Abstract
Nelumbo nucifera Gaertn. is an important perennial aquatic herb that has high ornamental, edible, medicinal, and economic value, being widely distributed and used in China. The NAC superfamily (NAM, ATAF1/2, CUC2) plays critical roles in plant growth, development, and response to abiotic and biotic stresses. Though there have been a few reports about NAC genes in lotus, systematic analysis is still relatively lacking. The present study aimed to characterize all the NAC genes in the lotus and obtain better insights on the NnNACs in response to salt stress by depending on ABA signaling. Here, 97 NAC genes were identified by searching the whole lotus genome based on the raw HMM models of the conserved NAM domain and NAC domain. They were characterized by bioinformatics analysis and divided into 18 subgroups based on the phylogenetic tree. Cis-element analysis demonstrated that NAC genes are responsive to biotic and abiotic stresses, light, low temperature, and plant hormones. Meanwhile, NAC genes had tissue expression specificity. qRT-PCR analysis indicated that NAC genes could be upregulated or downregulated by NaCl treatment, ABA, and fluoridone. In addition, NAC016, NAC025, and NAC070, whose encoding genes were significantly induced by NaCl and ABA, were located in the nucleus. Further analysis showed the three NAC proteins had transcriptional activation capabilities. The co-expression network analysis reflected that NAC proteins may form complexes with other proteins to play a role together. Our study provides a theoretical basis for further research to be conducted on the regulatory mechanisms of salinity resistance in the lotus.
1. Introduction
Plant transcription factors (TFs) play crucial roles in plant growth, development, and abiotic and biotic responses [1]. Considering the importance of TFs in responding to various stimuli, analyzing their function mechanism is becoming indispensable for vegetable crop breeding. With more plant genomes sequenced and transgenic technology developed, a large number of TFs have been identified, and their functions have been determined. NAC (non-apical meristem (NAM), Arabidopsis transcription activation factor (ATAF), and calathiform cotyledon (CUC)) transcription factors characterized by the presence of a highly conserved N-terminal NAC domain and diverse C-terminal domains have been widely studied [2,3]. The N-terminal NAC domain consists of about 160 amino acids that could bind to the cis-elements of their target genes, and C-terminal domains are a fundamental area to interact with other proteins and regulate other gene expression [4,5]. The structure of NAC proteins being well-identified provided a framework for further understanding the functions of NAC proteins at the transcriptional level. Subsequently, research on the functions of NAC transcription factors mushroomed in a variety of species.
Gene family analysis as a basic method plays an important role in investigating the functions of plant transcription factors. The members of the NAC family have been identified and reported in many species by gene family analysis. For instance, 108 members have been identified in Arabidopsis, 151 members in rice, 150 members in soybean, 261 members in M. sinensis, 132 members in peanut, 151 members in sunflower, 87 members in maize, 111 members in celery, 104 members in pepper, and 110 members in potato [6,7,8,9,10,11,12]. NAC proteins have not only been identified, but their functions have also been well studied. NAC TFs play important roles in many vital biological processes and plant stress responses [13,14,15,16,17,18].
Considering that abiotic stresses could adversely affect crop growth and productivity, it is very important to study the functions of NAC TFs in improving crop resistance to abiotic stresses, which have been well elucidated in many species in last decade. In grain crop rice, ONAC066, ONAC095, OsNAC2, OsNAC006, OsNAC67, OsNAC016, and OsNAP function as positive or negative regulators of the drought stress response [15,19,20,21,22,23,24]. In vegetable crops, NAC TF genes, such as JUB1, SlVOZ1, SlNAC35, StNAC053, and CaNAC46, also play an important role in plant responses to drought stresses [25,26,27,28,29]. In addition, NAC TFs have been widely focused on regarding plant resistance to salinity, which reflects the fact that one NAC TF can have multiple functions. Potato StNAC053, grapevine VvNAC17, pepper CaNAC46, rice ONAC022, and wheat TaNAC29 enhance not only plant drought tolerance but also salt tolerance [25,26,30,31,32]. These research works provide us with a clear research direction and a very important basis from which to elucidate the regulation mechanism of NAC TFs.
NAC TFs can regulate several plant signal networks and be regulated by environmental stimuli, plant hormones, and miRNAs. Environmental stimuli, such as light, temperature, drought, salinity, water, etc., could induce the expression of NAC TFs [33,34,35]. Furthermore, plant hormones are the essential regulators of plant growth and development. The interplay between plant hormones and NAC TFs has become an interesting research topic. Wu et al. discussed interaction between NAC TFs and ethylene and abscisic acid (ABA) pathways in controlling tomato fruit development and ripening [36]. OsNAC3, OsNAC45, and VvNAC17 improved transgenic plant ABA sensitivity and salt tolerance [31,37,38]. Further study demonstrated that CrNAC036 can directly bind and negatively regulate CrNCED5 expression to hinder ABA biosynthesis in citrus [39]. Interestingly, recent studies showed microRNAs (miRNAs) comprising about 22 nt endogenous noncoding RNAs could take part in plant abiotic responses by targeting NAC genes. In other words, NAC genes were regulated by plant miRNAs. Peu-miR164 regulated its target genes PeNAC070 and PeNAC081, which are involved in the poplar response to abiotic stress [40]. osa-miR164c negatively regulated OsCUC1 and OsCUC3, which are involved in rice meristem/organ boundary specification [41]. In addition, miR1514a modulates NAC700 transcription to trigger phasiRNA formation in response to water deficit in legumes [42]. These reports indicate that the NAC TF-related networks are complex and important for plant growth and development.
Lotus (Nelumbo nucifera Gaertn.), popular ornamental, edible, and medicinal plant which belongs to the small family Nelumbonaceae, will be an important model plant in horticulture [43]. With three lotus genomes sequenced, the lotus TFs, such as AP2/ERF, WRKY, bHLH, and the MADS gene family, have been widely studied [44,45,46,47,48,49,50]. However, the NAC gene family has not yet been comprehensively studied in lotus. Though 82 NnNAC genes have been found using the PlantTFDB 4.0, the NAC gene family still needs to be identified by searching the whole lotus genome. Here, all NAC members of lotus were identified, and their distribution, phylogenetic relationship, gene structure, and synteny were analyzed. In addition, certain candidate NAC TFs which had a higher expression in lotus leaves were further studied though subcellular localization and expression level analysis by qRT-PCR. These results will further deepen the understanding of NAC TFs and provide several candidate genes related to salinity and abscisic acid (ABA) response in lotus.
2. Results
2.1. Identification of NAC Transcription Factors in Nelumbo nucifera
To identify all NAC TFs in Nelumbo nucifera, the conserved NAM domain (PF02365) and NAC domain (PF01849) were used to seek the respective raw HMM models. Based on these two HMM models, 107 and 106 candidate members were identified, and the redundant forms of the same genes were removed. In total, 97 NAC TF members were identified and named NAC001–NAC097 according to their chromosome location (Table 1 and Figure 1). The basic information, including genomic length, amino acid residues, pI, and molecular weight, was calculated. For example, the polypeptide lengths of the predicted NACs varied widely, ranging from 76 to 684. In addition, the pI data showed that NACs have both acidic and alkaline members. The information also contained the members’ gene IDs, which could help researchers obtain recent news on NACs (Table 1).

Table 1.
The NAC genes and properties of the deduced proteins in lotus root.

Figure 1.
Chromosomal locations of lotus NAC genes. The brown sticks represent the chromosomes, and the chromosome numbers are shown on top of the sticks. The lengths of the chromosomes are on the left of the figure.
2.2. Phylogenetic Analysis and Classification of NAC Genes
Eighteen NAC subgroups have been classified in rice and Arabidopsis [51]. In our results, 18 new subgroups were redefined in which no lotus NAC members belong to ONAC001 and OsNAC3. However, the lotus NAC members family had two specific subgroups named NNACO23 and NNAC014, which indicated lotus is species-specific compared to Arabidopsis and rice (Figure 2).
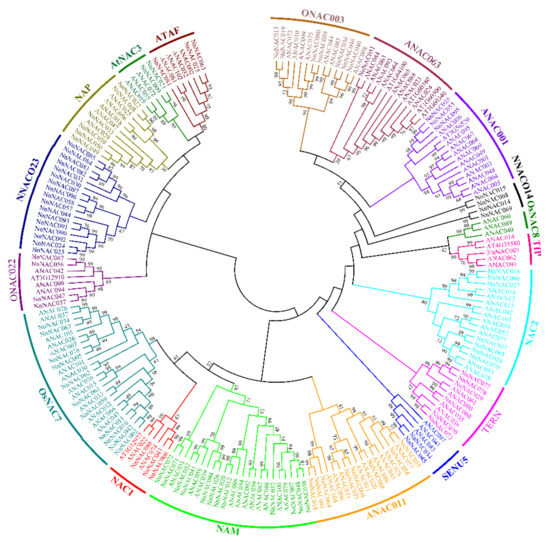
Figure 2.
Neighbor-joining phylogenetic tree of the NAC family in lotus and Arabidopsis.
2.3. Gene Structure and Motif Composition of the NAC Gene Family
The sequence features including introns, exons, and motifs were visualized by the TBtools software [52]. According to the frequency of occurrence, motifs 1–5 were the five most frequently presented motifs, covering almost all NAC members. Furthermore, motifs 8–10 only existed in the members NnNAC082, 083, 084, 085, and 031, which belong to subgroup NNAC023. Interestingly, NnNAC009 and NnNAC017 possessed two copies of motif 3. In addition, the motifs in subgroup ONAC003 showed a character of random arrangement. The exhibition of exon–intron structure showed that there is evolutionary diversity. Most of the members had more than three exons, and only four members had one exon. NnNAC069, 27, 28, and 79 possessed relatively longer intron lengths (Figure 3).

Figure 3.
The structures of NAC genes and NAC proteins. The left represents the phylogenetic relationship of NAC genes. The middle represents the conserved motifs in NAC proteins, in which different colors represent different motifs. The right represents the intron–exon structures of NAC genes. The exons, introns, and untranslated regions (UTRs), are indicated by the green boxes, black lines, and yellow boxes, respectively.
2.4. Cis-Acting Elements and Synteny Analysis of NAC Gene Promotors
To investigate the functions of NAC genes, the cis-acting elements of NAC gene promotors were analyzed by the online tool PlantCARE; the data were then visualized by TBtools. All NAC genes had the “light responsiveness” element, suggesting that the “light responsiveness” element is fundamental for plant growth and development. In addition, plant hormones were the important factors for plant growth, development, and response to the environment. Furthermore, the plant hormone elements “abscisic acid responsiveness”, “gibberellin responsiveness”, “MeJA responsiveness”, “salicylic acid responsiveness”, and “auxin responsiveness” were widely presented in NAC gene promotors (Figure 4). The “defense and stress responsiveness” element, which weas also widely presented in NAC gene promotors, played important roles in plant responses to abiotic and biotic stresses. The “low-temperature responsiveness” element was also important because lotus root is occasionally affected by low temperatures. The above elements suggest that NAC genes in lotus root may function in diverse aspects of lotus root growth and development. To compare the evolutionary events between lotus and Arabidopsis, synteny analysis was carried out using the orthologous NAC gene pairs. A total of 69 collinear genes were retrieved from the two species (Figure S1).

Figure 4.
The cis-acting elements in the promoters of soybean SRS genes.
2.5. Expression Patterns of NAC Genes in Different Plant Tissues
To investigate the tissue-specific expression of NAC genes, we obtained data from the “Nelumbo Genome Database” which contained the expression levels of NAC genes in the petiole, leaf, root, and cotyledon (Table S2). The results showed that NAC genes had tissue expression specificity, indicating that their functions were diverse. For example, NAC006 was only expressed in the leaf, root, and petiole. Unlike NAC006, three genes, NAC020, 012, and 071, were only expressed in the cotyledon. The leaf was the most important organ, and its phenotypes were easily observed after stress treatment. We selected the 10 most highly expressed genes—NAC003, 016, 025, 034, 043, 045, 060, 065, 070, and 078—for further research.
2.6. qRT-PCR Analysis of the NAC Genes
To investigate the functions of the NAC genes, the above selected genes were analyzed by qRT-PCR under NaCl, ABA, and ABA + FL treatment (Figure 5). Under NaCl treatment, the transcription levels of nine NAC genes increased gradually, with NnNAC078 being the sole exception. Under ABA treatment, the 10 NAC genes were all induced, and the transcription level of NnNAC003 fluctuated. Fluoridone, the inhibitor of ABA, also induced gene transcription. In most cases, FL affects the action of ABA. In addition, the interaction of ABA and FL had positive effects on the expression levels of NAC genes including NnNAC003, NnNAC025, NnNAC045, and NnNAC065. The expression levels of NnNAC16, NnNAC25, and NnNAC70 were significantly increased under ABA and NaCl treatment, while their expression levels were significantly reduced under ABA and FL treatment; they were selected for further research.
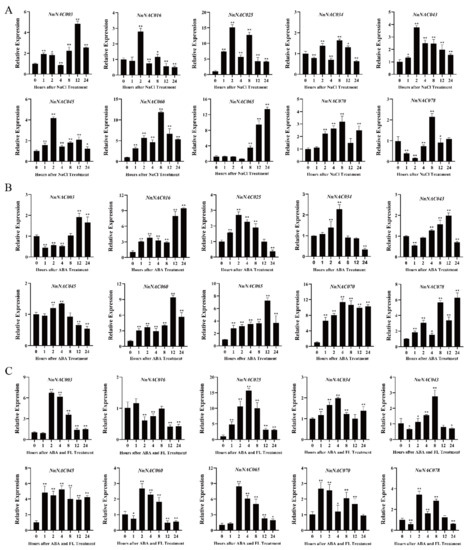
Figure 5.
Relative expression levels of lotus NAC genes. qRT-PCR analyses of plants treated with NaCl (A), ABA (B), and ABA + FL (C). Independent t-tests demonstrated that there was significant difference (* p <0.05, ** p < 0.01).
2.7. Subcellular Localization of NAC Proteins
Subcellular localization could indicate the role and position of NAC-encoded proteins. In order to understand the molecular characteristics of hormone- and stress-responsive lotus root NACs, we selected three lotus root NAC genes for subcellular localization. Green fluorescence of the NnNAC16, NnNAC25, and NnNAC70 proteins all appeared in the nucleus, while green fluorescence of the control protein appeared on the cell membrane, in the cytoplasm, and in the nucleus, which indicated that the NnNAC16, NnNAC25, and NnNAC70 proteins function in the nucleus (Figure 6).
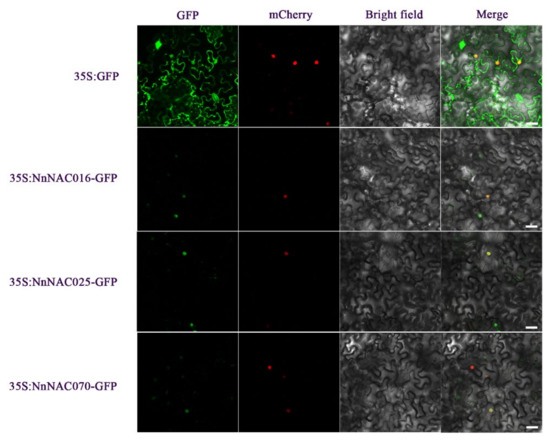
Figure 6.
Subcellular localization of the soybean NAC proteins. The first row of pictures above is a control, and those below it are the NnNAC016, NnNAC025, and NnNAC070 proteins. Scale bars = 25 μm.
2.8. The Transcriptional Activation Capabilities Analysis of NAC Proteins
Transcription factors generally have self-activation ability. The full-length cDNAs of NnNAC16, NnNAC25, and NnNAC70 were cloned into a PGBKT7 vector, and the self-activation ability of NnNAC16, NnNAC25, and NnNAC70 was verified by yeast experimentation. The results showed that NnNAC16, NnNAC25, NnNAC70, and the negative control pGBKT7 could all grow colonies on an SD/-Trp medium, while only the negative control pGBKT7 failed to grow colonies on an SD/-Trp/-Ade/His medium. This indicated that NnNAC16, NnNAC25, and NnNAC70 all had self-activation ability (Figure 7).
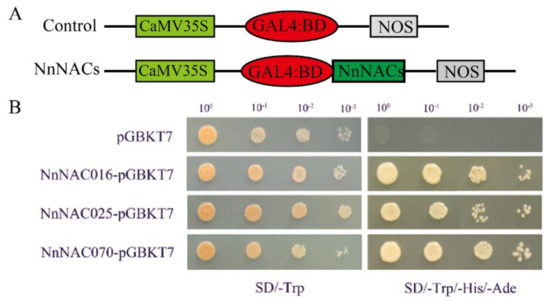
Figure 7.
Transcription activation analysis of the three selected ANC proteins. (A) The diagram shows the structure of the fusion plasmid. (B) Transcription activation analysis of the nine selected NAC proteins. pGBKT7 was used as a negative control.
To further investigate the functions of the three NnNAC genes, their homologs in Arabidopsis were identified. Then, their co-expression networks were drawn using the website STRING. The results indicated that the functions of NAC genes were diverse and closely connected with NaCl, drought, and the plant hormone ABA (Figure S2). Co-expression networks allowed us to better study NAC genes in lotus root.
3. Discussion
With the completion of the genome sequencing of lotus root, several gene families were identified [44,47,53]. NAC proteins have diverse functions and play vital roles in plant growth and development, including responses to biotic and abiotic stresses [14,17,54,55,56,57,58,59,60]. However, functional research of lotus NAC genes has rarely been reported. Eighty-two NAC genes were obtained using the website PlantTFDB, and we studied their potential functions in lotus seeds and response to complete submergence [61]. Compared to NAC genes of other plants, the number NAC genes in lotus root still needs to be found. Here, we identified 97 NAC gene members by searching the whole lotus genome based on the raw HMM models for the conserved NAM domain (PF02365) and NAC domain (PF01849), which could look for as many genes as possible. Then, the 97 members were divided into 18 subgroups based on the classification method used in other plants [62,63]. The promotor, as an important structure, plays vital roles in plant growth and development. All NAC genes had the “light responsiveness” element, which is the fundamental element for plant growth and development. ABA is an important phytohormone regulating plant growth, development, and stress responses [64]. The “abscisic acid responsiveness” element in the promotor of NAC genes indicated that NAC genes may be regulated by the ABA-related transcription factors. Meanwhile, the qRT-PCR analysis also showed that NAC genes were induced under ABA treatment (Figure 4). In addition, the co-expression network of Arabidopsis homologs of three NnNAC genes also indicated the functions of NAC genes and ABA signaling are closely connected (Figure 7). For example, PwNAC11 activated ERD1 by interaction with ABA-related proteins, ABF3 and DREB2A, to enhance drought tolerance in transgenic Arabidopsis [65]. OsNAC45 was involved in ABA response and salt tolerance in rice [38]. In addition, “gibberellin responsiveness”, “MeJA responsiveness”, “salicylic acid responsiveness”, and “auxin responsiveness” were also widely presented in NAC gene promotors, which indicates NAC genes are multifunctional, interacting with many plant hormones’ signaling. These could provide us with a new avenue by which to study the function of NAC genes. Previous research showed that MeJA enhanced ethylene synthesis by inducing NAC genes in kiwifruit [66]. In addition, NAC genes could function in response to biotic stresses based on the “defense and stress responsiveness” element in their promotors. In total, the functions of NAC genes still need to be further studied.
The lotus is an important ornamental, edible, and medicinal plant originating from India and China. It is famous for its products such as fresh rhizomes, lotus root starch, drinks, teas, and lotus seeds [67]. However, the plant’s yield and product quality are affected by the salinity of soil [68]. Therefore, to excavate the key resistance genes is an important work for breeding superior lotus varieties. Furthermore, our results showed that NAC genes could be candidate genes to improve lotus resistance to slat stress. For instance, NnNAC003, NnNAC016, NnNAC043, NnNAC045, NnNAC060, and NnNAC070 were significantly induced under NaCl treatment (Figure 4). Further results indicated that their functions in response to salt stress were dependent on ABA signaling (Figure 3 and Figure 4). In order to understand where NAC proteins function, we conducted a subcellular localization assay which demonstrated that NAC proteins function in the nucleus, as do most transcription factors (Figure 5). Like most transcription factors, the three NAC proteins, NnNAC16, NnNAC25, and NnNAC70, have transcriptional activation, which could bind them to the related cis-elements to regulate the expression levels of target genes. In addition, NAC proteins may form complexes with other proteins to play a role together (Figure 7).
In total, the 97 NAC genes may be involved in responses to biotic and abiotic stresses by depending on ABA or other plant hormone signaling and would be the key candidate genes for further functional research by modern genetic and biological techniques.
4. Materials and Methods
4.1. The Identification of NAC Transcription Factor Members
The Nelumbo nucifera Geartn. genome (China Antique v2.0) was obtained from the Nelumbo Genome Database [48]. The raw HMM models for the conserved NAM domain (PF02365) and NAC domain (PF01849) were downloaded from the Pfam database (https://pfam.xfam.org/ (accessed on 27 May 2022)). the raw HMM models were then used to search the Nelumbo nucifera Geartn. genome with the software TBtools [52] to obtain the candidate NAC transcription factor members. Then, certain NAC transcription factor members were determined by their sequence search in the Pfam database with the threshold of an e-value <e−5. Next, to exclude repeated sequences, all NAC sequences were aligned using DANMAN Version 7 and checked manually.
4.2. Basic Information and Phylogenetic Analyses
The NAC genes were visualized on the chromosomes using the software TBtools [52]. Their CDS length, molecular weight, and isoelectric point were predicted by the ExPASy tool (https://www.expasy.org/ (accessed on 5 June 2022)). The amino acid sequences of Arabidopsis and Nelumbo nucifera Geartn. were download from TAIR (https://www.arabidopsis.org/ (accessed on 5 June 2022)) and the Nelumbo Genome Database [48], respectively. Based on their amino acid sequences, the phylogenetic tree was constructed in MEGA7.0 by the neighbor-joining (NJ) algorithm; 1,000 replicates were used to evaluate the significance of nodes [69]. The annotation on the phylogenetic tree was created in the Adobe Illustrator SC5 software.
4.3. Synteny and Gene Structure Analyses
The applet MCScanX (default parameters) in TBtools was used to examine duplicate genes. Then, the homology of the NAC gene between Nelumbo nucifera Geartn. and Arabidopsis was visualized using the applet Dual Synteny Plotter in TBtools [52]. The conserved motifs of NAC proteins were identified using the online tool MEME Suite. The cis-element of the NAC gene promotor was analyzed via the website PlantCARE [70]. The motifs and cis-element were exhibited in one figure by TBtools.
4.4. Microarray Analysis of NAC Gene Expression Patterns
The tissue expression data of NAC genes was downloaded from the Nelumbo Genome Database. The analysis was carried out, which included the 97 NAC genes in the different tissues and development stages, and then it was visualized by TBtools.
4.5. Subcellular Localization Analysis
The selected proteins were fused with the expression vector pCAMBIA1300, containing a GFP tag for subcellular localization analysis, as described previously [71]. A diluted agrobacterium solution (NAC-pCAMBIA1300 or pCAMBIA1300) was injected into the leaves of tobacco. After 24 h of incubation, fluorescence images were captured by a confocal laser scanning microscope (TCS SP8; Leica). The primers used in this study can be found in Table S1.
4.6. Plant Materials and Treatments
Lotus seeds were germinated in soil in a greenhouse. Two-week-old seedlings were subjected to exogenous ABA and salinity treatments. For these treatments, seedlings were transferred to solutions containing 200 mM NaCl or 100 μM ABA. Seedlings were sampled at 0, 1, 2, 4, 8, 12, and 24 h after treatment. The samples were then dropped immediately into liquid nitrogen and stored at −80 °C.
4.7. Quantitative Real-Time PCR (qRT-PCR)
The samples were ground in liquid nitrogen, and the powder was used to extract the RNA. The cDNA synthesis was conducted following the manufacturer’s instructions using a PrimeScript™ 1st Strand cDNA Synthesis Kit (TaKaRa, Beijing China). Then, the expression patterns were analyzed with a BIO-RAD CFX MaestroTM system. In this assay, three biological and technical replicates were made. The histogram was made with the GraphPad Prism 8.0 software, and the standard error was calculated with the SPSS software. The primers used in this study can be found in Table S1.
4.8. The Transcriptional Activation Assays
The transactivation ability of NAC TFs was tested in yeast cells. The fusion plasmids NnNACs:pGBKT7 and the control plasmid pGBKT7 were transformed into Y2H Gold yeast cells. Transformed yeast competent cells were spread on plates containing SD/-Trp or SD/-Trp/-His/-Ade.
Supplementary Materials
The supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232012394/s1.
Author Contributions
Conceptualization, S.Z. and L.L.; data curation, S.Z., P.W. and K.F.; funding acquisition, S.Z. and L.L.; investigation, T.J., Y.Z. and K.Z.; methodology, S.Z.; project administration, S.Z.; software, S.Z. and T.J.; supervision, L.L.; visualization, S.Z.; writing—original draft, S.Z.; writing—review and editing, L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32202472), Jiangsu Seed Industry Revitalization ‘Jie Bang Gua Shuai’ project (JBGS [2021]017), the National Key R&D Program of China (2020YFD1000300), and the China Agriculture Research System (CARS-24).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, W.; Stewart, C.N. Plant synthetic promoters and transcription factors. Curr. Opin. Biotechnol. 2016, 37, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Munir, N.; Yukun, C.; Xiaohui, C.; Nawaz, M.A.; Iftikhar, J.; Rizwan, H.M.; Xu, S.; Yuling, L.; Xuhan, X.; Zhongxiong, L. Genome-wide identification and comprehensive analyses of NAC transcription factor gene family and expression patterns during somatic embryogenesis in Dimocarpus longan Lour. Plant Physiol. Biochem. 2020, 157, 169–184. [Google Scholar] [CrossRef]
- Ernst, H.A.; Olsen, A.N.; Larsen, S.; Lo Leggio, L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Kjaersgaard, T.; Jensen, M.K.; Christiansen, M.W.; Gregersen, P.; Kragelund, B.B.; Skriver, K. Senescence-associated barley NAC (NAM, ATAF1,2, CUC) transcription factor interacts with radical-induced cell death 1 through a disordered regulatory domain. J. Biol. Chem. 2011, 286, 35418–35429. [Google Scholar] [CrossRef] [PubMed]
- Bengoa Luoni, S.A.; Cenci, A.; Moschen, S.; Nicosia, S.; Radonic, L.M.; Sabio, Y.G.J.V.; Langlade, N.B.; Vile, D.; Rovere, C.V.; Fernandez, P. Genome-wide and comparative phylogenetic analysis of senescence-associated NAC transcription factors in sunflower (Helianthus annuus). BMC Genom. 2021, 22, 893. [Google Scholar] [CrossRef] [PubMed]
- Li, P.X.; Peng, Z.Y.; Xu, P.L.; Tang, G.Y.; Ma, C.L.; Zhu, J.Q.; Shan, L.; Wan, S.B. Genome-Wide Identification of NAC Transcription Factors and Their Functional Prediction of Abiotic Stress Response in Peanut. Front. Genet. 2021, 12, 630292. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Yang, Z.; He, J.; Liu, A.; Chen, J.; Wang, S.; Wang, X.; Feng, G.; Li, D.; Peng, Y.; et al. Genome-Wide Investigation of the NAC Transcription Factor Family in Miscanthus sinensis and Expression Analysis Under Various Abiotic Stress. Front. Plant Sci. 2021, 12, 766550. [Google Scholar] [CrossRef] [PubMed]
- Duan, A.Q.; Yang, X.L.; Feng, K.; Liu, J.X.; Xu, Z.S.; Xiong, A.S. Genome-wide analysis of NAC transcription factors and their response to abiotic stress in celery (Apium graveolens L.). Comput. Biol. Chem. 2020, 84, 107186. [Google Scholar] [CrossRef]
- Diao, W.; Snyder, J.C.; Wang, S.; Liu, J.; Pan, B.; Guo, G.; Ge, W.; Dawood, M. Genome-Wide Analyses of the NAC Transcription Factor Gene Family in Pepper (Capsicum annuum L.): Chromosome Location, Phylogeny, Structure, Expression Patterns, Cis-Elements in the Promoter, and Interaction Network. Int. J. Mol. Sci. 2018, 19, 1028. [Google Scholar] [CrossRef]
- Singh, A.K.; Sharma, V.; Pal, A.K.; Acharya, V.; Ahuja, P.S. Genome-wide organization and expression profiling of the NAC transcription factor family in potato (Solanum tuberosum L.). DNA Res. 2013, 20, 403–423. [Google Scholar] [CrossRef]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res. 2011, 18, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Wu, M.; Zhou, Y.; Gong, Z.; Yu, W.; Zhang, Y.; Yang, Z. NAC-mediated membrane lipid remodeling negatively regulates fruit cold tolerance. Hortic. Res. 2022, 9, uhac039. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Kang, X.; Liu, P.; She, K.; Zhang, Y.; Lin, X.; Li, B.; Chen, Z. The NAC-like transcription factor CsNAC7 positively regulates the caffeine biosynthesis-related gene yhNMT1 in Camellia sinensis. Hortic. Res. 2022, 9, uhab046. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, Y.; Xie, Z.; Yu, B.; Sun, Y.; Huang, J. OsNAC016 regulates plant architecture and drought tolerance by interacting with the kinases GSK2 and SAPK8. Plant Physiol. 2022, 189, 1296–1313. [Google Scholar] [CrossRef]
- Liu, G.S.; Li, H.L.; Grierson, D.; Fu, D.Q. NAC Transcription Factor Family Regulation of Fruit Ripening and Quality: A Review. Cells 2022, 11, 525. [Google Scholar] [CrossRef]
- Xu, Y.; Zou, S.; Zeng, H.; Wang, W.; Wang, B.; Wang, H.; Tang, D. A NAC Transcription Factor TuNAC69 Contributes to ANK-NLR-WRKY NLR-Mediated Stripe Rust Resistance in the Diploid Wheat Triticum urartu. Int. J. Mol. Sci. 2022, 23, 564. [Google Scholar] [CrossRef]
- Wang, J.; Tian, S.; Yu, Y.; Ren, Y.; Guo, S.; Zhang, J.; Li, M.; Zhang, H.; Gong, G.; Wang, M.; et al. Natural variation in the NAC transcription factor NONRIPENING contributes to melon fruit ripening. J. Integr. Plant Biol. 2022, 64, 1448–1461. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell Physiol. 2014, 55, 604–619. [Google Scholar] [CrossRef]
- Rahman, H.; Ramanathan, V.; Nallathambi, J.; Duraialagaraja, S.; Muthurajan, R. Over-expression of a NAC 67 transcription factor from finger millet (Eleusine coracana L.) confers tolerance against salinity and drought stress in rice. BMC Biotechnol. 2016, 16 (Suppl. S1), 35. [Google Scholar] [CrossRef]
- Wang, B.; Zhong, Z.; Wang, X.; Han, X.; Yu, D.; Wang, C.; Song, W.; Zheng, X.; Chen, C.; Zhang, Y. Knockout of the OsNAC006 Transcription Factor Causes Drought and Heat Sensitivity in Rice. Int. J. Mol. Sci. 2020, 21, 2288. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Lv, B.; Luo, L.; He, J.; Mao, C.; Xi, D.; Ming, F. The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 7, 40641. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Hong, Y.; Zhang, H.; Li, D.; Song, F. Rice NAC transcription factor ONAC095 plays opposite roles in drought and cold stress tolerance. BMC Plant Biol. 2016, 16, 203. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wang, H.; Cai, J.; Bi, Y.; Li, D.; Song, F. Rice NAC transcription factor ONAC066 functions as a positive regulator of drought and oxidative stress response. BMC Plant Biol. 2019, 19, 278. [Google Scholar] [CrossRef]
- Ma, J.; Wang, L.Y.; Dai, J.X.; Wang, Y.; Lin, D. The NAC-type transcription factor CaNAC46 regulates the salt and drought tolerance of transgenic Arabidopsis thaliana. BMC Plant Biol. 2021, 21, 11. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, C.; Li, Z.; Sun, J.; Deng, Z.; Wen, L.; Li, X.; Guo, Y. Potato NAC Transcription Factor StNAC053 Enhances Salt and Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 2568. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, S.; Ma, X.; Wang, Y.; Kong, F.; Meng, Q. A stress-associated NAC transcription factor (SlNAC35) from tomato plays a positive role in biotic and abiotic stresses. Physiol. Plant. 2016, 158, 45–64. [Google Scholar] [CrossRef]
- Chong, L.; Xu, R.; Huang, P.; Guo, P.; Zhu, M.; Du, H.; Sun, X.; Ku, L.; Zhu, J.K.; Zhu, Y. The tomato OST1-VOZ1 module regulates drought-mediated flowering. Plant Cell 2022, 34, 2001–2018. [Google Scholar] [CrossRef]
- Thirumalaikumar, V.P.; Devkar, V.; Mehterov, N.; Ali, S.; Ozgur, R.; Turkan, I.; Mueller-Roeber, B.; Balazadeh, S. NAC transcription factor JUNGBRUNNEN1 enhances drought tolerance in tomato. Plant Biotechnol. J. 2018, 16, 354–366. [Google Scholar] [CrossRef]
- Hong, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of a Stress-Responsive NAC Transcription Factor Gene ONAC022 Improves Drought and Salt Tolerance in Rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef]
- Ju, Y.L.; Yue, X.F.; Min, Z.; Wang, X.H.; Fang, Y.L.; Zhang, J.X. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC transcription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 146, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 268. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Rashotte, A.M.; Dane, F. Citrullus colocynthis NAC transcription factors CcNAC1 and CcNAC2 are involved in light and auxin signaling. Plant Cell Rep. 2014, 33, 1673–1686. [Google Scholar] [CrossRef] [PubMed]
- Diao, P.; Chen, C.; Zhang, Y.; Meng, Q.; Lv, W.; Ma, N. The role of NAC transcription factor in plant cold response. Plant Signal. Behav. 2020, 15, 1785668. [Google Scholar] [CrossRef]
- Wu, J.; Wang, L.; Wang, S. Comprehensive analysis and discovery of drought-related NAC transcription factors in common bean. BMC Plant Biol. 2016, 16, 193. [Google Scholar] [CrossRef]
- Kou, X.; Zhou, J.; Wu, C.E.; Yang, S.; Liu, Y.; Chai, L.; Xue, Z. The interplay between ABA/ethylene and NAC TFs in tomato fruit ripening: A review. Plant Mol. Biol. 2021, 106, 223–238. [Google Scholar] [CrossRef]
- Zhang, X.; Long, Y.; Chen, X.; Zhang, B.; Xin, Y.; Li, L.; Cao, S.; Liu, F.; Wang, Z.; Huang, H.; et al. A NAC transcription factor OsNAC3 positively regulates ABA response and salt tolerance in rice. BMC Plant Biol. 2021, 21, 546. [Google Scholar] [CrossRef]
- Zhang, X.; Long, Y.; Huang, J.; Xia, J. OsNAC45 is Involved in ABA Response and Salt Tolerance in Rice. Rice 2020, 13, 79. [Google Scholar] [CrossRef]
- Zhu, F.; Luo, T.; Liu, C.; Wang, Y.; Zheng, L.; Xiao, X.; Zhang, M.; Yang, H.; Yang, W.; Xu, R.; et al. A NAC transcription factor and its interaction protein hinder abscisic acid biosynthesis by synergistically repressing NCED5 in Citrus reticulata. J. Exp. Bot. 2020, 71, 3613–3625. [Google Scholar] [CrossRef]
- Lu, X.; Dun, H.; Lian, C.; Zhang, X.; Yin, W.; Xia, X. The role of peu-miR164 and its target PeNAC genes in response to abiotic stress in Populus euphratica. Plant Physiol. Biochem. 2017, 115, 418–438. [Google Scholar] [CrossRef]
- Wang, J.; Bao, J.; Zhou, B.; Li, M.; Li, X.; Jin, J. The osa-miR164 target OsCUC1 functions redundantly with OsCUC3 in controlling rice meristem/organ boundary specification. New Phytol. 2021, 229, 1566–1581. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Valencia, G.; Palomar, M.; Covarrubias, A.A.; Reyes, J.L. The legume miR1514a modulates a NAC transcription factor transcript to trigger phasiRNA formation in response to drought. J. Exp. Bot. 2017, 68, 2013–2026. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, C.; Cao, D.; Damaris, R.N.; Yang, P. The Latest Studies on Lotus (Nelumbo nucifera)—An Emerging Horticultural Model Plant. Int. J. Mol. Sci. 2019, 20, 3680. [Google Scholar] [CrossRef]
- Cao, D.; Lin, Z.; Huang, L.; Damaris, R.N.; Yang, P. Genome-wide analysis of AP2/ERF superfamily in lotus (Nelumbo nucifera) and the association between NnADAP and rhizome morphology. BMC Genom. 2021, 22, 171. [Google Scholar] [CrossRef]
- Li, J.; Xiong, Y.; Li, Y.; Ye, S.; Yin, Q.; Gao, S.; Yang, D.; Yang, M.; Palva, E.T.; Deng, X. Comprehensive Analysis and Functional Studies of WRKY Transcription Factors in Nelumbo nucifera. Int. J. Mol. Sci. 2019, 20, 5006. [Google Scholar] [CrossRef] [PubMed]
- Mao, T.Y.; Liu, Y.Y.; Zhu, H.H.; Zhang, J.; Yang, J.X.; Fu, Q.; Wang, N.; Wang, Z. Genome-wide analyses of the bHLH gene family reveals structural and functional characteristics in the aquatic plant Nelumbo nucifera. PeerJ 2019, 7, e7153. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Cao, D.; Damaris, R.N.; Yang, P. Genome-wide identification of MADS-box gene family in sacred lotus (Nelumbo nucifera) identifies a SEPALLATA homolog gene involved in floral development. BMC Plant Biol. 2020, 20, 497. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; VanBuren, R.; Liu, Y.; Yang, M.; Han, Y.; Li, L.T.; Zhang, Q.; Kim, M.J.; Schatz, M.C.; Campbell, M.; et al. Genome of the long-living sacred lotus (Nelumbo nucifera Gaertn.). Genome Biol. 2013, 14, R41. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, G.; Liu, Y.; Sun, F.; Shi, C.; Liu, X.; Peng, J.; Chen, W.; Huang, X.; Cheng, S.; et al. The sacred lotus genome provides insights into the evolution of flowering plants. Plant J. 2013, 76, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, T.; Cheng, T.; Zhao, L.; Zheng, X.; Zhu, F.; Dong, C.; Xu, J.; Xie, K.; Hu, Z.; et al. Genomic variation reveals demographic history and biological adaptation of the ancient relictual, lotus (Nelumbo Adans). Hortic. Res. 2022, 9, uhac029. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, S.; Zhou, Y.; Zhou, Y.; Yang, J.; Tang, X. Genome-wide identification and characterization of GRAS transcription factors in sacred lotus (Nelumbo nucifera). PeerJ 2016, 4, e2388. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Hernandez, B.Y.; Nunez-Munoz, L.; Calderon-Perez, B.; Xoconostle-Cazares, B.; Ruiz-Medrano, R. The NAC Transcription Factor ANAC087 Induces Aerial Rosette Development and Leaf Senescence in Arabidopsis. Front. Plant Sci. 2022, 13, 818107. [Google Scholar] [CrossRef]
- Tao, Y.; Wan, J.X.; Liu, Y.S.; Yang, X.Z.; Shen, R.F.; Zhu, X.F. The NAC transcription factor ANAC017 regulates aluminum tolerance by regulating the cell wall-modifying genes. Plant Physiol. 2022, 189, 2517–2534. [Google Scholar] [CrossRef]
- Song, S.; Willems, L.A.J.; Jiao, A.; Zhao, T.; Eric Schranz, M.; Bentsink, L. The membrane associated NAC transcription factors ANAC060 and ANAC040 are functionally redundant in the inhibition of seed dormancy in Arabidopsis thaliana. J. Exp. Bot. 2022, 73, 5514–5528. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Li, S.; Chen, B.; Jian, C.; Mei, F.; Zhang, Y.; Li, F.; Chen, N.; Li, T.; Du, L.; et al. Variation in cis-regulation of a NAC transcription factor contributes to drought tolerance in wheat. Mol. Plant 2022, 15, 276–292. [Google Scholar] [CrossRef]
- Luo, P.; Chen, Y.; Rong, K.; Lu, Y.; Wang, N.; Xu, Z.; Pang, B.; Zhou, D.; Weng, J.; Li, M.; et al. ZmSNAC13, a maize NAC transcription factor conferring enhanced resistance to multiple abiotic stresses in transgenic Arabidopsis. Plant Physiol. Biochem. 2022, 170, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lin, Y.; Xu, M.; Bian, H.; Zhang, C.; Wang, J.; Wang, H.; Xu, Y.; Niu, Q.; Zuo, J.; et al. The role and interaction between transcription factor NAC-NOR and DNA demethylase SlDML2 in the biosynthesis of tomato fruit flavor volatiles. New Phytol. 2022, 235, 1913–1926. [Google Scholar] [CrossRef]
- Du, Q.; Lv, W.; Guo, Y.; Yang, J.; Wang, S.; Li, W.X. MIR164b represses iron uptake by regulating the NAC domain transcription factor5-Nuclear Factor Y, Subunit A8 module in Arabidopsis. Plant Physiol. 2022, 189, 1095–1109. [Google Scholar] [CrossRef]
- Song, H.; Liu, Y.; Dong, G.; Zhang, M.; Wang, Y.; Xin, J.; Su, Y.; Sun, H.; Yang, M. Genome-Wide Characterization and Comprehensive Analysis of NAC Transcription Factor Family in Nelumbo nucifera. Front. Genet. 2022, 13, 901838. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Zhang, Q.; Shi, Y.; Sun, Z.; Zhang, Q.; Hou, S.; Wu, R.; Jiang, L.; Zhao, X.; Guo, Y. Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao. Genes 2019, 11, 35. [Google Scholar] [CrossRef]
- Liu, M.; Ma, Z.; Sun, W.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide analysis of the NAC transcription factor family in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2019, 20, 113. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, J.; Du, B.; Zhang, M.; Wang, A.; Zhang, L. NAC Transcription Factor PwNAC11 Activates ERD1 by Interaction with ABF3 and DREB2A to Enhance Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 6952. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Liu, X.F.; Fu, B.L.; Zhang, Q.Y.; Tong, Y.; Wang, J.; Wang, W.Q.; Grierson, D.; Yin, X.R. Methyl Jasmonate Enhances Ethylene Synthesis in Kiwifruit by Inducing NAC Genes That Activate ACS1. J. Agric. Food Chem. 2020, 68, 3267–3276. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, M.; Wang, S. Processing characteristics and flavour of full lotus root powder beverage. J. Sci. Food Agric. 2010, 90, 2482–2489. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Li, S.; Hussain, J.; Xu, X.; Yin, J.; Zhang, Y.; Chen, X.; Li, L. Isolation and functional characterization of a salt responsive transcriptional factor, LrbZIP from lotus root (Nelumbo nucifera Gaertn). Mol. Biol. Rep. 2013, 40, 4033–4045. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Rombauts, S.; Dehais, P.; Van Montagu, M.; Rouze, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.P.; Xu, Z.S.; Zheng, W.J.; Zhao, W.; Wang, Y.X.; Yu, T.F.; Chen, M.; Zhou, Y.B.; Min, D.H.; Ma, Y.Z.; et al. Genome-Wide Analysis of the RAV Family in Soybean and Functional Identification of GmRAV-03 Involvement in Salt and Drought Stresses and Exogenous ABA Treatment. Front. Plant Sci. 2017, 8, 905. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).