

Altered Expression of AQP1 and AQP4 in Brain Barriers and Cerebrospinal Fluid May Affect Cerebral Water Balance during Chronic Hypertension

 , , , and
, , , and
Abstract
:1. Introduction
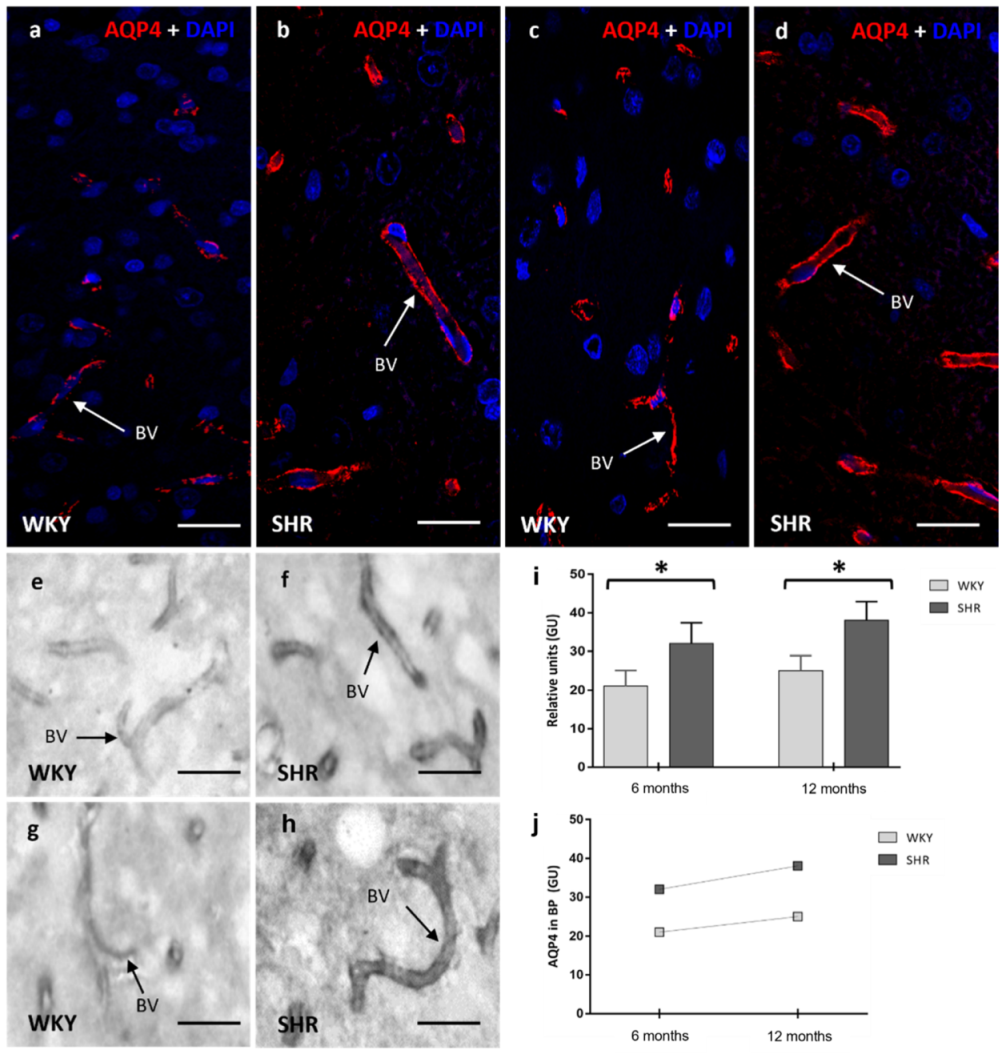
2. Results
2.1. AQP1 in ChP Epithelium (BCSFB)
2.2. AQP4 in Ependymal Cells and Subependymal Astrocytes (Brain–CSF Interface)
2.3. AQP4 in Astrocytes (BBB)
2.4. AQP1 and AQP4 in CSF
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cerebrospinal Fluid Extraction
4.3. Immunofluorescence and Immunohistochemistry
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Western Blotting
4.6. Image Acquisition and Immunofluorescence-Quantification
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SHR | Spontaneously hypertensive rat |
| AQP | Aquaporin |
| CSF | Cerebrospinal fluid |
| ChP | Choroid plexus |
| BBB | Blood–brain barrier |
| BCSFB | Blood–cerebrospinal fluid barrier |
| HT | Hypertension |
| WKY | Wistar–Kyoto |
| BP | Blood pressure |
| EVs | Extracellular vesicle |
References
- Johanson, C.E.; Duncan, J.A., 3rd; Klinge, P.M.; Brinker, T.; Stopa, E.G.; Silverberg, G.D. Multiplicity of cerebrospinal fluid functions: New challenges in health and disease. Cereb. Fluid Res. 2008, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Orešković, D.; Radoš, M.; Klarica, M. Role of choroid plexus in cerebrospinal fluid hydrodynamics. Neuroscience 2017, 354, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Trillo-Contreras, J.L.; Toledo-Aral, J.J.; Echevarría, M.; Villadiego, J. AQP1 and AQP4 Contribution to Cerebrospinal Fluid Homeostasis. Cells 2019, 8, 197. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. The glymphatic pathway in neurological disorders. Lancet Neurol. 2018, 17, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin water channels in the nervous system. Nat. Rev. Neurosci. 2013, 14, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanotto, C.; Simão, F.; Gasparin, M.S.; Biasibetti, R.; Tortorelli, L.S.; Nardin, P.; Gonçalves, C.A. Exendin-4 Reverses Biochemical and Functional Alterations in the Blood-Brain and Blood-CSF Barriers in Diabetic Rats. Mol. Neurobiol. 2017, 54, 2154–2166. [Google Scholar] [CrossRef] [PubMed]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Meyer, G.; Castañeyra-Perdomo, A. A Distal to Proximal Gradient of Human Choroid Plexus Development, with Antagonistic Expression of Glut1 and AQP1 in Mature Cells vs. Calbindin and PCNA in Proliferative Cells. Front. Neuroanat. 2016, 10, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, S.; Smith, B.L.; Christensen, E.I.; Agre, P. Distribution of the aquaporin CHIP in secretory and resorptive epithelia and capillary endothelia. Proc. Natl. Acad. Sci. USA 1993, 90, 7275–7279. [Google Scholar] [CrossRef] [Green Version]
- Speake, T.; Freeman, L.J.; Brown, P.D. Expression of aquaporin 1 and aquaporin 4 water channels in rat choroid plexus. Biochim. Biophys. Acta 2003, 1609, 80–86. [Google Scholar] [CrossRef]
- Verbavatz, J.M.; Ma, T.; Gobin, R.; Verkman, A.S. Absence of orthogonal arrays in kidney, brain and muscle from transgenic knockout mice lacking water channel aquaporin-4. J. Cell Sci. 1997, 110 Pt 22, 2855–2860. [Google Scholar] [CrossRef] [PubMed]
- Rash, J.E.; Yasumura, T.; Hudson, C.S.; Agre, P.; Nielsen, S. Direct immunogold labeling of aquaporin-4 in square arrays of astrocyte and ependymocyte plasma membranes in rat brain and spinal cord. Proc. Natl. Acad. Sci. USA 1998, 95, 11981–11986. [Google Scholar] [CrossRef] [Green Version]
- Al-Sarraf, H.; Philip, L. Effect of hypertension on the integrity of blood brain and blood CSF barriers, cerebral blood flow and CSF secretion in the rat. Brain Res. 2003, 975, 179–188. [Google Scholar] [CrossRef]
- Veglio, F.; Paglieri, C.; Rabbia, F.; Bisbocci, D.; Bergui, M.; Cerrato, P. Hypertension and cerebrovascular damage. Atherosclerosis 2009, 205, 331–341. [Google Scholar] [CrossRef] [PubMed]
- González-Marrero, I.; Castañeyra-Ruiz, L.; González-Toledo, J.M.; Castañeyra-Ruiz, A.; de Paz-Carmona, H.; Castro, R.; Hernandez-Fernaud, J.R.; Castañeyra-Perdomo, A.; Carmona-Calero, E.M. High blood pressure effects on the blood to cerebrospinal fluid barrier and cerebrospinal fluid protein composition: A two-dimensional electrophoresis study in spontaneously hypertensive rats. Int. J. Hypertens. 2013, 2013, 164653. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Marrero, I.; Hernández-Abad, L.G.; Castañeyra-Ruiz, L.; Carmona-Calero, E.M.; Castañeyra-Perdomo, A. Changes in the choroid plexuses and brain barriers associated with high blood pressure and ageing. Neurologia 2022, 37, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Al-Sarraf, H.; Ghaaedi, F.; Redzic, Z. Time course of hyperosmolar opening of the blood-brain and blood-CSF barriers in spontaneously hypertensive rats. J. Vasc. Res. 2007, 44, 99–109. [Google Scholar] [CrossRef]
- Ritter, S.; Dinh, T.T. Progressive postnatal dilation of brain ventricles in spontaneously hypertensive rats. Brain Res. 1986, 370, 327–332. [Google Scholar] [CrossRef]
- Bendel, P.; Eilam, R. Quantitation of ventricular size in normal and spontaneously hypertensive rats by magnetic resonance imaging. Brain Res. 1992, 574, 224–228. [Google Scholar] [CrossRef]
- Tajima, A.; Hans, F.J.; Livingstone, D.; Wei, L.; Finnegan, W.; DeMaro, J.; Fenstermacher, J. Smaller local brain volumes and cerebral atrophy in spontaneously hypertensive rats. Hypertension 1993, 21, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Peña y Valenzuela, I.; Carmona-Calero, E.M.; Pérez-González, H.; Ormazabal-Ramos, C.; Fernández-Rodríguez, P.; González-Marrero, I.; Castañeyra-Perdomo, A.; Ferres-Torres, R. Alterations of the cerebrospinal fluid proteins and subcommissural organ secretion in the arterial hypertension and ventricular dilatation. A study in SHR rats. Histol. Histopathol. 2006, 21, 179–185. [Google Scholar]
- Gu, C.; Hao, X.; Li, J.; Hua, Y.; Keep, R.F.; Xi, G. Effects of minocycline on epiplexus macrophage activation, choroid plexus injury and hydrocephalus development in spontaneous hypertensive rats. J. Cereb. Blood Flow Metab. 2019, 39, 1936–1948. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Ye, F.; Holste, K.G.; Hua, Y.; Garton, H.J.L.; Keep, R.F.; Xi, G. Delayed Minocycline Treatment Ameliorates Hydrocephalus Development and Choroid Plexus Inflammation in Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2022, 23, 2306. [Google Scholar] [CrossRef]
- Abbott, N.J.; Pizzo, M.E.; Preston, J.E.; Janigro, D.; Thorne, R.G. The role of brain barriers in fluid movement in the CNS: Is there a ‘glymphatic’ system? Acta Neuropathol. 2018, 135, 387–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Papadopoulos, M.C.; Liu, J.; Li, L.; Zhang, D.; Zhang, H.; Verkman, A.S.; Ma, T. Sporadic obstructive hydrocephalus in Aqp4 null mice. J. Neurosci. Res. 2009, 87, 1150–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestre, H.; Hablitz, L.M.; Xavier, A.L.; Feng, W.; Zou, W.; Pu, T.; Monai, H.; Murlidharan, G.; Castellanos Rivera, R.M.; Simon, M.J.; et al. Aquaporin-4-dependent glymphatic solute transport in the rodent brain. Elife 2018, 7, e40070. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Mi, X.; Zhan, R.; Li, M.; Wei, L.; Sun, J. Aquaporin 4 Silencing Aggravates Hydrocephalus Induced by Injection of Autologous Blood in Rats. Med. Sci. Monit. 2018, 24, 4204–4212. [Google Scholar] [CrossRef]
- Mortensen, K.N.; Sanggaard, S.; Mestre, H.; Lee, H.; Kostrikov, S.; Xavier, A.L.R.; Gjedde, A.; Benveniste, H.; Nedergaard, M. Impaired Glymphatic Transport in Spontaneously Hypertensive Rats. J. Neurosci. 2019, 39, 6365–6377. [Google Scholar] [CrossRef] [Green Version]
- Tomassoni, D.; Bramanti, V.; Amenta, F. Expression of aquaporins 1 and 4 in the brain of spontaneously hypertensive rats. Brain Res. 2010, 1325, 155–163. [Google Scholar] [CrossRef]
- Naessens, D.M.P.; de Vos, J.; VanBavel, E.; Bakker, E.N.T.P. Blood-brain and blood-cerebrospinal fluid barrier permeability in spontaneously hypertensive rats. Fluids Barriers CNS 2018, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Naessens, D.M.P.; Coolen, B.F.; de Vos, J.; VanBavel, E.; Strijkers, G.J.; Bakker, E.N.T.P. Altered brain fluid management in a rat model of arterial hypertension. Fluids Barriers CNS 2020, 17, 41. [Google Scholar] [CrossRef]
- Oshio, K.; Watanabe, H.; Song, Y.; Verkman, A.S.; Manley, G.T. Reduced cerebrospinal fluid production and intracranial pressure in mice lacking choroid plexus water channel Aquaporin-1. FASEB J. 2005, 19, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Nykanen, M.; Yang, N.; Winlaw, D.; North, K.; Verkman, A.S.; Owler, B.K. Altered cellular localization of aquaporin-1 in experimental hydrocephalus in mice and reduced ventriculomegaly in aquaporin-1 deficiency. Mol. Cell. Neurosci. 2011, 46, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Paul, L.; Madan, M.; Rammling, M.; Chigurupati, S.; Chan, S.L.; Pattisapu, J.V. Expression of aquaporin 1 and 4 in a congenital hydrocephalus rat model. Neurosurgery 2011, 68, 462–473. [Google Scholar] [CrossRef]
- Castañeyra-Ruiz, L.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Castañeyra-Perdomo, A.; González-Marrero, I. AQP1 Overexpression in the CSF of Obstructive Hydrocephalus and Inversion of Its Polarity in the Choroid Plexus of a Chiari Malformation Type II Case. J. Neuropathol. Exp. Neurol. 2019, 78, 641–647. [Google Scholar] [CrossRef]
- Johanson, C.; Stopa, E.; McMillan, P.; Roth, D.; Funk, J.; Krinke, G. The distributional nexus of choroid plexus to cerebrospinal fluid, ependyma and brain: Toxicologic/pathologic phenomena, periventricular destabilization, and lesion spread. Toxicol. Pathol. 2011, 39, 186–212. [Google Scholar] [CrossRef]
- Amiry-Moghaddam, M.; Ottersen, O.P. The molecular basis of water transport in the brain. Nat. Rev. Neurosci. 2003, 4, 991–1001. [Google Scholar] [CrossRef]
- Binder, D.K.; Oshio, K.; Ma, T.; Verkman, A.S.; Manley, G.T. Increased seizure threshold in mice lacking aquaporin-4 water channels. Neuroreport 2004, 15, 259–262. [Google Scholar] [CrossRef]
- Manley, G.T.; Fujimura, M.; Ma, T.; Noshita, N.; Filiz, F.; Bollen, A.W.; Chan, P.; Verkman, A.S. Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat. Med. 2000, 6, 159–163. [Google Scholar] [CrossRef]
- Filippidis, A.S.; Kalani, M.Y.; Rekate, H.L. Hydrocephalus and aquaporins: The role of aquaporin-4. Acta Neurochir. Suppl. 2012, 113, 55–58. [Google Scholar]
- Mao, X.; Enno, T.L.; Del Bigio, M.R. Aquaporin 4 changes in rat brain with severe hydrocephalus. Eur. J. Neurosci. 2006, 23, 2929–2936. [Google Scholar] [CrossRef]
- Tourdias, T.; Dragonu, I.; Fushimi, Y.; Deloire, M.S.; Boiziau, C.; Brochet, B.; Moonen, C.; Petry, K.G.; Dousset, V. Aquaporin 4 correlates with apparent diffusion coefficient and hydrocephalus severity in the rat brain: A combined MRI-histological study. Neuroimage 2009, 47, 659–666. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Manley, G.T.; Krishna, S.; Verkman, A.S. Aquaporin-4 facilitates reabsorption of excess fluid in vasogenic brain edema. FASEB J. 2004, 18, 1291–1293. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Qiao, M.; Lin, L.; Del Bigio, M.R.; Tomanek, B.; Tuor, U.I. Correspondence of AQP4 expression and hypoxic-ischaemic brain oedema monitored by magnetic resonance imaging in the immature and juvenile rat. Eur. J. Neurosci. 2004, 19, 2261–2269. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 2012, 4. [Google Scholar] [CrossRef] [Green Version]
- Iliff, J.; Simon, M. CrossTalk proposal: The glymphatic system supports convective exchange of cerebrospinal fluid and brain interstitial fluid that is mediated by perivascular aquaporin-4. J. Physiol. 2019, 597, 4417–4419. [Google Scholar] [CrossRef]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Pardo, M.R.; Baz-Davila, R.; Lee, S.; Muhonen, M.; Borges, R.; Castañeyra-Perdomo, A. AQP4 labels a subpopulation of white matter-dependent glial radial cells affected by pediatric hydrocephalus, and its expression increased in glial microvesicles released to the cerebrospinal fluid in obstructive hydrocephalus. Acta Neuropathol. Commun. 2022, 10, 41. [Google Scholar] [CrossRef]
- Clarke-Bland, C.E.; Bill, R.M.; Devitt, A. Emerging roles for AQP in mammalian extracellular vesicles. Biochim Biophys Acta Biomembr 2022, 1864, 183826. [Google Scholar] [CrossRef]
- Blocher, J.; Eckert, I.; Elster, J.; Wiefek, J.; Eiffert, H.; Schmidt, H. Aquaporins AQP1 and AQP4 in the cerebrospinal fluid of bacterial meningitis patients. Neurosci. Lett. 2011, 504, 23–27. [Google Scholar] [CrossRef]
- Schmidt, M.J.; Rummel, C.; Hauer, J.; Kolecka, M.; Ondreka, N.; McClure, V.; Roth, J. Increased CSF aquaporin-4, and interleukin-6 levels in dogs with idiopathic communicating internal hydrocephalus and a decrease after ventriculo-peritoneal shunting. Fluids Barriers CNS 2016, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.L.; Kassem, N.A.; Redzic, Z.B.; Chen, C.P.; Segal, M.B.; Preston, J.E. Age-related changes in choroid plexus and blood-cerebrospinal fluid barrier function in the sheep. Exp. Gerontol. 2009, 44, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Goetzl, E.J.; Peltz, C.B.; Mustapic, M.; Kapogiannis, D.; Yaffe, K. Neuron-Derived Plasma Exosome Proteins after Remote Traumatic Brain Injury. J. Neurotrauma 2020, 37, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Beard, K.; Meaney, D.F.; Issadore, D. Clinical Applications of Extracellular Vesicles in the Diagnosis and Treatment of Traumatic Brain Injury. J. Neurotrauma 2020, 37, 2045–2056. [Google Scholar] [CrossRef] [PubMed]
- Bejerot, S.; Hesselmark, E.; Mobarrez, F.; Wallén, H.; Hietala, M.A.; Nybom, R.; Wetterberg, L. Neuromyelitis optica spectrum disorder with increased aquaporin-4 microparticles prior to autoantibodies in cerebrospinal fluid: A case report. J. Med. Case Rep. 2019, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- González-Molina, L.A.; Villar-Vesga, J.; Henao-Restrepo, J.; Villegas, A.; Lopera, F.; Cardona-Gómez, G.P.; Posada-Duque, R. Extracellular Vesicles From 3xTg-AD Mouse and Alzheimer’s Disease Patient Astrocytes Impair Neuroglial and Vascular Components. Front. Aging Neurosci. 2021, 13, 593927. [Google Scholar] [CrossRef] [PubMed]
- Takata, K. Aquaporin-2 (AQP2): Its intracellular compartment and trafficking. Cell. Mol. Biol. 2006, 52, 34–39. [Google Scholar]
- Takata, K.; Matsuzaki, T.; Tajika, Y.; Ablimit, A.; Hasegawa, T. Localization and trafficking of aquaporin 2 in the kidney. Histochem. Cell Biol. 2008, 130, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Conner, M.T.; Conner, A.C.; Bland, C.E.; Taylor, L.H.; Brown, J.E.; Parri, H.R.; Bill, R.M. Rapid aquaporin translocation regulates cellular water flow: Mechanism of hypotonicity-induced subcellular localization of aquaporin 1 water channel. J. Biol. Chem. 2012, 287, 11516–11525. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, P.; Salman, M.M.; Halsey, A.M.; Clarke-Bland, C.; MacDonald, J.A.; Ishida, H.; Vogel, H.J.; Almutiri, S.; Logan, A.; Kreida, S.; et al. Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020, 181, 784–799.e19. [Google Scholar] [CrossRef] [PubMed]
- González-Marrero, I.; Giménez-Llort, L.; Johanson, C.E.; Carmona-Calero, E.M.; Castañeyra-Ruiz, L.; Brito-Armas, J.M.; Castañeyra-Perdomo, A.; Castro-Fuentes, R. Choroid plexus dysfunction impairs beta-amyloid clearance in a triple transgenic mouse model of Alzheimer’s disease. Front. Cell. Neurosci. 2015, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Marrero, I.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Castañeyra-Ruiz, L.; Abreu-Reyes, J.A.; Castañeyra-Perdomo, A. Systemic Hypertension Effects on the Ciliary Body, and Iris. An Immunofluorescence Study with Aquaporin 1, Aquaporin 4, and Na⁺, K⁺ ATPase in Hypertensive Rats. Cells 2018, 7, 210. [Google Scholar] [CrossRef] [Green Version]
- Castañeyra-Ruiz, L.; González-Marrero, I.; González-Toledo, J.M.; Castañeyra-Ruiz, A.; de Paz-Carmona, H.; Castañeyra-Perdomo, A.; Carmona-Calero, E.M. Aquaporin-4 expression in the cerebrospinal fluid in congenital human hydrocephalus. Fluids Barriers CNS 2013, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Carmona-Calero, E.M.; Abreu-Gonzalez, P.; Lecuona, M.; Brage, L.; Rodríguez, E.M.; Castañeyra-Perdomo, A. Cerebrospinal fluid levels of tumor necrosis factor alpha and aquaporin 1 in patients with mild cognitive impairment and idiopathic normal pressure hydrocephalus. Clin. Neurol. Neurosurg. 2016, 146, 76–81. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (ng/mL) | Group | WKY | SHR |
|---|---|---|---|
| AQP1 in CSF | 6 months | 1.95 ± 0.13 | 1.64 ± 0.24 |
| (mean ± SD) | 12 months | 2.01 ± 0.10 | 3.14 ± 0.80 |
| AQP4 in CSF | 6 months | 8.59 ± 0.59 | 9.78 ± 0.82 |
| (mean ± SD) | 12 months | 11.04 ± 0.39 | 18.68 ± 1.33 |
| Group | WKY | SHR | |
|---|---|---|---|
| Body weight | 6 months | 386 ± 2.1 | 352 ± 1.9 * |
| (g. ± SD) | 12 months | 439 ± 2.7 | 398 ± 2.4 * |
| Systolic blood pressure | 6 months | 131 ± 2.9 | 179 ± 1.7 * |
| (mmHg ± SD) | 12 months | 125 ± 1.6 | 185 ± 0.8 * |
| Diastolic blood pressure | 6 months | 58 ± 1.7 | 64 ± 2.7 * |
| (mmHg ± SD) | 12 months | 68 ± 3.0 | 80 ± 2.1 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Marrero, I.; Hernández-Abad, L.G.; González-Gómez, M.; Soto-Viera, M.; Carmona-Calero, E.M.; Castañeyra-Ruiz, L.; Castañeyra-Perdomo, A. Altered Expression of AQP1 and AQP4 in Brain Barriers and Cerebrospinal Fluid May Affect Cerebral Water Balance during Chronic Hypertension. Int. J. Mol. Sci. 2022, 23, 12277. https://doi.org/10.3390/ijms232012277
González-Marrero I, Hernández-Abad LG, González-Gómez M, Soto-Viera M, Carmona-Calero EM, Castañeyra-Ruiz L, Castañeyra-Perdomo A. Altered Expression of AQP1 and AQP4 in Brain Barriers and Cerebrospinal Fluid May Affect Cerebral Water Balance during Chronic Hypertension. International Journal of Molecular Sciences. 2022; 23(20):12277. https://doi.org/10.3390/ijms232012277
Chicago/Turabian StyleGonzález-Marrero, Ibrahim, Luis G. Hernández-Abad, Miriam González-Gómez, María Soto-Viera, Emilia M. Carmona-Calero, Leandro Castañeyra-Ruiz, and Agustín Castañeyra-Perdomo. 2022. "Altered Expression of AQP1 and AQP4 in Brain Barriers and Cerebrospinal Fluid May Affect Cerebral Water Balance during Chronic Hypertension" International Journal of Molecular Sciences 23, no. 20: 12277. https://doi.org/10.3390/ijms232012277
APA StyleGonzález-Marrero, I., Hernández-Abad, L. G., González-Gómez, M., Soto-Viera, M., Carmona-Calero, E. M., Castañeyra-Ruiz, L., & Castañeyra-Perdomo, A. (2022). Altered Expression of AQP1 and AQP4 in Brain Barriers and Cerebrospinal Fluid May Affect Cerebral Water Balance during Chronic Hypertension. International Journal of Molecular Sciences, 23(20), 12277. https://doi.org/10.3390/ijms232012277