
The Mayo Clinic Florida Microdosimetric Kinetic Model of Clonogenic Survival: Application to Various Repair-Competent Rodent and Human Cell Lines




Abstract
:1. Introduction
2. Results and Discussion
2.1. Rodent Cell Lines




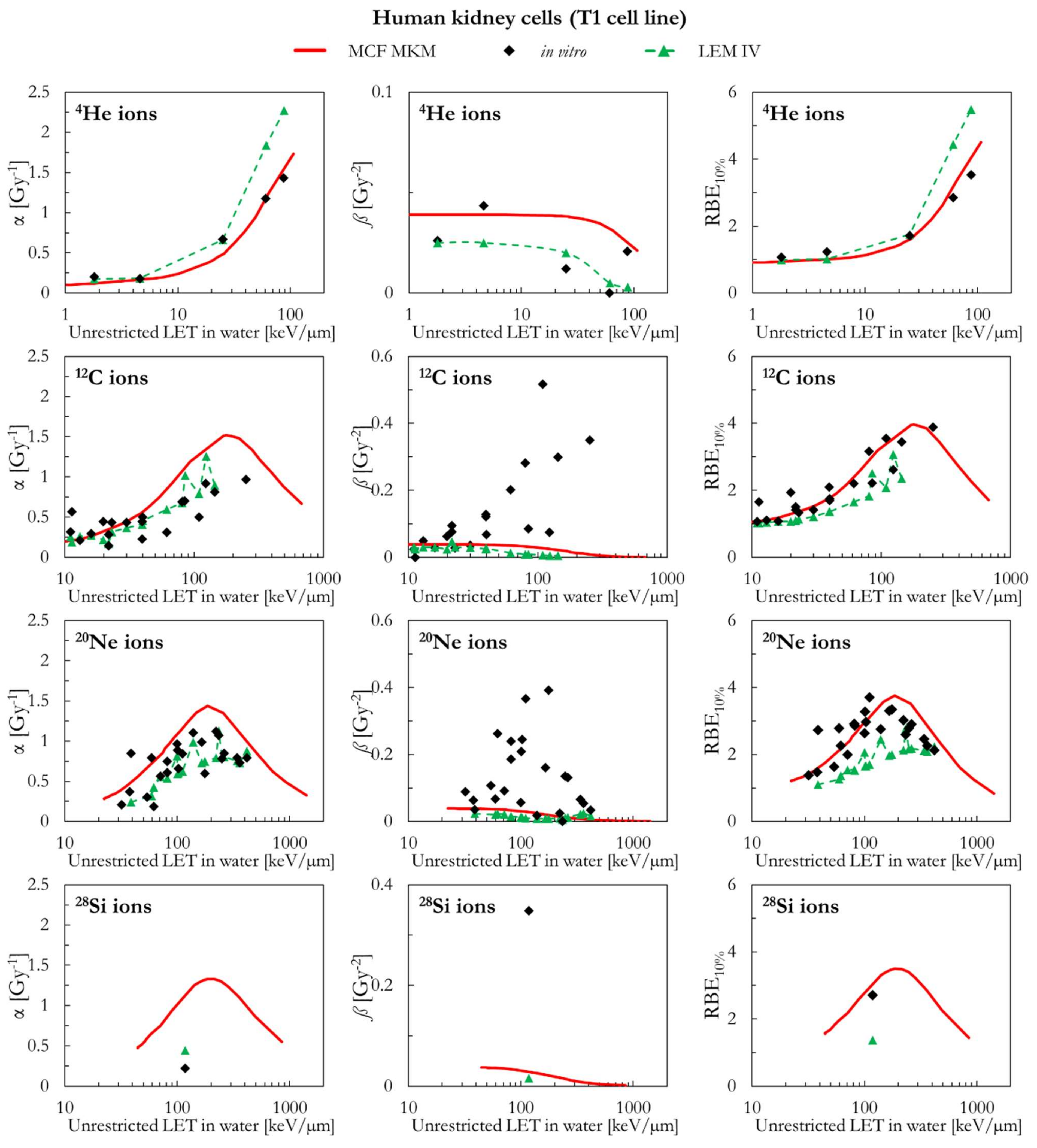
2.2. Human Cell Lines







3. Materials and Methods
3.1. Relative Biological Effectiveness
3.2. Clonogenic Survival Data
3.2.1. Particle Irradiation Data Ensemble
3.2.2. Initial Filtering
3.2.3. Data Selection for Rodent Cell Lines
3.2.4. Data Selection for Human Cell Lines
3.3. Biophysical Modeling
3.3.1. Model Parameters
Mean DNA Content of the Irradiated Population,
Mean Radius of the Cell Nucleus, Rn
Mean Radius of the Subnuclear Domains, rn
LQM Terms for the Reference Photon Exposure, αref and ßref
LQM Terms in the Limit of y → 0, α0 and ß0
3.3.2. Radiation Transport Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α | linear term of the linear-quadratic model of clonogenic survival |
| αref | α for the reference photon exposure |
| ß | quadratic term of the linear-quadratic model of clonogenic survival |
| ßref | ß for the reference photon exposure |
| DNA | deoxyribonucleic acid |
| DSB | double strand break |
| LEM | local effect model [6] |
| LEM IV | fourth version of the local effect model [26] |
| LET | linear energy transfer |
| LQM | linear-quadratic model of clonogenic survival [17,18] |
| MBM | mixed beam model [7] |
| MCF | Mayo Clinic Florida, Jacksonville, Florida, United States of America |
| MCF MKM | Mayo Clinic Florida microdosimetric kinetic model [33] |
| MKM | microdosimetric kinetic model [5] |
| modified MKM | modified microdosimetric kinetic model [9] |
| non-Poisson MKM | corrected version of the original microdosimetric kinetic model to account for the non-Poisson distribution of lethal lesions [8] |
| PHITS | Particle and Heavy Ion Transport code System [75] |
| PIDE | Particle Irradiation Data Ensemble [29,34] |
| RBE | relative biological effectiveness |
| RBES | in vitro clonogenic cell survival RBE for the surviving fraction S |
| rd | mean radius of the subnuclear domains |
| Rn | mean radius of the cell nucleus |
References
- Scholz, M. State-of-the-Art and Future Prospects of Ion Beam Therapy: Physical and Radiobiological Aspects. IEEE Trans. Radiat. Plasma Med. Sci. 2020, 4, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.D.; Carlson, D.J.; Butkus, M.P.; Hawkins, R.; Friedrich, T.; Scholz, M. A comparison of mechanism-inspired models for particle relative biological effectiveness (RBE). Med. Phys. 2018, 45, e925–e952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganetti, H. Relating the proton relative biological effectiveness to tumor control and normal tissue complication probabilities assuming interpatient variability in α/β. Acta Oncol. 2017, 56, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Kellerer, A.M.; Rossi, H.H. A Generalized Formulation of Dual Radiation Action. Radiat. Res. 1978, 75, 471. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, R.B. A Statistical Theory of Cell Killing by Radiation of Varying Linear Energy Transfer. Radiat. Res. 1994, 140, 366. [Google Scholar] [CrossRef]
- Scholz, M.; Kellerer, A.M.; Kraft-Weyrather, W.; Kraft, G. Computation of cell survival in heavy ion beams for therapy. Radiat. Environ. Biophys. 1997, 36, 59–66. [Google Scholar] [CrossRef]
- Kanai, T.; Endo, M.; Minohara, S.; Miyahara, N.; Koyama-Ito, H.; Tomura, H.; Matsufuji, N.; Futami, Y.; Fukumura, A.; Hiraoka, T.; et al. Biophysical characteristics of HIMAC clinical irradiation system for heavy-ion radiation therapy. Int. J. Radiat. Oncol. 1999, 44, 201–210. [Google Scholar] [CrossRef]
- Hawkins, R.B. A Microdosimetric-Kinetic Model for the Effect of Non-Poisson Distribution of Lethal Lesions on the Variation of RBE with LET. Radiat. Res. 2003, 160, 61–69. [Google Scholar] [CrossRef]
- Kase, Y.; Kanai, T.; Matsumoto, Y.; Furusawa, Y.; Okamoto, H.; Asaba, T.; Sakama, M.; Shinoda, H. Microdosimetric Measurements and Estimation of Human Cell Survival for Heavy-Ion Beams. Radiat. Res. 2006, 166, 629–638. [Google Scholar] [CrossRef]
- Carlson, D.J.; Stewart, R.D.; Semenenko, V.A.; Sandison, G.A. Combined use of Monte Carlo DNA damage simulations and deterministic repair models to examine putative mechanisms of cell killing. Radiat. Res. 2008, 169, 447–459. [Google Scholar] [CrossRef]
- Sato, T.; Furusawa, Y. Cell Survival Fraction Estimation Based on the Probability Densities of Domain and Cell Nucleus Specific Energies Using Improved Microdosimetric Kinetic Models. Radiat. Res. 2012, 178, 341–356. [Google Scholar] [CrossRef]
- Mairani, A.; Böhlen, T.T.; Dokic, I.; Cabal, G.; Brons, S.; Haberer, T. Modelling of cell killing due to sparsely ionizing radiation in normoxic and hypoxic conditions and an extension to high LET radiation. Int. J. Radiat. Biol. 2013, 89, 782–793. [Google Scholar] [CrossRef]
- Verkhovtsev, A.; Surdutovich, E.; Solov’yov, A.V. Multiscale approach predictions for biological outcomes in ion-beam cancer therapy. Sci. Rep. 2016, 6, 1–10. [Google Scholar]
- McMahon, S.J.; McNamara, A.L.; Schuemann, J.; Paganetti, H.; Prise, K.M. A general mechanistic model enables predictions of the biological effectiveness of different qualities of radiation. Sci. Rep. 2017, 7, 10790. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Li, C.; Qiu, R.; Chen, Y.; Wu, Z.; Zhang, H.; Li, J. Modelling of cellular survival following radiation-induced DNA double-strand breaks. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fujitaka, S.; Fujii, Y.; Nihongi, H.; Nakayama, S.; Takashina, M.; Hamatani, N.; Tsubouchi, T.; Yagi, M.; Minami, K.; Ogawa, K.; et al. Physical and biological beam modeling for carbon beam scanning at Osaka Heavy Ion Therapy Center. J. Appl. Clin. Med. Phys. 2021, 22, 77–92. [Google Scholar] [CrossRef]
- Fowler, J.F. The linear-quadratic formula and progress in fractionated radiotherapy. British J. Radiol. 1989, 62, 679–694. [Google Scholar] [CrossRef]
- McMahon, S.J. The linear quadratic model: Usage, interpretation and challenges. Phys. Med. Biol. 2018, 64, 01TR01. [Google Scholar] [CrossRef]
- Sakama, M.; Kanai, T.; Kase, Y.; Yusa, K.; Tashiro, M.; Torikai, K.; Shimada, H.; Yamada, S.; Ohno, T.; Nakano, T. Design of ridge filters for spread-out Bragg peaks with Monte Carlo simulation in carbon ion therapy. Phys. Med. Biol. 2012, 57, 6615–6633. [Google Scholar] [CrossRef]
- Agosteo, S. Detectors for measurement of microdosimetric quantities. Radiat. Meas. 2022, 156, 106807. [Google Scholar] [CrossRef]
- Mein, S.; Klein, C.; Kopp, B.; Magro, G.; Harrabi, S.; Karger, C.P.; Haberer, T.; Debus, J.; Abdollahi, A.; Dokic, I.; et al. Assessment of RBE-weighted dose models for carbon ion therapy toward modernization of clinical practice at HIT: In vitro, in vivo, and in patients. Int. J. Radiat. Oncol. Biol. Phys. 2020, 108, 779–791. [Google Scholar] [CrossRef]
- Eichkorn, T.; König, L.; Held, T.; Naumann, P.; Harrabi, S.; Ellerbrock, M.; Herfarth, K.; Haberer, T.; Debus, J. Carbon Ion Radiation Therapy: One Decade of Research and Clinical Experience at Heidelberg Ion Beam Therapy Center. Int. J. Radiat. Oncol. Biol. Phys. 2021, 111, 597–609. [Google Scholar] [CrossRef]
- Eichkorn, T.; Karger, C.P.; Brons, S.; Koerber, S.A.; Mielke, T.; Haberer, T.; Debus, J.; Herfarth, K. Results of a pro-spective randomized trial on long-term effectiveness of protons and carbon ions in prostate cancer: LEM I and α/β = 2 Gy overestimates the RBE. Radiother. Oncol. 2022, 173, 223–230. [Google Scholar] [CrossRef]
- Elsässer, T.; Scholz, M. Cluster Effects within the Local Effect Model. Radiat. Res. 2007, 167, 319–329. [Google Scholar] [CrossRef]
- Elsässer, T.; Krämer, M.; Scholz, M. Accuracy of the Local Effect Model for the Prediction of Biologic Effects of Carbon Ion Beams In Vitro and In Vivo. Int. J. Radiat. Oncol. Biol. Phys. 2008, 71, 866–872. [Google Scholar] [CrossRef]
- Elsässer, T.; Weyrather, W.K.; Friedrich, T.; Durante, M.; Iancu, G.; Krämer, M.; Kragl, G.; Brons, S.; Winter, M.; Weber, K.J.; et al. Quantification of the relative biological effectiveness for ion beam radiotherapy: Direct experimental comparison of proton and carbon ion beams and a novel approach for treatment planning. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, 1177–1183. [Google Scholar] [CrossRef]
- Friedrich, T.; Grün, R.; Scholz, U.; Elsässer, T.; Durante, M.; Scholz, M. Sensitivity analysis of the relative biological effectiveness predicted by the local effect model. Phys. Med. Biol. 2013, 58, 6827–6849. [Google Scholar] [CrossRef]
- Pfuhl, T.; Friedrich, T.; Scholz, M. Comprehensive comparison of local effect model IV predictions with the particle irradiation data ensemble. Med. Phys. 2022, 49, 714–726. [Google Scholar] [CrossRef]
- Friedrich, T.; Scholz, U.; Elsässer, T.; Durante, M.; Scholz, M. Systematic analysis of RBE and related quantities using a database of cell survival experiments with ion beam irradiation. J. Radiat. Res. 2013, 54, 494–514. [Google Scholar] [CrossRef] [Green Version]
- Kellerer, A.M.; Rossi, H.H. The theory of dual radiation action. Curr. Top. Radiat. Res. Q. 1974, 8, 85–158. [Google Scholar]
- Parisi, A.; Furutani, K.M.; Beltran, C.J. On the calculation of the relative biological effectiveness of ion radiation therapy using a biological weighting function, the microdosimetric kinetic model (MKM) and subsequent corrections (non-Poisson MKM and modified MKM). Phys. Med. Biol. 2022, 67, 095014. [Google Scholar] [CrossRef]
- Beltran, C.; Amos, R.A.; Rong, Y. We are ready for clinical implementation of Carbon Ion Radiotherapy in the United States. J. Appl. Clin. Med. Phys. 2020, 21, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Parisi, A.; Beltran, C.J.; Furutani, K.M. The Mayo Clinic Florida microdosimetric kinetic model of clonogenic survival: Formalism and first benchmark against in vitro and in silico data. Phys. Med. Biol. 2022, 67, 185013. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, T.; Pfuhl, T.; Scholz, M. Update of the particle irradiation data ensemble (PIDE) for cell survival. J. Radiat. Res. 2021, 62, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Parisi, A.; Sato, T.; Matsuya, Y.; Kase, Y.; Magrin, G.; Verona, C.; Tran, L.; Rosenfeld, A.; Bianchi, A.; Olko, P.; et al. Development of a new microdosimetric biological weighting function for the RBE10 assessment in case of the V79 cell line exposed to ions from 1H to 238U. Phys. Med. Biol. 2020, 65, 235010. [Google Scholar] [CrossRef] [PubMed]
- Parisi, A.; Struelens, L.; Vanhavere, F. Comparison between the results of a recently-developed biological weighting function (V79-RBE10 BWF) and the in vitro clonogenic survival RBE10 of other repair-competent asynchronized normoxic mammalian cell lines and ions not used for the development of the model. Phys. Med. Biol. 2021, 66, 235006. [Google Scholar] [CrossRef]
- McMahon, S.J.; Schuemann, J.; Paganetti, H.; Prise, K. Mechanistic Modelling of DNA Repair and Cellular Survival Following Radiation-Induced DNA Damage. Sci. Rep. 2016, 6, 33290. [Google Scholar] [CrossRef] [Green Version]
- Capes-Davis, A.; Theodosopoulos, G.; Atkin, I.; Drexler, H.G.; Kohara, A.; MacLeod, R.A.; Masters, J.R.; Nakamura, Y.; Reid, Y.A.; Reddel, R.; et al. Check your cultures! A list of cross-contaminated or misidentified cell lines. Int. J. Cancer 2010, 127, 1–8. [Google Scholar] [CrossRef]
- Allen, M.; Bjerke, M.; Edlund, H.; Nelander, S.; Westermark, B. Origin of the U87MG glioma cell line: Good news and bad news. Sci. Transl. Med. 2016, 8, 354re3. [Google Scholar] [CrossRef]
- Guan, F.; Bronk, L.; Titt, U.; Lin, S.H.; Mirkovic, D.; Kerr, M.D.; Zhu, X.R.; Dinh, J.; Sobieski, M.; Stephan, C.; et al. Spatial mapping of the biologic effectiveness of scanned particle beams: Towards biologically optimized particle therapy. Sci. Rep. 2015, 5, 9850. [Google Scholar] [CrossRef] [Green Version]
- Seed, T.M.; Xiao, S.; Manley, N.R.; Nikolich-Zugich, J.; Pugh, J.; Brink, M.V.D.; Hirabayashi, Y.; Yasutomo, K.; Iwama, A.; Koyasu, S.; et al. An interlaboratory comparison of dosimetry for a multi-institutional radiobiological research project: Observations, problems, solutions and lessons learned. Int. J. Radiat. Biol. 2015, 92, 59–70. [Google Scholar] [CrossRef]
- Trompier, F.; Baumann, M.; Barrios, L.; Gregoire, E.; Abend, M.; Ainsbury, E.; Barnard, S.; Barquinero, J.F.; Bautista, J.A.; Brzozowska, B.; et al. Investigation of the influence of calibration practices on cytogenetic laboratory performance for dose estimation. Int. J. Radiat. Biol. 2017, 93, 118–126. [Google Scholar] [CrossRef]
- Durante, M.; Paganetti, H.; Pompos, A.; Kry, S.F.; Wu, X.; Grosshans, D.R. Report of a National Cancer Institute special panel: Characterization of the physical parameters of particle beams for biological research. Med. Phys. 2019, 46, e37–e52. [Google Scholar] [CrossRef] [Green Version]
- Draeger, E.; Sawant, A.; Johnstone, C.; Koger, B.; Becker, S.; Vujaskovic, Z.; Jackson, I.-L.; Poirier, Y. A Dose of Reality: How 20 Years of Incomplete Physics and Dosimetry Reporting in Radiobiology Studies May Have Contributed to the Reproducibility Crisis. Int. J. Radiat. Oncol. Biol. Phys. 2020, 106, 243–252. [Google Scholar] [CrossRef]
- Parisi, A.; Beltran, C.J.; Furutani, K.M. The effect of fitting the reference photon dose-response on the clonogenic survival RBE calculated with the Mayo Clinic Florida MKM. Radiat. Prot. Dosim. 2022, in press. [Google Scholar]
- Wulf, H.; Kraft-Weyrather, W.; Miltenburger, H.G.; Blakely, E.A.; Tobias, C.A.; Kraft, G. Heavy-Ion Effects on Mammalian Cells: Inactivation Measurements with Different Cell Lines. Radiat. Res. 1985, 104, S122. [Google Scholar] [CrossRef]
- Von Neubeck, C. Radiobiological Experiments for Carbon Ion Prostate Cancer Therapy: Interplay of Normal and Tumor Cells in Co-Culture and Measurement of the Oxygen Enhancement Ratio. Ph.D. Thesis, Technische Universität Darmstadt, Germany, 2009. [Google Scholar]
- Quintanilla, M.; Haddow, S.; Jones, D.; Jaffe, D.; Bowden, G.; Balmain, A. Comparison of ras activation during epidermal carcinogenesis in vitro and in vivo. Carcinogenesis 1991, 12, 1875–1881. [Google Scholar] [CrossRef]
- Chaudhary, P.; Marshall, T.I.; Perozziello, F.M.; Manti, L.; Currell, F.J.; Hanton, F.; McMahon, S.J.; Kavanagh, J.N.; Cirrone, G.A.P.; Romano, F.; et al. Relative Biological Effectiveness Variation Along Monoenergetic and Modulated Bragg Peaks of a 62-MeV Therapeutic Proton Beam: A Preclinical Assessment. Int. J. Radiat. Oncol. Biol. Phys. 2014, 90, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, K.; Moritake, T.; Tsuchida, Y.; Tokuuye, K.; Matsumura, A.; Ando, K. Cell Cycle Checkpoint and Apoptosis Induction in Glioblastoma Cells and Fibroblasts Irradiated with Carbon Beam. J. Radiat. Res. 2007, 48, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Krämer, M.; Scholz, M. Rapid calculation of biological effects in ion radiotherapy. Phys. Med. Biol. 2006, 51, 1959–1970. [Google Scholar] [CrossRef]
- Ohno, H.; Doi, S.; Fukuhara, S.; Yamasowa, M.; Kannagi, M.; Ishikura, H.; Nishikori, M.; Uchino, H. Establishment of a new myeloid leukemia cell line (TK-1), and isolation of cells having a translocation involving a band 17q23. Int. J. Cancer 1986, 37, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Macville, M.; Schröck, E.; Padilla-Nash, H.; Keck, C.; Ghadimi, B.M.; Zimonjic, D.; Popescu, N.; Ried, T. Compre-hensive and definitive molecular cytogenetic characterization of HeLa cells by spectral karyotyping. Cancer Res. 1999, 59, 141–150. [Google Scholar] [PubMed]
- Liang, J.C.; Ning, Y.; Wang, R.-Y.; Padilla-Nash, H.M.; Schröck, E.; Soenksen, D.; Nagarajan, L.; Ried, T. Spectral Karyotypic Study of the HL-60 Cell Line: Detection of Complex Rearrangements Involving Chromosomes 5, 7, and 16 and Delineation of Critical Region of Deletion on 5q31.1. Cancer Genet. Cytogenet. 1999, 113, 105–109. [Google Scholar] [CrossRef]
- Fusenig, N.; Dzarlieva-Petrusevska, R.T.; Breitkreutz, D. Phenotypic and cytogenetic characteristics of different stages during spontaneous transformation of mouse keratinocytes in vitro. Carcinog. Compr. Surv. 1985, 9, 293–326. [Google Scholar]
- Oesten, H.; von Neubeck, C.; Jakob, A.; Enghardt, W.; Krause, M.; McMahon, S.J.; Grassberger, C.; Paganetti, H.; Lühr, A. Predicting In Vitro Cancer Cell Survival Based on Measurable Cell Characteristics. Radiat. Res. 2019, 191, 532–544. [Google Scholar] [CrossRef]
- Lammerding, J. Mechanics of the nucleus. Compr. Physiol. 2011, 1, 783. [Google Scholar]
- Tracy, B.L.; Stevens, D.L.; Goodhead, D.T.; Hill, M.A. Variation in RBE for Survival of V79-4 Cells as a Function of Alpha-Particle (Helium Ion) Energy. Radiat. Res. 2015, 184, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Kashani, A.S.; Packirisamy, M. Cellular deformation characterization of human breast cancer cells under hydro-dynamic forces. AIMS Biophys. 2017, 4, 400–414. [Google Scholar] [CrossRef]
- Sakata, D.; Lampe, N.; Karamitros, M.; Kyriakou, I.; Belov, O.; Bernal, M.A.; Bolst, D.; Bordage, M.-C.; Breton, V.; Brown, J.M.; et al. Evaluation of early radiation DNA damage in a fractal cell nucleus model using Geant4-DNA. Phys. Med. 2019, 62, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Weyrather, W.K.; Ritter, S.; Scholz, M.; Kraft, G. RBE for carbon track-segment irradiation in cell lines of differing repair capacity. Int. J. Radiat. Biol. 1999, 75, 1357–1364. [Google Scholar]
- Konishi, T.; Takeyasu, A.; Yasuda, N.; Natsume, T.; Nakajima, H.; Matsumoto, K.; Asuka, T.; Sato, Y.; Furusawa, Y.; Hieda, K. Number of Fe ion traversals through a cell nucleus for mammalian cell inactivation near the Bragg peak. J. Radiat. Res. 2005, 46, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Gacsi, M.; Nagy, G.; Pinter, G.; Basnakian, A.G.; Banfalvi, G. Condensation of interphase chromatin in nuclei of synchronized Chinese hamster ovary (CHO-K1) cells. DNA Cell Biol. 2005, 24, 43–53. [Google Scholar] [CrossRef]
- Monier, K.; Armas, J.C.G.; Etteldorf, S.; Ghazal, P.; Sullivan, K.F. Annexation of the interchromosomal space during viral infection. Nat. Cell Biol. 2000, 2, 661–665. [Google Scholar] [CrossRef]
- Suzuki, M.; Kase, Y.; Yamaguchi, H.; Kanai, T.; Ando, K. Relative biological effectiveness for cell-killing effect on various human cell lines irradiated with heavy-ion medical accelerator in Chiba (HIMAC) carbon-ion beams. Int. J. Radiat. Oncol. Biol. Phys. 2000, 48, 241–250. [Google Scholar] [CrossRef]
- Azzam, E.I.; De Toledo, S.M.; Gooding, T.; Little, J.B. Intercellular communication is involved in the bystander regulation of gene expression in human cells exposed to very low fluences of alpha particles. Radiat. Res. 1998, 150, 497–504. [Google Scholar] [CrossRef]
- Chaudhary, P.; Marshall, T.I.; Currell, F.J.; Kacperek, A.; Schettino, G.; Prise, K.M. Variations in the processing of DNA double-strand breaks along 60-MeV therapeutic proton beams. Int. J. Radiat. Oncol. Biol. Phys. 2016, 95, 86–94. [Google Scholar] [CrossRef]
- Kassis, A.I.; Fayad, F.; Kinsey, B.M.; Sastry, K.S.; Adelstein, S.J. Radiotoxicity of an 125I-labeled DNA intercalator in mammalian cells. Radiat. Res. 1989, 118, 283–294. [Google Scholar] [CrossRef]
- Howell, R.W.; Rao, D.V.; Hou, D.Y.; Narra, V.R.; Sastry, K.S. The question of relative biological effectiveness and quality factor for Auger emitters incorporated into proliferating mammalian cells. Radiat. Res. 1991, 128, 282–292. [Google Scholar] [CrossRef]
- Friedrich, T.; Durante, M.; Scholz, M. Modeling Cell Survival after Photon Irradiation Based on Double-Strand Break Clustering in Megabase Pair Chromatin Loops. Radiat. Res. 2012, 178, 385–394. [Google Scholar] [CrossRef]
- Yokota, H.; Van Den Engh, G.; Hearst, J.E.; Sachs, R.K.; Trask, B.J. Evidence for the organization of chromatin in megabase pair-sized loops arranged along a random walk path in the human G0/G1 interphase nucleus. J. Cell Biol. 1995, 130, 1239–1249. [Google Scholar] [CrossRef] [Green Version]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA Double-stranded Breaks Induce Histone H2AX Phosphorylation on Serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Watanabe, R.; Niita, K. Development of a calculation method for estimating specific energy distribution in complex radiation fields. Radiat. Prot. Dosim. 2006, 122, 41–45. [Google Scholar] [CrossRef]
- Sato, T.; Watanabe, R.; Sihver, L.; Niita, K. Applications of the microdosimetric function implemented in the mac-roscopic particle transport simulation code PHITS. Int. J. Radiat. Biol. 2012, 88, 143–150. [Google Scholar] [CrossRef]
- Sato, T.; Iwamoto, Y.; Hashimoto, S.; Ogawa, T.; Furuta, T.; Abe, S.-I.; Kai, T.; Tsai, P.-E.; Matsuda, N.; Iwase, H.; et al. Features of Particle and Heavy Ion Transport code System (PHITS) version 3.02. J. Nucl. Sci. Technol. 2018, 55, 684–690. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line Abbreviation | Species | Type of Cells | Number of Entries | Ions |
|---|---|---|---|---|
| C3H10T1/2 | mouse | embryonic fibroblasts | 20 | 1H, 2H, 3He, 4He, 12C, 16O, 20Ne, 28Si, 40Ar, 56Fe, 238U |
| CHO, CHO-K1 | Chinese hamster | ovary epithelial cells | 38 | 12C, 16O, 20Ne, 40Ar, 56Fe |
| HeLa | human | cervical cancer cells | 4 | 1H, 4He |
| HF19 | human | fetal lung fibroblasts | 6 | 1H, 12C |
| HL-60 | human | leukemia cells | 7 | 12C, 28Si, 56Fe |
| M/10 | human | mammary epithelial cells | 6 | 12C |
| NB1RGB | human | skin fibroblasts | 29 | 12C, 20Ne, 28Si |
| PDV | mouse | transformed epidermal cells | 4 | 1H, 7Li |
| RAT-1 | rat | prostatic adenocarcinoma epithelial cells | 3 | 12C |
| SQ20B | human | laryngeal squamous cell carcinoma | 11 | 1H, 12C, 40Ar |
| T1 | human | kidney cells | 63 | 4He, 12C, 20Ne, 28Si, 40Ar, 56Fe, 238U |
| TK1 | human | myeloid leukemia cells | 10 | 12C |
| U-87 | human | glioblastoma cells | 6 | 1H |
| U-251MG | human | astrocytoma cells | 4 | 12C |
| Cell Line Abbreviation | α for the Reference Photon Exposure, [Gy−1] | β for the Reference Photon Exposure, [Gy−2] | [Gy] | Mean Radius of the Cell Nucleus, Rn [µm] | Mean Radius of the Subnuclear Domains, rd [µm] |
|---|---|---|---|---|---|
| C3H10T1/2 | 0.173 * | 0.389 * | 4.44 | 4.0 | 0.26 |
| CHO, CHO-K1 | 0.226 * | 0.0231 * | 9.78 | 4.2 | 0.27 |
| HeLa | 0.536 | 0.0278 | 19.3 | 5.6 | 0.29 |
| HF19 | 0.557 * | 0.0189 * | 29.5 | 4.7 ** | 0.29 |
| HL-60 | 0.315 | 0.0558 | 5.64 | 4.6 ** | 0.29 |
| M/10 | 0.3 | 0.068 | 4.41 | 4.7 ** | 0.29 |
| NB1RGB | 0.476 * | 0.0458 * | 10.4 | 5.1 | 0.32 |
| PDV | 0.13 | 0.037 | 3.51 | 5.1 ** | 0.29 |
| RAT-1 | 0.201 | 0.0266 | 7.53 | 5.0 ** | 0.29 |
| SQ20B | 0.122 * | 0.0238 * | 5.12 | 4.5 ** | 0.30 |
| T1 | 0.159 * | 0.0391 * | 4.06 | 4.7 ** | 0.29 |
| TK1 | 0.107 * | 0.0384 * | 2.79 | 4.7 ** | 0.29 |
| U-87 | 0.106 | 0.0557 | 1.91 | 4.5 ** | 0.30 |
| U-251MG | 0.031 | 0.0551 | 0.563 | 4.9 ** | 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parisi, A.; Beltran, C.J.; Furutani, K.M. The Mayo Clinic Florida Microdosimetric Kinetic Model of Clonogenic Survival: Application to Various Repair-Competent Rodent and Human Cell Lines. Int. J. Mol. Sci. 2022, 23, 12491. https://doi.org/10.3390/ijms232012491
Parisi A, Beltran CJ, Furutani KM. The Mayo Clinic Florida Microdosimetric Kinetic Model of Clonogenic Survival: Application to Various Repair-Competent Rodent and Human Cell Lines. International Journal of Molecular Sciences. 2022; 23(20):12491. https://doi.org/10.3390/ijms232012491
Chicago/Turabian StyleParisi, Alessio, Chris J. Beltran, and Keith M. Furutani. 2022. "The Mayo Clinic Florida Microdosimetric Kinetic Model of Clonogenic Survival: Application to Various Repair-Competent Rodent and Human Cell Lines" International Journal of Molecular Sciences 23, no. 20: 12491. https://doi.org/10.3390/ijms232012491
APA StyleParisi, A., Beltran, C. J., & Furutani, K. M. (2022). The Mayo Clinic Florida Microdosimetric Kinetic Model of Clonogenic Survival: Application to Various Repair-Competent Rodent and Human Cell Lines. International Journal of Molecular Sciences, 23(20), 12491. https://doi.org/10.3390/ijms232012491