
Anti-Allergic Activity of Fucoidan Can Be Enhanced by Coexistence with Quercetin
 , ,
, ,
Abstract
:1. Introduction
2. Results
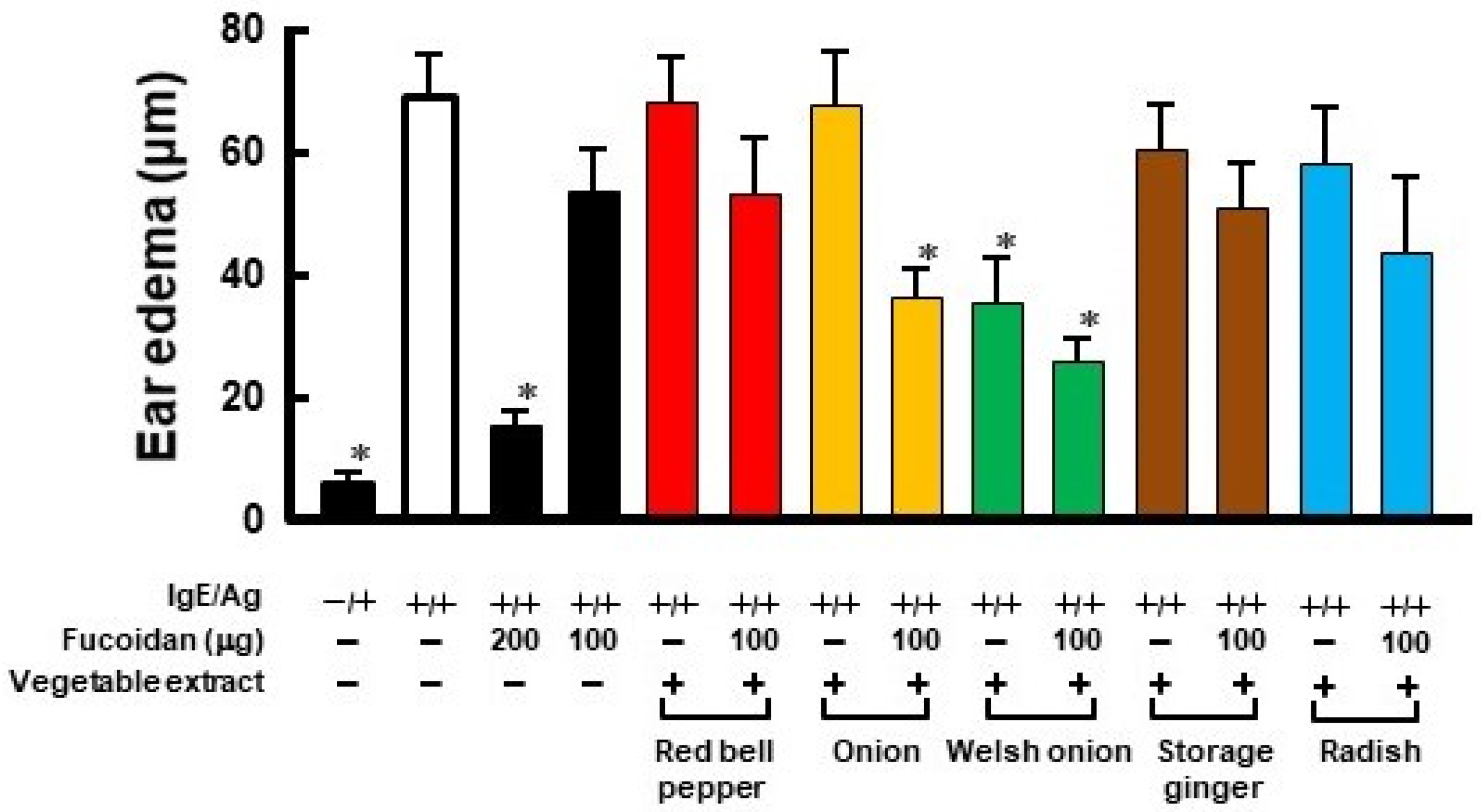
2.1. Anti-Allergic Effect Using a Combination of Fucoidan and Vegetable Extract in Passive Cutaneous Anaphylaxis Reaction
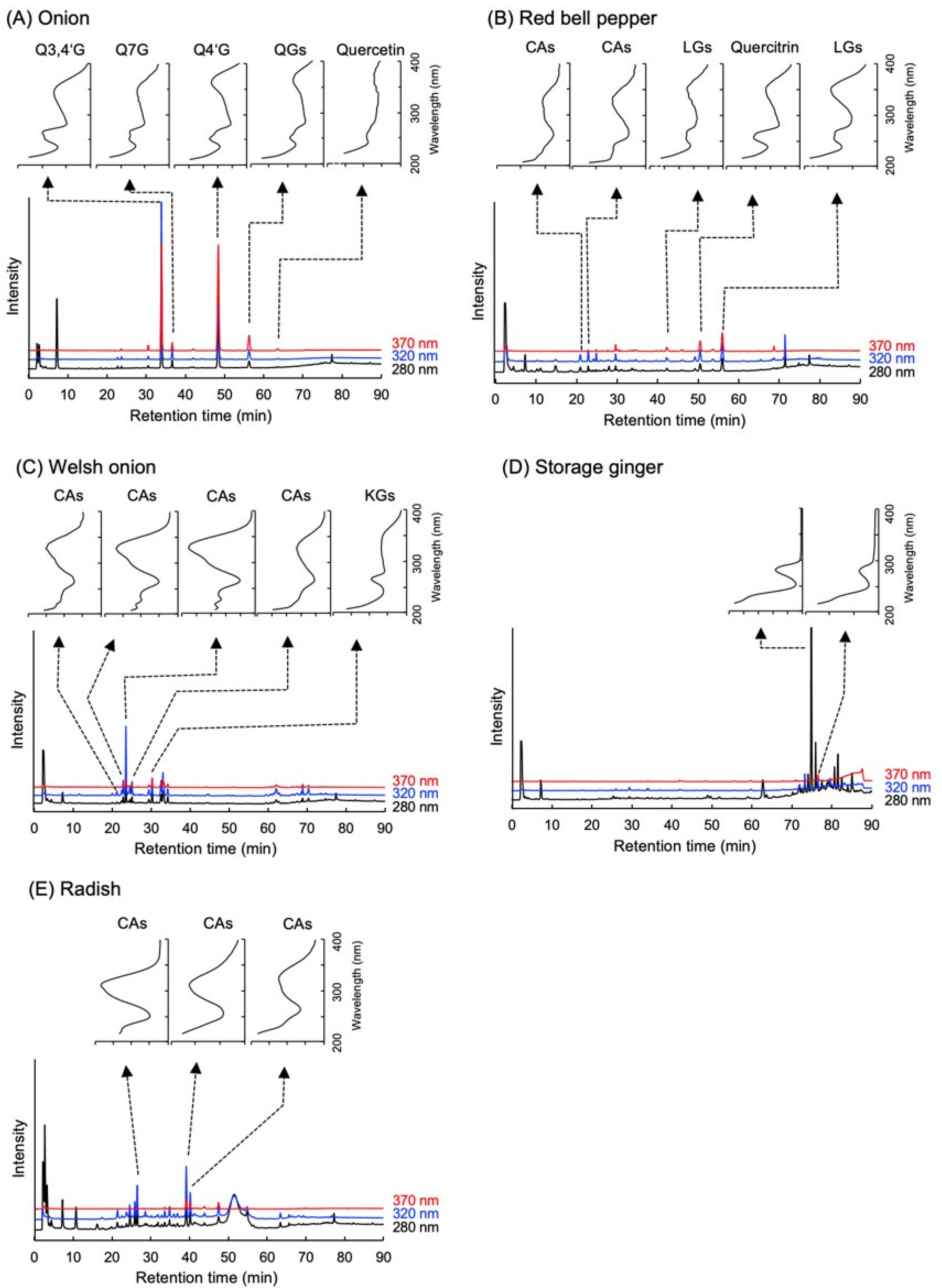
2.2. Polyphenol Contents in Various Vegetables
2.3. Anti-Allergic Activity of Coexistence of Fucoidan and Quercetin or Kaempferol in PCA Reaction
2.4. Involvement of Gal9 in Anti-Allergic Activity by Orally Simultaneous Administration of Fucoidan and Quercetin
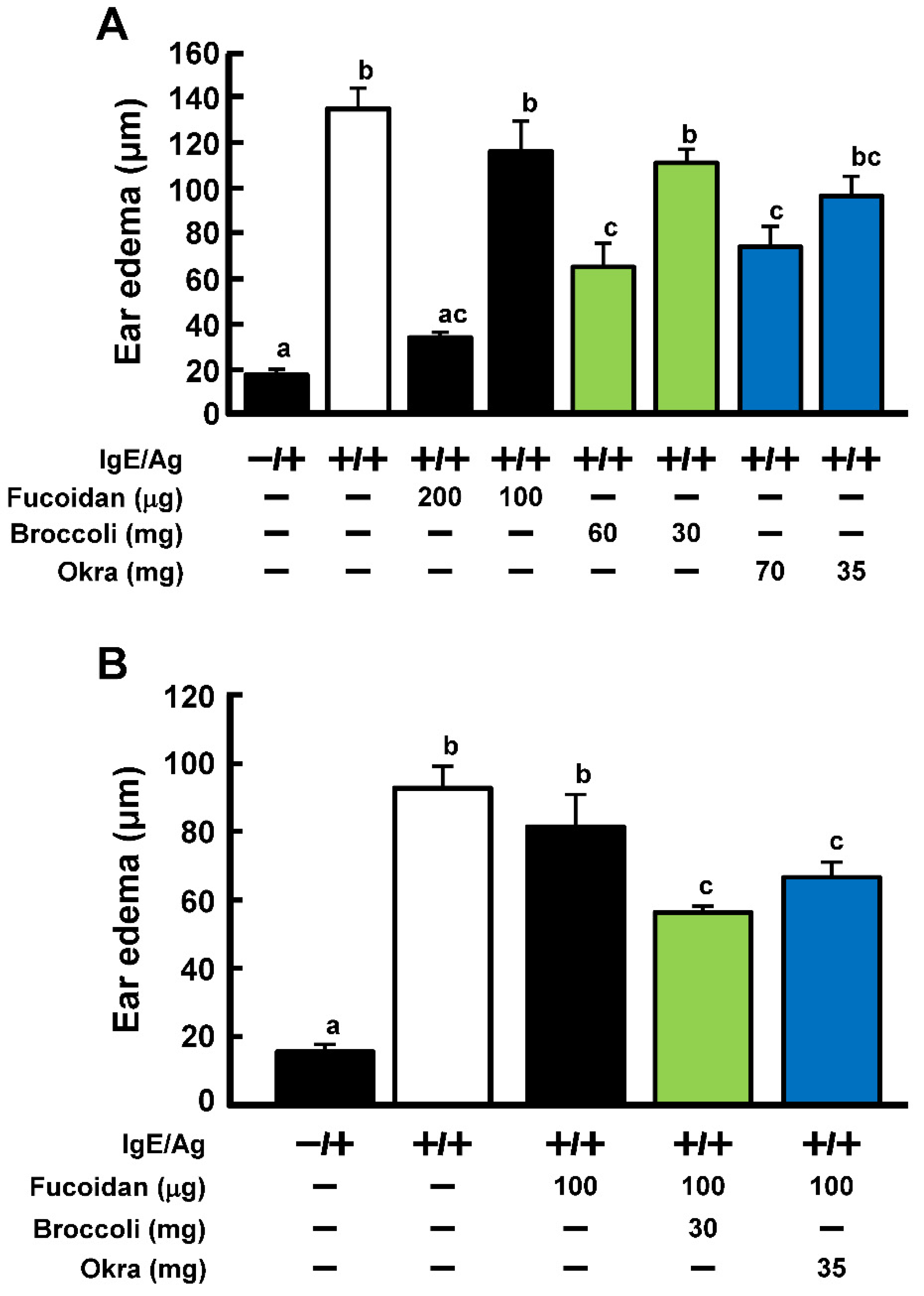
2.5. Effect of Extract from Broccoli and Okra on Anti-Allergic Activity of Fucoidan in PCA Reaction
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Mice
4.3. Passive Cutaneous Anaphylaxis Reaction
4.4. Extraction of Polyphenol Fractions from Vegetables
4.5. Analyses of Polyphenol Contents in Vegetables by High Performance Liquid Chromatography (HPLC)-Diode Array Detector (DAD)
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DAD | diode array detector |
| Gal9 | galectin-9 |
| HPLC | high performance liquid chromatography |
| PCA | passive cutaneous anaphylaxis |
| TNP | 2,4,6-trinitrophenyl |
References
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. Consorzio Interuniversitario Nazionale per la Bio-Oncologia, Italy, A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. The effect of sulfated (1→3)-α-L-fucan from the brown alga Saccharina cichorioides miyabe on resveratrol-induced apoptosis in colon carcinoma cells. Mar. Drugs 2013, 11, 194–212. [Google Scholar] [CrossRef] [Green Version]
- Nishino, T.; Nagumo, T.; Kiyohara, H.; Yamada, H. Structural characterization of a new anticoagulant fucan sulfate from the brown seaweed Ecklonia kurome. Carbohyd. Res. 1991, 211, 77–90. [Google Scholar] [CrossRef]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.O. Fucoidan from Macrocystis pyrifera has powerful Immune-modulatory effects compared to three other fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, H.; Tamauchi, H.; Hashimoto, M.; Nakano, T. Suppression of Th2 immune responses by Mekabu fucoidan from Undaria pinnatifida sporophylls. Int. Arch. Aller. Immunol. 2005, 137, 289–294. [Google Scholar] [CrossRef]
- Iwamoto, K.; Hiragun, T.; Takahagi, S.; Yanase, Y.; Morioke, S.; Mihara, S.; Kameyoshi, Y.; Hide, M. Fucoidan suppresses IgE production in peripheral blood mononuclear cells from patients with atopic dermatitis. Arch. Dermatol. Res. 2011, 303, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Tanino, Y.; Hashimoto, T.; Ojima, T.; Mizuno, M. F-fucoidan from Saccharina japonica is a novel inducer of galectin-9 and exhibits anti-allergic activity. J. Clin. Biochem. Nutr. 2016, 59, 25–30. [Google Scholar] [CrossRef]
- Mizuno, M.; Sakaguchi, K.; Sakane, I. Oral administration of fucoidan can exert anti-allergic activity after allergen sensitization by enhancement of galectin-9 secretion in blood. Biomolecules 2020, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, M.; Ray, S.K. Synergistic anti-tumor actions of luteolin and silibinin prevented cell migration an invasion and induced apoptosis in glioblastoma SNB19 cells and glioblastoma stem cells. Brain Res. 2015, 1629, 85–93. [Google Scholar] [CrossRef]
- Mizuno, M.; Yamashita, S.; Hashimoto, T. Enhancement of anti-inflammatory and anti-allergic activities with combination of luteolin and quercetin in in vitro co-culture system. Food Sci. Technol. Res. 2017, 23, 811–818. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Sakane, I.; Mizuno, M. Synergistic anti-allergy activity using combination of Enterococcus faecalis IC-1 and luteolin. Food Biosci. 2021, 41, 100924. [Google Scholar] [CrossRef]
- Ovary, Z. In vitro and in vivo interaction of anti-hapten antibodies with monovalent and bivalent haptens. In Conceptual Advances in Immunology and Oncology; Cumley, R.W., Aldridge, D.M., MacCay, J.H.J., Eds.; Harvert Medical Division, Harper and Row: New York, NY, USA, 1963; pp. 206–219. [Google Scholar]
- Singh, A.; Holvoet, S.; Mercenier, A. Dietary polyphenols in the prevention and treatment of allergic diseases. Clin. Exp. Allergy 2011, 41, 1346–1359. [Google Scholar] [CrossRef]
- Kimata, M.; Shichijo, M.; Miura, T.; Serizawa, I.; Inagaki, N.; Nagai, H. Effects of luteolin, quercetin and baicalein on immunoglobulin E-mediated mediator release from human cultured mast cells. Clin. Exp. Allergy 2000, 30, 501–508. [Google Scholar] [CrossRef]
- Cruz, E.A.; Reuter, S.; Martin, H.; Dehzad, N.; Muzitano, M.F.; Costa, S.S.; Rossi-Bermann, B.; Buhl, R.; Stassen, M.; Taube, C. Kalanchoe pinnata inhibits mast cell activation and prevents allergic airway disease. Phytomedicine 2012, 19, 115–121. [Google Scholar] [CrossRef]
- Baolin, L.; Inami, Y.; Tanaka, H.; Inagaki, N.; Iinuma, M.; Nagai, H. Resveratrol inhibits the release of mediators from bone marrow-derived mouse mast cells in vitro. Planta Med. 2004, 70, 305–309. [Google Scholar]
- Yamashita, S.; Yokoyama, Y.; Hashimoto, T.; Mizuno, M. A novel in vitro co-culture model comprised of Coco-2/RBL-2H3 cells to evaluate anti-allergic effects of food factors through the intestine. J. Immunol. Methods 2016, 435, 1–6. [Google Scholar] [CrossRef]
- Cao, J.; Li, C.; Ma, P.; Ding, Y.; Gao, J.; Jia, Q.; Zhu, J.; Zhang, T. Effect of kaempferol on IgE-mediated anaphylaxis in C57BL/6 mice and LAD2 cells. Phytomedicine 2020, 79, 153346. [Google Scholar] [CrossRef]
- Pellow, J.; Nolte, A.; Temane, A.; Solomon, E.M. Health supplements for allergic rhinitis: A mixed-methods systematic review. Complement. Ther. Med. 2020, 51, 102425. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Honda, Y.; Nakagawa, S.; Ashida, H.; Kanazawa, K. Simultaneous determination of all polyphenols in vegetables, fruits, and teas. J. Agric. Food Chem. 2003, 51, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, H.; Tanaka, W.; Miyoshi, N.; Miyazaki, T.; Michimoto, H.; Sakakibara, H. Beneficial effects of the consumption of sun-dried radishes (Paphanus sativus cv. YR-Hyuga-Risou) on dyslipidemia in apolipoprotein E-deficient mice. J. Food Biochem. 2021, 45, e13727. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
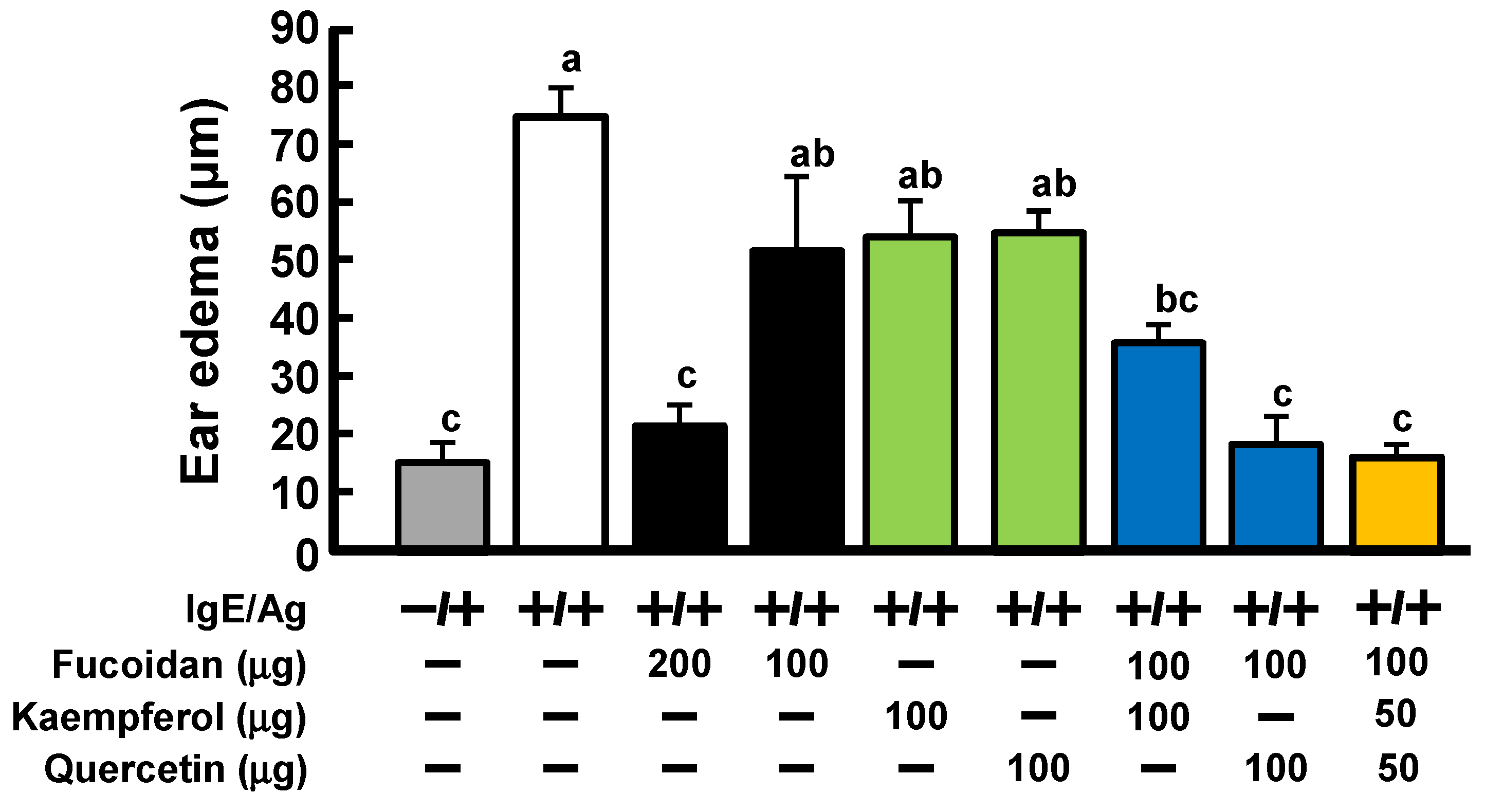{kind=link}
{kind=link}
{kind=link}
| Flavonoids | Phenolic Acids | |||
|---|---|---|---|---|
| μmol/g Extracts | μmol/g Extracts | |||
| Onion | quercetin-3,4’-diglucosides | 4.81 | – | |
| quercetin-4’-glucoside | 5.30 | |||
| quercetin-7-glucoside | 0.20 | |||
| other quercetin glucosides | 1.18 | |||
| quercetin | 0.07 | |||
| Red bell pepper | quercetin-3-rhamnoside | 0.77 | cinnamic acids | 0.32 |
| luteolin glucosides | 1.99 | |||
| Welsh onion | kaempferol glycosides | 3.15 | cinnamic acids | 3.36 |
| Storage ginger | – | – | ||
| Radish | – | cinnamic acids | 2.58 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizuno, M.; Fujioka, A.; Bitani, S.; Minato, K.-i.; Sakakibara, H. Anti-Allergic Activity of Fucoidan Can Be Enhanced by Coexistence with Quercetin. Int. J. Mol. Sci. 2022, 23, 12163. https://doi.org/10.3390/ijms232012163
Mizuno M, Fujioka A, Bitani S, Minato K-i, Sakakibara H. Anti-Allergic Activity of Fucoidan Can Be Enhanced by Coexistence with Quercetin. International Journal of Molecular Sciences. 2022; 23(20):12163. https://doi.org/10.3390/ijms232012163
Chicago/Turabian StyleMizuno, Masashi, Asuka Fujioka, Shiho Bitani, Ken-ichiro Minato, and Hiroyuki Sakakibara. 2022. "Anti-Allergic Activity of Fucoidan Can Be Enhanced by Coexistence with Quercetin" International Journal of Molecular Sciences 23, no. 20: 12163. https://doi.org/10.3390/ijms232012163