
Integrated Analysis of Cytokine Profiles in Malaria Patients Discloses Selective Upregulation of TGF-β1, β3, and IL-9 in Mild Clinical Presentation

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Demographic Characteristics of Participants
2.2. Identification of Human Plasmodium Infections Infecting the Participants
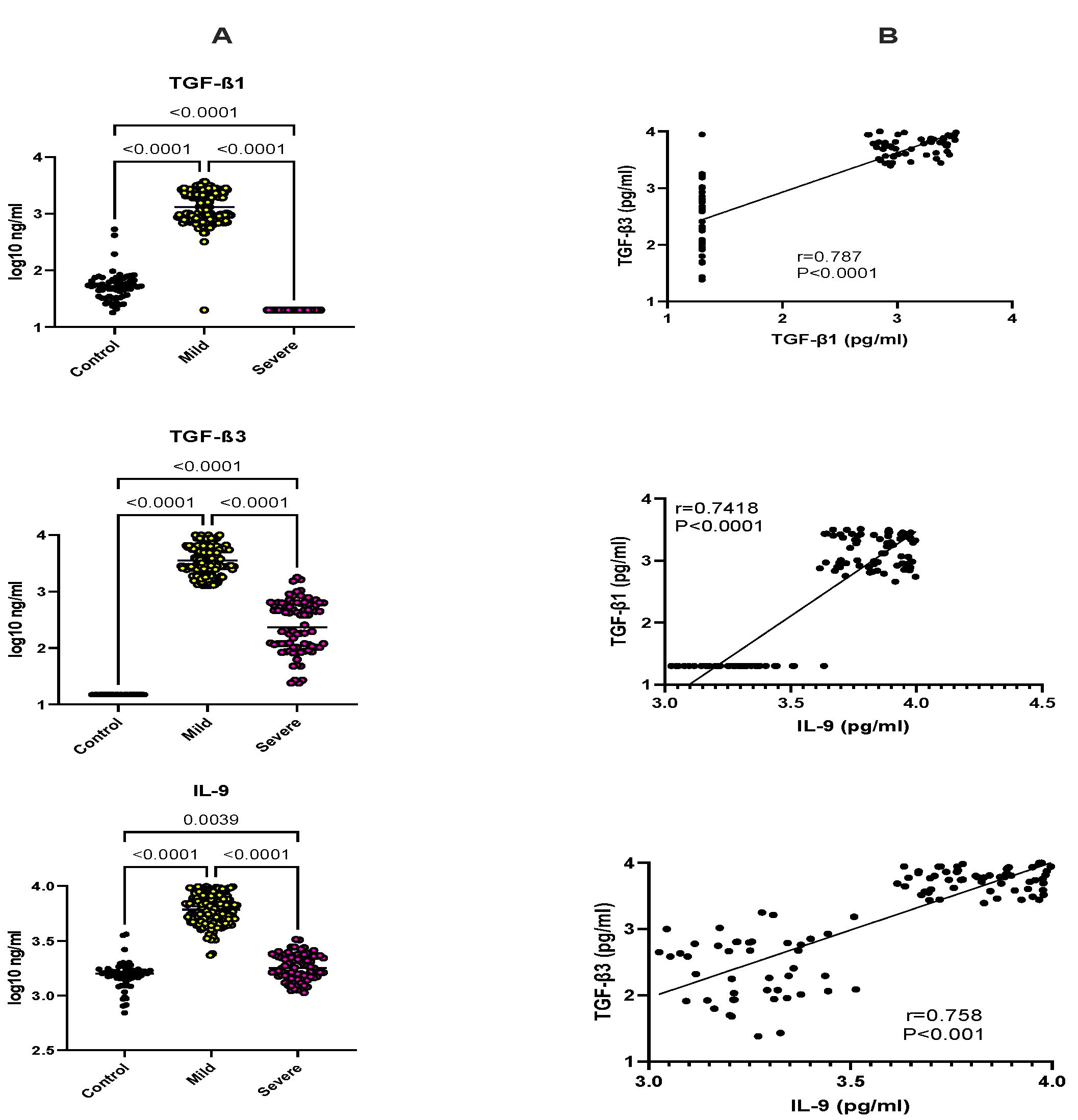
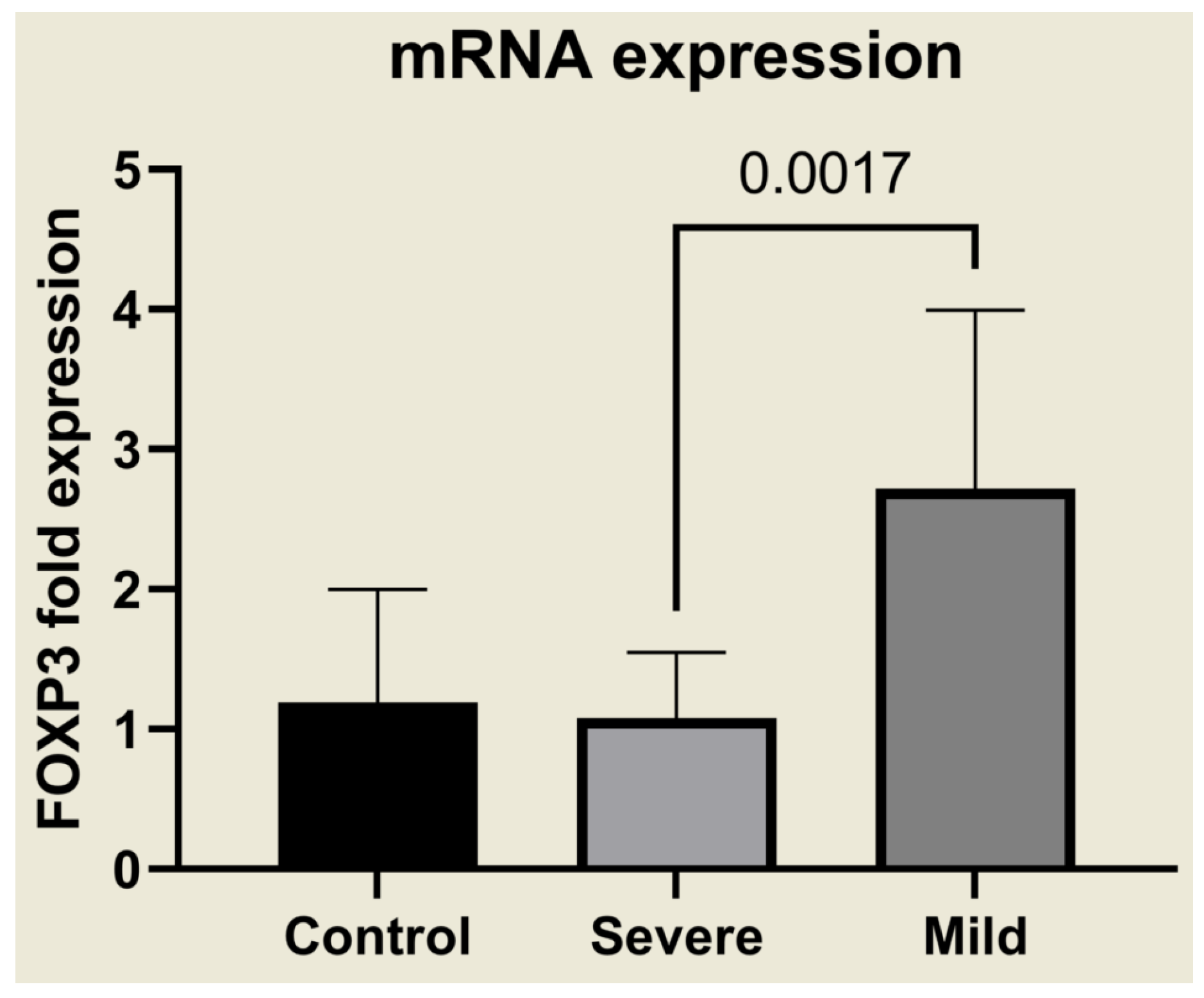
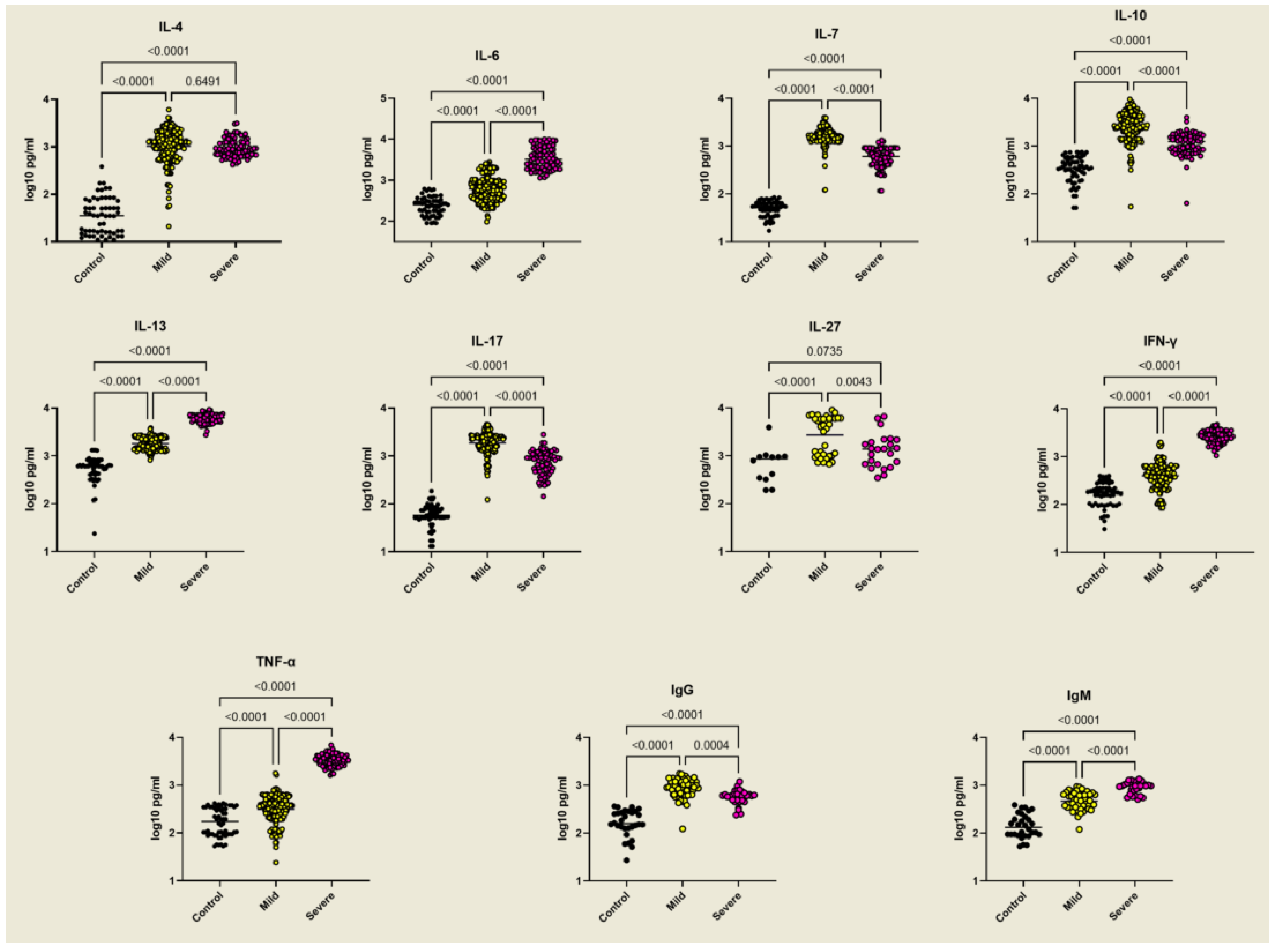
2.3. Selective Upregulation of TGF-β1, TGF-β3, IL-9, and FOXP3 in Mild Malaria
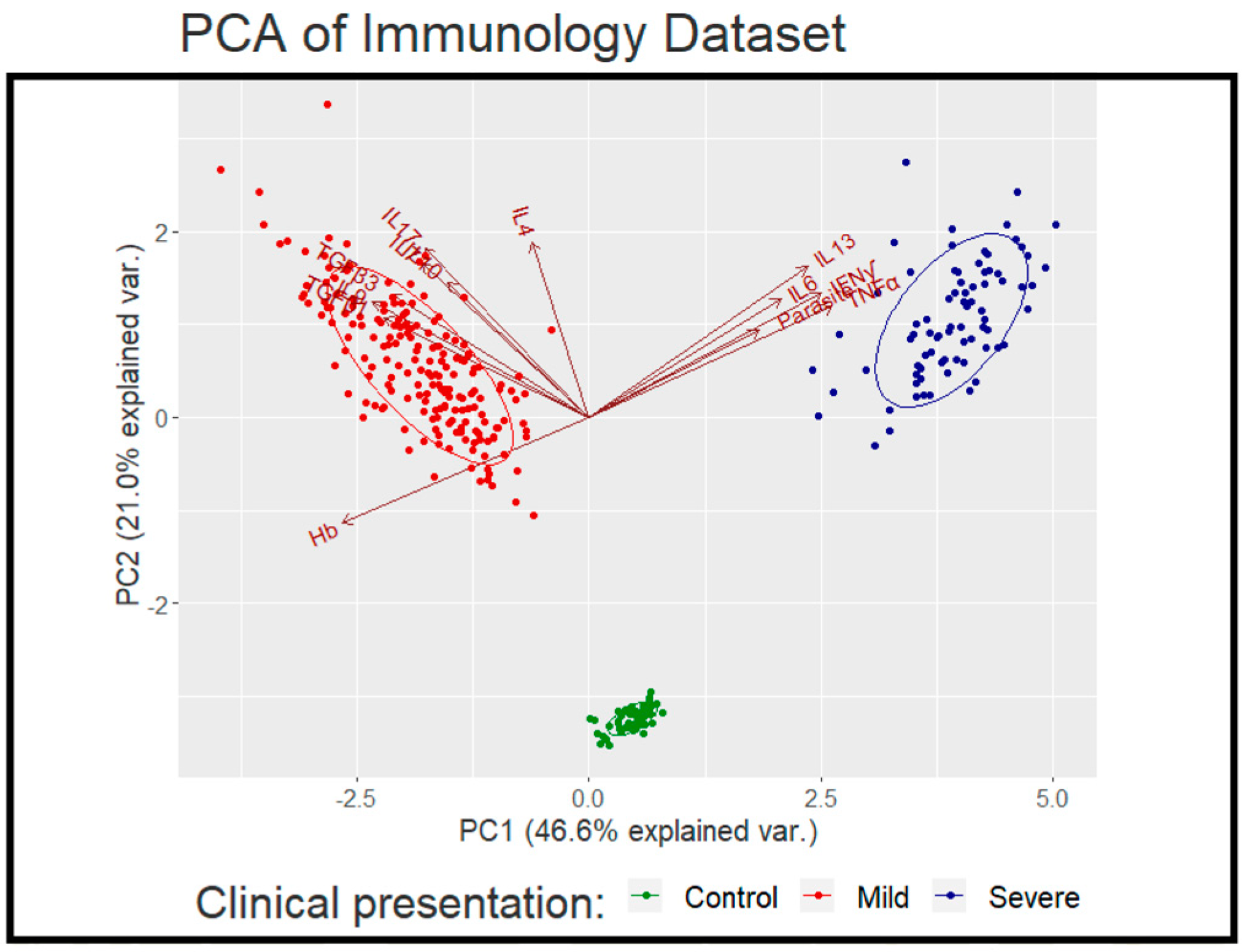
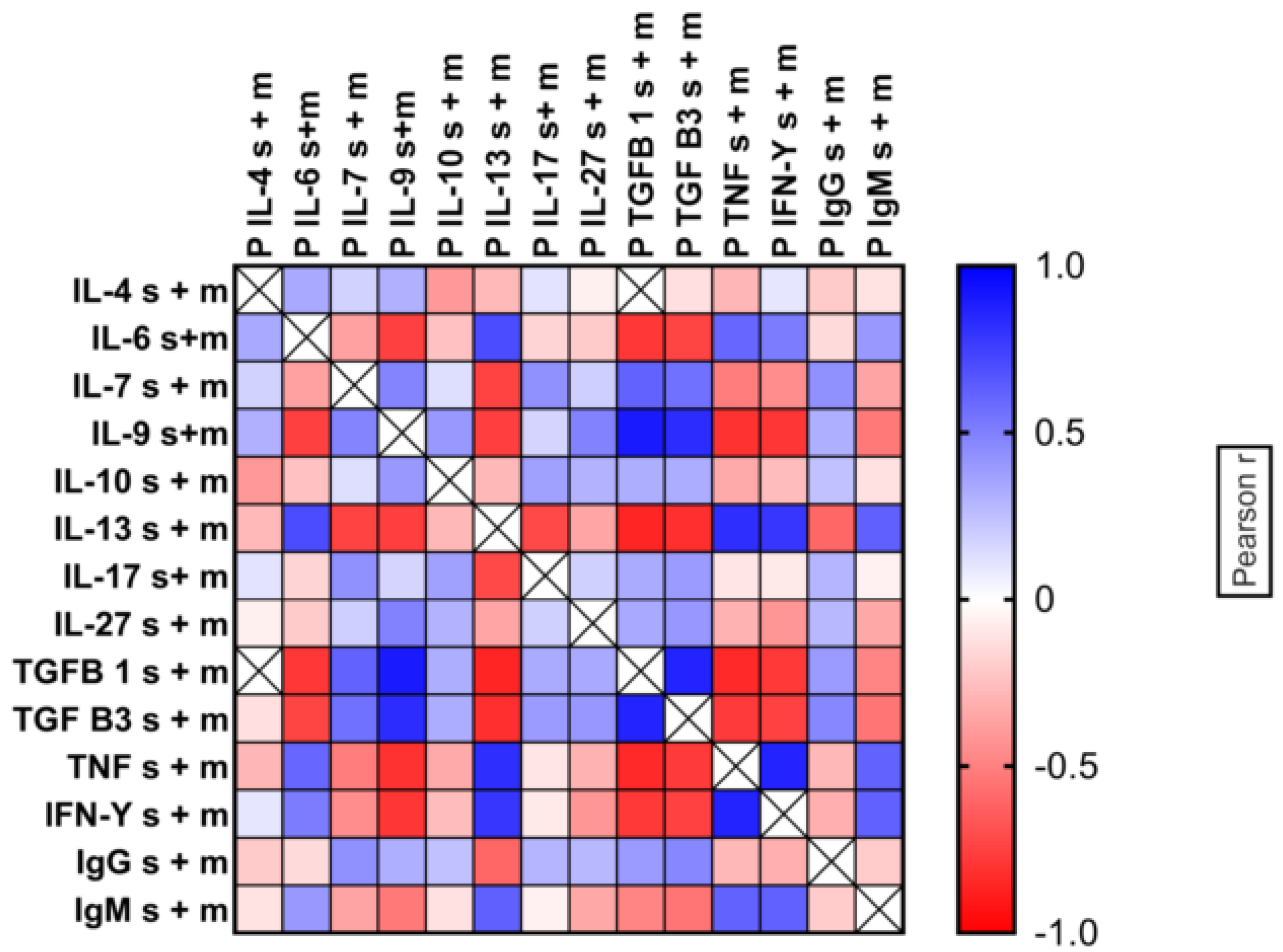
2.4. Multi-Factor Analysis in Mild and Severe Malaria
2.5. Patterns of Cytokine Expression in Individual Patients
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Malaria Diagnosis
4.3. Cytokine Enzyme-Linked Immunosorbent Assay (ELISAs)
4.4. Evaluation of FOXP3 Expression via Quantitative RT-PCR
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Litsios, S. The World Health Organization’s changing goals and expectations concerning malaria, 1948–2019. Hist. Cienc. Saude Manguinhos 2020, 27, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Artavanis-Tsakonas, K.; Tongren, J.E.; Riley, E.M. The war between the malaria parasite and the immune system: Immunity, immunoregulation and immunopathology. Clin. Exp. Immunol. 2003, 133, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Drewry, L.L.; Harty, J.T. Balancing in a black box: Potential immunomodulatory roles for TGF-beta signalling during blood-stage malaria. Virulence 2020, 11, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omer, F.M.; Riley, E.M. Transforming growth factor beta production is inversely correlated with severity of murine malaria infection. J. Exp. Med. 1998, 188, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Omer, F.M.; de Souza, J.B.; Riley, E.M. Differential induction of TGF-beta regulates proinflammatory cytokine production and determines the outcome of lethal and nonlethal Plasmodium yoelii infections. J. Immunol. 2003, 171, 5430–5436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, P.; Li, J.; Kulkarni, A.B.; Perruche, S.; Chen, W. A critical function for TGF-beta signaling in the development of natural CD4+CD25+Foxp3+ regulatory T cells. Nat. Immunol. 2008, 9, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Wenisch, C.; Parschalk, B.; Burgmann, H.; Looareesuwan, S.; Graninger, W. Decreased serum levels of TGF-beta in patients with acute Plasmodium falciparum malaria. J. Clin. Immunol. 1995, 15, 69–73. [Google Scholar] [CrossRef]
- Chaiyaroj, S.C.; Rutta, A.S.; Muenthaisong, K.; Watkins, P.; Ubol, M.N.; Looareesuwan, S. Reduced levels of transforming growth factor-beta1, interleukin-12 and increased migration inhibitory factor are associated with severe malaria. Acta Trop. 2004, 89, 319–327. [Google Scholar] [CrossRef]
- De Jong, G.M.; McCall, M.B.B.; Dik, W.A.; Urbanus, R.T.; Wammes, L.J.; Koelewijn, R.; Sauerwein, R.W.; Verbon, A.; van Hellemond, J.J.; van Genderen, P.J.J. Transforming growth factor-beta profiles correlate with clinical symptoms and parameters of haemostasis and inflammation in a controlled human malaria infection. Cytokine 2020, 125, 154838. [Google Scholar] [CrossRef]
- Walther, M.; Tongren, J.E.; Andrews, L.; Korbel, D.; King, E.; Fletcher, H.; Andersen, R.F.; Bejon, P.; Thompson, F.; Dunachie, S.J.; et al. Upregulation of TGF-beta, FOXP3, and CD4+CD25+ regulatory T cells correlates with more rapid parasite growth in human malaria infection. Immunity 2005, 23, 287–296. [Google Scholar] [CrossRef]
- Bueno, L.L.; Morais, C.G.; Araujo, F.F.; Gomes, J.A.; Correa-Oliveira, R.; Soares, I.S.; Lacerda, M.V.; Fujiwara, R.T.; Braga, E.M. Plasmodium vivax: Induction of CD4+CD25+FoxP3+ regulatory T cells during infection are directly associated with level of circulating parasites. PLoS ONE 2010, 5, e9623. [Google Scholar] [CrossRef] [PubMed]
- Lourembam, S.D.; Sawian, C.E.; Baruah, S. Dysregulation of cytokines expression in complicated falciparum malaria with increased TGF-beta and IFN-gamma and decreased IL-2 and IL-12. Cytokine 2013, 64, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Uyttenhove, C.; Marillier, R.G.; Tacchini-Cottier, F.; Charmoy, M.; Caspi, R.R.; Damsker, J.M.; Goriely, S.; Su, D.; van Damme, J.; Struyf, S.; et al. Amine-reactive OVA multimers for autovaccination against cytokines and other mediators: Perspectives illustrated for GCP-2 in L. major infection. J. Leukoc. Biol. 2011, 89, 1001–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Snick, J.; Catherine, U. Anti-TGF-β3 Patent. WO 2016/201282 A2, 15 December 2016. pp. 1–102. [Google Scholar]
- Schmitt, E.; Germann, T.; Goedert, S.; Hoehn, P.; Huels, C.; Koelsch, S.; Kühn, R.; Müller, W.; Palm, N.; Rüde, E. IL-9 production of naive CD4+ T cells depends on IL-2, is synergistically enhanced by a combination of TGF-β and IL-4 and is inhibited by IFNg. J. Immunol. 1994, 153, 3989–3996. [Google Scholar] [PubMed]
- Veldhoen, M.; Uyttenhove, C.; van Snick, J.; Helmby, H.; Westendorf, A.; Buer, J.; Martin, B.; Wilhelm, C.; Stockinger, B. Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 2008, 9, 1341–1346. [Google Scholar] [CrossRef]
- WHO. Guidelines For The Treatment of Malaria; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Day, N.P.; Hien, T.T.; Schollaardt, T.; Loc, P.P.; Chuong, L.V.; Chau, T.T.; Mai, N.T.; Phu, N.H.; Sinh, D.X.; White, N.J.; et al. The prognostic and pathophysiologic role of pro- and antiinflammatory cytokines in severe malaria. J. Infect. Dis. 1999, 180, 1288–1297. [Google Scholar] [CrossRef] [Green Version]
- Eller, K.; Wolf, D.; Huber, J.M.; Metz, M.; Mayer, G.; McKenzie, A.N.; Maurer, M.; Rosenkranz, A.R.; Wolf, A.M. IL-9 production by regulatory T cells recruits mast cells that are essential for regulatory T cell-induced immune suppression. J. Immunol. 2011, 186, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Elyaman, W.; Bradshaw, E.M.; Uyttenhove, C.; Dardalhon, V.; Awasthi, A.; Imitola, J.; Bettelli, E.; Oukka, M.; van Snick, J.; Renauld, J.C.; et al. IL-9 induces differentiation of TH17 cells and enhances function of FoxP3+ natural regulatory T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 12885–12890. [Google Scholar] [CrossRef] [Green Version]
- Richard, M.; Grencis, R.K.; Humphreys, N.E.; Renauld, J.-C.; van Snick, J. Anti-IL-9 vaccination prevents worm expulsion and blood eosinophilia in Trichuris muris-infected mice. Proc. Natl. Acad. Sci. USA 2000, 97, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Hültner, L.; Druez, C.; Moeller, J.; Uyttenhove, C.; Schmitt, E.; Rüde, E.; Dörmer, P.; van Snick, J. Mast cell growth enhancing activity (MEA) is structurally related and functionally identical to the novel mouse T cell growth factor P40/TCGFIII (interleukin 9). Eur. J. Immunol. 1990, 20, 1413–1416. [Google Scholar] [CrossRef]
- Furuta, T.; Kikuchi, T.; Iwakura, Y.; Watanabe, N. Protective roles of mast cells and mast cell-derived TNF in murine malaria. J. Immunol. 2006, 177, 3294–3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, N.O.; Bythwood, T.; Solomon, W.; Jolly, P.; Yatich, N.; Jiang, Y.; Shuaib, F.; Adjei, A.A.; Anderson, W.; Stiles, J.K. Elevated levels of IL-10 and G-CSF associated with asymptomatic malaria in pregnant women. Infect. Dis. Obstet. Gynecol. 2010, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisia, L.E.; Kempaiah, P.; Anyona, S.B.; Munde, E.O.; Achieng, A.O.; Ong’echa, J.M.; Lambert, C.G.; Chelimo, K.; Ouma, C.; Perkins, D.J.; et al. Genetic variation in interleukin-7 is associated with a reduced erythropoietic response in Kenyan children infected with Plasmodium falciparum. BMC Med. Genet. 2019, 20, 140. [Google Scholar] [CrossRef] [PubMed]
- Mazzucchelli, R.; Hixon, J.A.; Spolski, R.; Chen, X.; Li, W.Q.; Hall, V.L.; Willette-Brown, J.; Hurwitz, A.A.; Leonard, W.J.; Durum, S.K. Development of regulatory T cells requires IL-7Ralpha stimulation by IL-7 or TSLP. Blood 2008, 112, 3283–3292. [Google Scholar] [CrossRef]
- Bienkowska, A.; Kiernozek, E.; Kozlowska, E.; Bugajski, L.; Drela, N. A new approach to the role of IL-7 and TGF-ss in the in vitro generation of thymus-derived CD4+CD25+Foxp3+ regulatory T cells. Cytokine 2018, 102, 107–116. [Google Scholar] [CrossRef]
- Findlay, E.G.; Greig, R.; Stumhofer, J.S.; Hafalla, J.C.; de Souza, J.B.; Saris, C.J.; Hunter, C.A.; Riley, E.M.; Couper, K.N. Essential role for IL-27 receptor signaling in prevention of Th1-mediated immunopathology during malaria infection. J. Immunol. 2010, 185, 2482–2492. [Google Scholar] [CrossRef] [Green Version]
- Villegas-Mendez, A.; de Souza, J.B.; Lavelle, S.W.; Findlay, E.G.; Shaw, T.N.; van Rooijen, N.; Saris, C.J.; Hunter, C.A.; Riley, E.M.; Couper, K.N. IL-27 receptor signalling restricts the formation of pathogenic, terminally differentiated Th1 cells during malaria infection by repressing IL-12 dependent signals. PLoS Pathog. 2013, 9, e1003293. [Google Scholar] [CrossRef]
- Ayimba, E.; Hegewald, J.; Segbena, A.Y.; Gantin, R.G.; Lechner, C.J.; Agosssou, A.; Banla, M.; Soboslay, P.T. Proinflammatory and regulatory cytokines and chemokines in infants with uncomplicated and severe Plasmodium falciparum malaria. Clin. Exp. Immunol. 2011, 166, 218–226. [Google Scholar] [CrossRef]
- Freitas do Rosario, A.P.; Lamb, T.; Spence, P.; Stephens, R.; Lang, A.; Roers, A.; Muller, W.; O’Garra, A.; Langhorne, J. IL-27 promotes IL-10 production by effector Th1 CD4+ T cells: A critical mechanism for protection from severe immunopathology during malaria infection. J. Immunol. 2012, 188, 1178–1190. [Google Scholar] [CrossRef] [Green Version]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef]
- Bueno, L.L.; Morais, C.G.; Lacerda, M.V.; Fujiwara, R.T.; Braga, E.M. Interleukin-17 producing T helper cells are increased during natural Plasmodium vivax infection. Acta Trop. 2012, 123, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Herbert, F.; Tchitchek, N.; Bansal, D.; Jacques, J.; Pathak, S.; Becavin, C.; Fesel, C.; Dalko, E.; Cazenave, P.A.; Preda, C.; et al. Evidence of IL-17, IP-10, and IL-10 involvement in multiple-organ dysfunction and IL-17 pathway in acute renal failure associated to Plasmodium falciparum malaria. J. Transl. Med. 2015, 13, 369. [Google Scholar] [CrossRef] [PubMed]
- McLeod, J.J.; Baker, B.; Ryan, J.J. Mast cell production and response to IL-4 and IL-13. Cytokine 2015, 75, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannon, P.; Reinisch, W. Interleukin 13 and its role in gut defence and inflammation. Gut 2012, 61, 1765–1773. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, J.; Naka, I.; Patarapotikul, J.; Hananantachai, H.; Looareesuwan, S.; Tokunaga, K. A single-nucleotide substitution from C to T at position -1055 in the IL-13 promoter is associated with protection from severe malaria in Thailand. Genes Immun. 2003, 4, 528–531. [Google Scholar] [CrossRef] [Green Version]
- Devos, S.; Cormont, F.; Vrtala, S.; Hooghe-Peters, E.; Pirson, F.; Snick, J. Allergen-induced interleukin-9 production in vitro: Correlation with atopy in human adults and comparison with interleukin-5 and interleukin-13. Clin. Exp. Allergy 2006, 36, 174–182. [Google Scholar] [CrossRef]
- Silva, N.S.; Orikaza, C.M.; Santana, F.R.; dos Santos, L.A.; Salu, B.R.; Oliva, M.L.; Sinigaglia, R.D.; Mortara, R.A. Interleukin-9 in Immunopathology of Trypanosoma cruzi Experimental Infection. Front. Cell. Infect Microbiol. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- WHO. Malaria Microscopy Standard Operating Procedure 2016. World Health Organization 2016. Available online: https://apps.who.int/iris/handle/10665/274382 (accessed on 27 March 2022).
- Gaignage, M.; Marillier, R.G.; Cochez, P.M.; Dumoutier, L.; Uyttenhove, C.; Coutelier, J.P.; van Snick, J. The TLR7 ligand R848 prevents mouse graft-versus-host disease and cooperates with anti-interleukin-27 antibody for maximal protection and regulatory T-cell upregulation. Haematologica 2019, 104, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K.; Ravindran, B. Isolation of Human PBMCs. Bio-Protocol 2013, 3, e323. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmodium Species Identification | % Mild Case; n = 186 | % Severe Anaemic Malaria, n = 80 |
|---|---|---|
| Plasmodium falciparum | 82.79 | 88.75 |
| Plasmodium ovale | 6.45 | 2.5 |
| Plasmodium malariae | 6.82 | 7.5 |
| Plasmodium Vivax | 0.53 | 0 |
| Mixed | ||
| Plasmodium infections | ||
| P. falciparum and P. malariae | 1.61 | 1.25 |
| P. falciparum and P. ovale | 1.61 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndoricyimpaye, E.L.; Van Snick, J.; Niyoyita, J.d.D.; Kanimba, P.; Mbonimpa, J.B.; Rutayisire, R.; Rutayisire, R.; Ndahindwa, V.; Cheou, P.; Coutelier, J.P.; et al. Integrated Analysis of Cytokine Profiles in Malaria Patients Discloses Selective Upregulation of TGF-β1, β3, and IL-9 in Mild Clinical Presentation. Int. J. Mol. Sci. 2022, 23, 12665. https://doi.org/10.3390/ijms232012665
Ndoricyimpaye EL, Van Snick J, Niyoyita JdD, Kanimba P, Mbonimpa JB, Rutayisire R, Rutayisire R, Ndahindwa V, Cheou P, Coutelier JP, et al. Integrated Analysis of Cytokine Profiles in Malaria Patients Discloses Selective Upregulation of TGF-β1, β3, and IL-9 in Mild Clinical Presentation. International Journal of Molecular Sciences. 2022; 23(20):12665. https://doi.org/10.3390/ijms232012665
Chicago/Turabian StyleNdoricyimpaye, Ella Larissa, Jacques Van Snick, Jean de Dieu Niyoyita, Philbert Kanimba, Jean Bosco Mbonimpa, Robert Rutayisire, Réverien Rutayisire, Vedaste Ndahindwa, Paméla Cheou, Jean Paul Coutelier, and et al. 2022. "Integrated Analysis of Cytokine Profiles in Malaria Patients Discloses Selective Upregulation of TGF-β1, β3, and IL-9 in Mild Clinical Presentation" International Journal of Molecular Sciences 23, no. 20: 12665. https://doi.org/10.3390/ijms232012665
APA StyleNdoricyimpaye, E. L., Van Snick, J., Niyoyita, J. d. D., Kanimba, P., Mbonimpa, J. B., Rutayisire, R., Rutayisire, R., Ndahindwa, V., Cheou, P., Coutelier, J. P., & Rujeni, N. (2022). Integrated Analysis of Cytokine Profiles in Malaria Patients Discloses Selective Upregulation of TGF-β1, β3, and IL-9 in Mild Clinical Presentation. International Journal of Molecular Sciences, 23(20), 12665. https://doi.org/10.3390/ijms232012665