natl. Acad. Sci. USA 2016, 113, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Zucchetti, A.E.; Bataille, L.; Carpier, J.M.; Dogniaux, S.; San Roman-Jouve, M.; Maurin, M.; Stuck, M.W.; Rios, R.M.; Baldari, C.T.; Pazour, G.J.; et al. Tethering of Vesicles to the Golgi by GMAP210 Controls LAT Delivery to the Immune Synapse. Nat. Commun. 2019, 10, 2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galgano, D.; Onnis, A.; Pappalardo, E.; Galvagni, F.; Acuto, O.; Baldari, C.T. The T Cell IFT20 Interactome Reveals New Players in Immune Synapse Assembly. J. Cell Sci. 2017, 130, 1110–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Garrett-Sinha, L.A.; Sarkar, D.; Yang, S. Deletion of IFT20 in Early Stage T Lymphocyte Differentiation Inhibits the Development of Collagen-Induced Arthritis. Bone Res. 2014, 2, 14038. [Google Scholar] [CrossRef] [Green Version]
- Aoki, T.; Nishita, M.; Sonoda, J.; Ikeda, T.; Kakeji, Y.; Minami, Y. Intraflagellar Transport 20 Promotes Collective Cancer Cell Invasion by Regulating Polarized Organization of Golgi-Associated Microtubules. Cancer Sci. 2019, 110, 1306–1316. [Google Scholar] [CrossRef]
- Nishita, M.; Park, S.Y.; Nishio, T.; Kamizaki, K.; Wang, Z.; Tamada, K.; Takumi, T.; Hashimoto, R.; Otani, H.; Pazour, G.J.; et al. Ror2 Signaling Regulates Golgi Structure and Transport through IFT20 for Tumor Invasiveness. Sci. Rep. 2017, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Castro-Castro, A.; Marchesin, V.; Monteiro, P.; Lodillinsky, C.; Rossé, C.; Chavrier, P. Cellular and Molecular Mechanisms of MT1-MMP-Dependent Cancer Cell Invasion. Annu. Rev. Cell Dev. Biol. 2016, 32, 555–576. [Google Scholar] [CrossRef] [PubMed]
- Boehlke, C.; Janusch, H.; Hamann, C.; Powelske, C.; Mergen, M.; Herbst, H.; Kotsis, F.; Nitschke, R.; Kuehn, E.W. A Cilia Independent Role of Ift88/Polaris during Cell Migration. PLoS ONE 2015, 10, e0140378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rønn Veland, I.; Lindbaek, L.; Christensen, S.T. Linking the Primary Cilium to Cell Migration in Tissue Repair and Brain Development. BioScience 2014, 64, 1115. [Google Scholar] [CrossRef] [PubMed]
- Cassioli, C.; Baldari, C.T. A Ciliary View of the Immunological Synapse. Cells 2019, 8, 789. [Google Scholar] [CrossRef] [PubMed]
- Peralta, M.; Ortiz Lopez, L.; Jerabkova, K.; Lucchesi, T.; Vitre, B.; Han, D.; Guillemot, L.; Dingare, C.; Sumara, I.; Mercader, N.; et al. Intraflagellar Transport Complex B Proteins Regulate the Hippo Effector Yap1 during Cardiogenesis. Cell Rep. 2020, 32, 107932. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, S.; Heallen, T.; Martin, J.F. The Hippo Pathway in the Heart: Pivotal Roles in Development, Disease, and Regeneration. Nat. Rev. Cardiol. 2018, 15, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Clement, C.A.; Kristensen, S.G.; Møllgard, K.; Pazour, G.J.; Yoder, B.K.; Larsen, L.A.; Christensen, S.T. The Primary Cilium Coordinates Early Cardiogenesis and Hedgehog Signaling in Cardiomyocyte Differentiation. J. Cell Sci. 2009, 122, 3070–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finetti, F.; Capitani, N.; Baldari, C.T. Emerging Roles of the Intraflagellar Transport System in the Orchestration of Cellular Degradation Pathways. Front. Cell Dev. Biol. 2019, 7, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pampliega, O.; Orhon, I.; Patel, B.; Sridhar, S.; Díaz-Carretero, A.; Beau, I.; Codogno, P.; Satir, B.H.; Satir, P.; Cuervo, A.M. Functional Interaction between Autophagy and Ciliogenesis. Nature 2013, 502, 194–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in Major Human Diseases. EMBO J. 2021, 40, 108863. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy Pathway: Cellular and Molecular Mechanisms. Autophagy 2018, 14, 207. [Google Scholar] [CrossRef] [Green Version]
- Orhon, I.; Dupont, N.; Zaidan, M.; Boitez, V.; Burtin, M.; Schmitt, A.; Capiod, T.; Viau, A.; Beau, I.; Wolfgang Kuehn, E.; et al. Primary-Cilium-Dependent Autophagy Controls Epithelial Cell Volume in Response to Fluid Flow. Nat. Cell Biol. 2016, 18, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Cassioli, C.; Cianfanelli, V.; Zevolini, F.; Onnis, A.; Gesualdo, M.; Brunetti, J.; Cecconi, F.; Baldari, C.T. The Intraflagellar Transport Protein IFT20 Recruits ATG16L1 to Early Endosomes to Promote Autophagosome Formation in T Cells. Front. Cell Dev. Biol. 2021, 9, 634003. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Cassioli, C.; Cianfanelli, V.; Onnis, A.; Paccagnini, E.; Kabanova, A.; Baldari, C.T. The Intraflagellar Transport Protein IFT20 Controls Lysosome Biogenesis by Regulating the Post-Golgi Transport of Acid Hydrolases. Cell Death Differ. 2020, 27, 310–328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, W.; Zhang, Y.; Zhang, L.; Teves, M.E.; Liu, H.; Strauss, J.F.; Pazour, G.J.; Foster, J.A.; Hess, R.A.; et al. Intraflagellar Transport Protein IFT20 Is Essential for Male Fertility and Spermiogenesis in Mice. Mol. Biol. Cell 2016, 27, 3705–3716. [Google Scholar] [CrossRef] [PubMed]
- Noda, K.; Kitami, M.; Kitami, K.; Kaku, M.; Komatsu, Y. Canonical and Noncanonical Intraflagellar Transport Regulates Craniofacial Skeletal Development. Proc. Natl. Acad. Sci. USA 2016, 113, E2589–E2597. [Google Scholar] [CrossRef] [Green Version]
- Boyadjiev, S.A.; Fromme, J.C.; Ben, J.; Chong, S.S.; Nauta, C.; Hur, D.J.; Zhang, G.; Hamamoto, S.; Schekman, R.; Ravazzola, M.; et al. Cranio-Lenticulo-Sutural Dysplasia Is Caused by a SEC23A Mutation Leading to Abnormal Endoplasmic-Reticulum-to-Golgi Trafficking. Nat. Genet. 2006, 38, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Boyadjiev, S.; Kim, S.D.; Hata, A.; Haldeman-Englert, C.; Zackai, E.; Naydenov, C.; Hamamoto, S.; Schekman, R.; Kim, J. Cranio-Lenticulo-Sutural Dysplasia Associated with Defects in Collagen Secretion. Clin. Genet. 2011, 80, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, P.; Bolton, A.D.; Funari, V.; Hong, M.; Boyden, E.D.; Lu, L.; Manning, D.K.; Dwyer, N.D.; Moran, J.L.; Prysak, M.; et al. Lethal Skeletal Dysplasia in Mice and Humans Lacking the Golgin GMAP-210. N. Engl. J. Med. 2010, 362, 206–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernet-Gallay, K.; Antony, C.; Johannes, L.; Bornens, M.; Goud, B.; Rios, R.M. The Overexpression of GMAP-210 Blocks Anterograde and Retrograde Transport between the ER and the Golgi Apparatus. Traffic 2002, 3, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Kitami, M.; Yamaguchi, H.; Ebina, M.; Kaku, M.; Chen, D.; Komatsu, Y. IFT20 Is Required for the Maintenance of Cartilaginous Matrix in Condylar Cartilage. Biochem. Biophys. Res. Commun. 2019, 509, 222–226. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Terajima, M.; Kitami, M.; Wang, J.; He, L.; Saeki, M.; Yamauchi, M.; Komatsu, Y. IFT20 Is Critical for Collagen Biosynthesis in Craniofacial Bone Formation HHS Public Access. Biochem. Biophys. Res. Commun. 2020, 533, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Li, X.; Yuan, X.; Yang, S.; Han, L.; Yang, S. Primary Cilia Control Cell Alignment and Patterning in Bone Development via Ceramide-PKCζ-β-Catenin Signaling. Commun. Biol. 2020, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.W.; Cho, J.H.; Conway, H.E.; DiGruccio, M.R.; Ng, X.W.; Roseman, H.F.; Abreu, D.; Urano, F.; Piston, D.W. Primary Cilia Control Glucose Homeostasis via Islet Paracrine Interactions. Proc. Natl. Acad. Sci. USA 2020, 117, 8912–8923. [Google Scholar] [CrossRef]
- Volta, F.; Scerbo, M.J.; Seelig, A.; Wagner, R.; O’Brien, N.; Gerst, F.; Fritsche, A.; Häring, H.U.; Zeigerer, A.; Ullrich, S.; et al. Glucose Homeostasis Is Regulated by Pancreatic β-Cell Cilia via Endosomal EphA-Processing. Nat. Commun. 2019, 10, 5686. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yang, S.; Liu, Y.; Qin, L.; Yang, S. IFT20 Governs Mesenchymal Stem Cell Fate through Positively Regulating TGF-β-Smad2/3-Glut1 Signaling Mediated Glucose Metabolism. Redox Biol. 2022, 54, 102373. [Google Scholar] [CrossRef]
- Jonassen, J.A.; Agustin, J.S.; Follit, J.A.; Pazour, G.J. Deletion of IFT20 in the Mouse Kidney Causes Misorientation of the Mitotic Spindle and Cystic Kidney Disease. J. Cell Biol. 2008, 183, 377–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.; Kaci, N.; Estibals, V.; Goudin, N.; Garfa-Traore, M.; Benoist-Lasselin, C.; Dambroise, E.; Legeai-Mallet, L. Constitutively-Active FGFR3 Disrupts Primary Cilium Length and IFT20 Trafficking in Various Chondrocyte Models of Achondroplasia. Hum. Mol. Genet. 2018, 27, 1–13. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Kitami, M.; Uchima Koecklin, K.H.; He, L.; Wang, J.; Lagor, W.R.; Perrien, D.S.; Komatsu, Y. Temporospatial Regulation of Intraflagellar Transport Is Required for the Endochondral Ossification in Mice. Dev. Biol. 2022, 482, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Kfoury, Y.; Scadden, D.T. Mesenchymal Cell Contributions to the Stem Cell Niche. Cell Stem Cell 2015, 16, 239–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chen, Y.; Liao, W.; Yu, Q.; Lin, H.; Shi, Y.; Zhang, L.; Fu, G.; Wang, Z.; Li, X.; et al. Associations of IFT20 and GM130 Protein Expressions with Clinicopathological Features and Survival of Patients with Lung Adenocarcinoma. BMC Cancer 2022, 22, 809. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finetti, F.; Onnis, A.; Baldari, C.T. IFT20: An Eclectic Regulator of Cellular Processes beyond Intraflagellar Transport. Int. J. Mol. Sci. 2022, 23, 12147. https://doi.org/10.3390/ijms232012147
Finetti F, Onnis A, Baldari CT. IFT20: An Eclectic Regulator of Cellular Processes beyond Intraflagellar Transport. International Journal of Molecular Sciences. 2022; 23(20):12147. https://doi.org/10.3390/ijms232012147
Chicago/Turabian StyleFinetti, Francesca, Anna Onnis, and Cosima T. Baldari. 2022. "IFT20: An Eclectic Regulator of Cellular Processes beyond Intraflagellar Transport" International Journal of Molecular Sciences 23, no. 20: 12147. https://doi.org/10.3390/ijms232012147
APA StyleFinetti, F., Onnis, A., & Baldari, C. T. (2022). IFT20: An Eclectic Regulator of Cellular Processes beyond Intraflagellar Transport. International Journal of Molecular Sciences, 23(20), 12147. https://doi.org/10.3390/ijms232012147