
A Non-Canonical Teleost NK-Lysin: Antimicrobial Activity via Multiple Mechanisms
Abstract
:1. Introduction
2. Results
2.1. Identification of a Non-Canonical NKL in Flounder
2.2. NKLnc Expression Is Regulated by Bacterial Pathogen and Required for Effective Blocking of Bacterial Infection
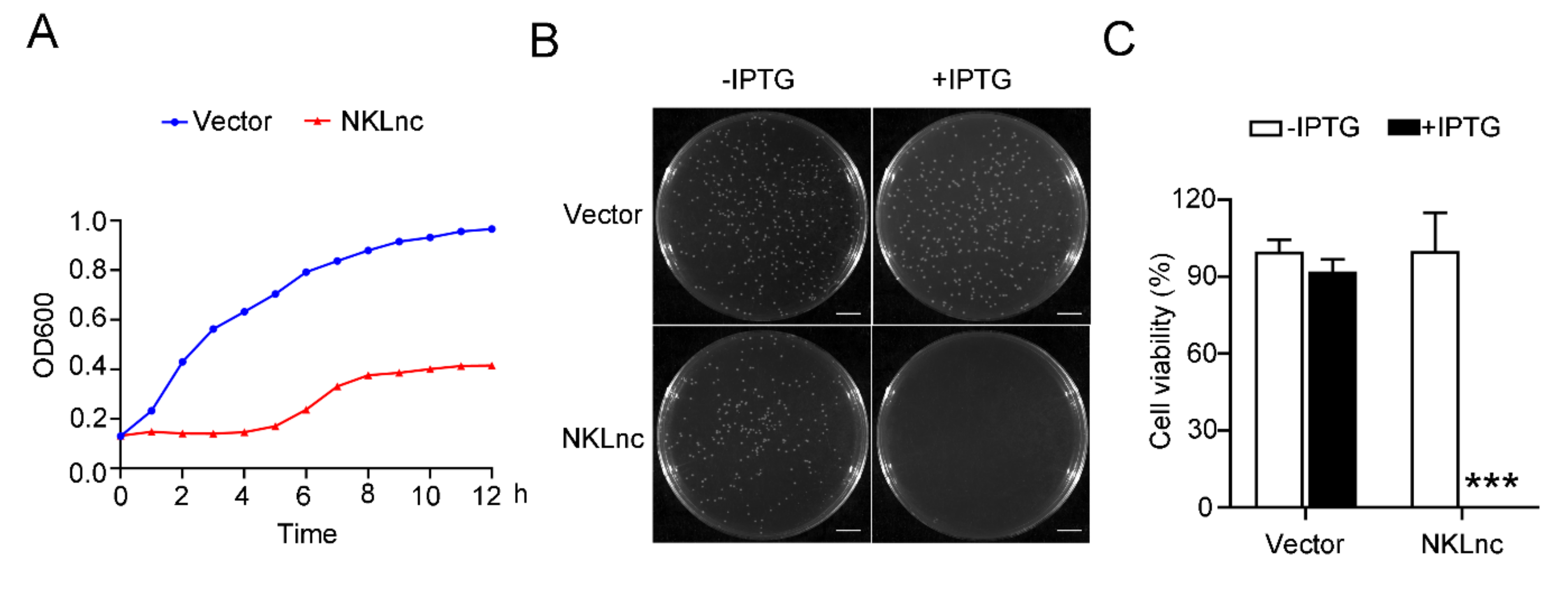
2.3. NKLnc Expression Is Detrimental to Host Bacteria
2.4. A NKLnc-Derived Peptide Is Bactericidal in a Manner That Depends on Two Alpha Helices of SapB
2.5. P35 kills Bacteria by Damaging Bacterial Plasma Membrane and Chromosomal DNA
2.6. P35 Possesses Chemotactic Activity and Promotes Bacterial Clearance from Fish Tissues
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Bacteria and Cell Line
4.3. Sequence and Structure Analyses
4.4. Quantitative Real Time PCR (qRT-PCR)
4.5. The Effect of NKLnc Expression on the Growth and Survival of Host Bacteria
4.6. Peptides
4.7. Effect of NKLnc Knockdown on Bacterial Infection in Flounder Cells
4.8. Antibacterial Activity of NKLnc Peptides
4.9. Propidium Iodide (PI) Uptake
4.10. Electron Microscopy
4.11. Effect of NKLnc on Bacterial Genomic DNA (gDNA)
4.12. Chemotaxis Assay
4.13. In Vivo Effect of P35 on Bacterial Infection and Host Survival
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Alanis, A.J. Resistance to antibiotics: Are we in the post-antibiotic era? Arch. Med. Res. 2005, 36, 697–705. [Google Scholar] [PubMed]
- Heymann, D.L. Resistance to anti-infective drugs and the threat to public health. Cell 2006, 124, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [PubMed] [Green Version]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial peptides: Key components of the innate immune system. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar]
- William, C.W. Describing the Mechanism of Antimicrobial Peptide Action with the Interfacial Activity Model. ACS Chem. Biol. 2010, 10, 905–917. [Google Scholar]
- Bustillo, M.E.; Fischer, A.L.; LaBouyer, M.A.; Klaips, J.A.; Webb, A.C.; Elmore, D.E. Modular analysis of hipposin, a histone-derived antimicrobial peptide consisting of membrane translocating and membrane permeabilizing fragments. Biochim. Biophys. Acta 2014, 1838, 2228–2233. [Google Scholar] [CrossRef] [Green Version]
- Mardirossian, M.; Grzela, R.; Giglione, C.; Meinnel, T.; Gennaro, R.; Mergaert, P.; Scocchi, M. The host antimicrobial peptide Bac71-35 binds to bacterial ribosomal proteins and inhibits protein synthesis. Chem. Biol. 2014, 21, 1639–1647. [Google Scholar]
- Haney, E.F.; Petersen, A.P.; Lau, C.K.; Jing, W.; Storey, D.G.; Vogel, H.J. Mechanism of action of puroindoline derived tryptophan-rich antimicrobial peptides. Biochim. Biophys. Acta 2013, 1828, 1802–1813. [Google Scholar]
- Guani-Guerra, E.; Santos-Mendoza, T.; Lugo-Reyes, S.O.; Teran, L.M. Antimicrobial peptides: General overview and clinical implications in human health and disease. Clin. Immunol. 2010, 135, 1–11. [Google Scholar]
- Hwang, J.S.; Lee, J.; Kim, Y.J.; Bang, H.S.; Yun, E.Y.; Kim, S.R.; Suh, H.J.; Kang, B.R.; Nam, S.H.; Jeon, J.P.; et al. Isolation and Characterization of a Defensin-Like Peptide (Coprisin) from the Dung Beetle, Copris tripartitus. Int J. Pept. 2009, 2009, 136284. [Google Scholar] [CrossRef] [PubMed]
- Rollins-Smith, L.A. The role of amphibian antimicrobial peptides in protection of amphibians from pathogens linked to global amphibian declines. Biochim. Biophys. Acta 2009, 1788, 1593–1599. [Google Scholar] [PubMed] [Green Version]
- Berrocal-Lobo, M.; Molina, A.; Rodriguez-Palenzuela, P.; Garcia-Olmedo, F.; Rivas, L. Leishmania donovani: Thionins, plant antimicrobial peptides with leishmanicidal activity. Exp. Parasitol. 2009, 122, 247–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.; Gunne, H.; Agerberth, B.; Boman, A.; Bergman, T.; Olsson, B.; Dagerlind, A.; Wigzell, H.; Boman, H.G.; Gudmundsson, G.H. Nk-Lysin, Structure and Function of a Novel Effector Molecule of Porcine T and Nk Cells. Vet. Immunol. Immunop. 1996, 54, 123–126. [Google Scholar] [CrossRef]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef] [PubMed]
- Jean-Marie, R.; Erik, G.; HombleLa, F.; Mats, A.; Edvards, L.; Gottfried, O. Lipid Membrane Binding of Nk-Lysin. FEBS Lett. 1998, 425, 341–344. [Google Scholar]
- Dotiwala, F.; Lieberman, J. Granulysin: Killer lymphocyte safeguard against microbes. Curr. Opin. Immunol. 2019, 60, 19–29. [Google Scholar] [CrossRef]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Evaluation of the Immunomodulatory Activity of the Chicken NK-Lysin-Derived Peptide cNK-2. Sci. Rep. 2017, 7, 45099. [Google Scholar] [CrossRef] [Green Version]
- Endsley, J.J.; Furrer, J.L.; Endsley, M.A.; McIntosh, M.A.; Maue, A.C.; Waters, W.R.; Lee, D.R.; Estes, D.M. Characterization of bovine homologues of granulysin and NK-lysin. J. Immunol. 2004, 173, 2607–2614. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Wang, K.; Dang, W.; Chen, R.; Xie, J.; Zhang, B.; Song, J.; Wang, R. Two hits are better than one: Membrane-active and DNA binding-related double-action mechanism of NK-18, a novel antimicrobial peptide derived from mammalian NK-lysin. Antimicrob. Agents Chemother. 2013, 57, 220–228. [Google Scholar]
- Pereiro, P.; Varela, M.; Diaz-Rosales, P.; Romero, A.; Dios, S.; Figueras, A.; Novoa, B. Zebrafish Nk-Lysins: First Insights about Their Cellular and Functional Diversification. Dev. Comp. Immunol. 2015, 51, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Z.; Yang, Y.; Guan, N.; Xia, X.; Liu, W. NKL-24: A novel antimicrobial peptide derived from zebrafish NK-lysin that inhibits bacterial growth and enhances resistance against Vibrio parahaemolyticus infection in Yesso scallop, Patinopecten yessoensis. Fish Shellfish Immunol. 2020, 106, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Long, H.; Sun, L. A NK-lysin from Cynoglossus semilaevis enhances antimicrobial defense against bacterial and viral pathogens. Dev. Comp. Immunol. 2013, 40, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Wang, M.C.; Liu, Y.L.; Zhang, Q.; Li, C.F.; Liu, P.T.; Li, E.Z.; Nie, P.; Xie, H.X. Identification, expression analysis, and antibacterial activity of NK-lysin from common carp Cyprinus carpio. Fish Shellfish Immunol. 2018, 73, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lama, R.; Pereiro, P.; Costa, M.M.; Encinar, J.A.; Medina-Gali, R.M.; Perez, L.; Lamas, J.; Leiro, J.; Figueras, A.; Novoa, B. Turbot (Scophthalmus maximus) Nk-lysin induces protection against the pathogenic parasite Philasterides dicentrarchi via membrane disruption. Fish Shellfish Immunol. 2018, 82, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, J.; Roa, F.; Gonzalez-Chavarria, I.; Astuya, A.; Maura, R.; Montesino, R.; Munoz, C.; Camacho, F.; Saavedra, P.; Valenzuela, A.; et al. In vitro immunomodulatory activities of peptides derived from Salmo salar NK-lysin and cathelicidin in fish cells. Fish Shellfish Immunol. 2019, 88, 587–594. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, Q.; Niu, J.; Tang, J.; Wang, B.; Abarike, E.D.; Lu, Y.; Cai, J.; Jian, J. NK-lysin from Oreochromis niloticus improves antimicrobial defence against bacterial pathogens. Fish Shellfish Immunol. 2018, 72, 259–265. [Google Scholar] [CrossRef]
- Ding, F.F.; Li, C.H.; Chen, J. Molecular characterization of the NK-lysin in a teleost fish, Boleophthalmus pectinirostris: Antimicrobial activity and immunomodulatory activity on monocytes/macrophages. Fish Shellfish Immunol. 2019, 92, 256–264. [Google Scholar] [CrossRef]
- Ma, H.; Han, Y.C.; Palti, Y.; Gao, G.; Liu, S.; Palmquist, D.E.; Wiens, G.D.; Shepherd, B.S. Structure and regulation of the NK-lysin (1-4) and NK-lysin like (a and b) antimicrobial genes in rainbow trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2021, 116, 103961. [Google Scholar] [CrossRef]
- Zhu, R.; Wu, Y.S.; Liu, X.X.; Lv, X.; Wu, Y.Q.; Song, J.J.; Wang, X.G. Membrane disruptive antimicrobial potential of NK-lysin from yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish Immunol. 2020, 97, 571–580. [Google Scholar] [CrossRef]
- Liu, B.; Liu, G.D.; Guo, H.Y.; Zhu, K.C.; Guo, L.; Liu, B.S.; Zhang, N.; Jiang, S.G.; Zhang, D.C. Functional characterization of NK-lysin in golden pompano Trachinotus ovatus (Linnaeus 1758). Dev. Comp. Immunol. 2020, 107, 103658. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.-J.; Wang, J.; Mao, Y.; Liu, M.; Su, Y.-Q.; Ke, Q.-Z.; Chen, J.; Zheng, W.-Q. Molecular structure, expression and antibacterial characterization of a novel antimicrobial peptide NK-lysin from the large yellow croaker Larimichthys crocea. Aquaculture 2019, 500, 315–321. [Google Scholar] [CrossRef]
- Valero, Y.; Arizcun, M.; Cortes, J.; Ramirez-Cepeda, F.; Guzman, F.; Mercado, L.; Esteban, M.A.; Chaves-Pozo, E.; Cuesta, A. NK-lysin, dicentracin and hepcidin antimicrobial peptides in European sea bass. Ontogenetic development and modulation in juveniles by nodavirus. Dev. Comp. Immunol. 2020, 103, 103516. [Google Scholar] [CrossRef]
- Hirono, I.; Kondo, H.; Koyama, T.; Arma, N.R.; Hwang, J.Y.; Nozaki, R.; Midorikawa, N.; Aoki, T. Characterization of Japanese flounder (Paralichthys olivaceus) NK-lysin, an antimicrobial peptide. Fish Shellfish Immunol. 2007, 22, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.J.; Wang, J.; Liu, M.; Qiao, Y.; Hong, W.S.; Su, Y.Q.; Han, K.H.; Ke, Q.Z.; Zheng, W.Q. Identification, expression and antibacterial activities of an antimicrobial peptide NK-lysin from a marine fish Larimichthys crocea. Fish Shellfish Immunol. 2016, 55, 195–202. [Google Scholar] [CrossRef]
- Zhang, M.; Li, M.F.; Sun, L. NKLP27: A teleost NK-lysin peptide that modulates immune response, induces degradation of bacterial DNA, and inhibits bacterial and viral infection. PLoS ONE 2014, 9, e106543. [Google Scholar] [CrossRef] [PubMed]
- Browne, K.; Chakraborty, S.; Chen, R.; Willcox, M.D.; Black, D.S.; Walsh, W.R.; Kumar, N. A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. Int. J. Mol. Sci. 2020, 21, 7047. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Bucki, R.; Leszczynska, K.; Namiot, A.; Sokolowski, W. Cathelicidin LL-37: A multitask antimicrobial peptide. Arch. Immunol. Ther. Exp. 2010, 58, 15–25. [Google Scholar] [CrossRef]
- Cheng, S.; Zhang, W.W.; Zhang, M.; Sun, L. Evaluation of the vaccine potential of a cytotoxic protease and a protective immunogen from a pathogenic Vibrio harveyi strain. Vaccine 2010, 28, 1041–1047. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Sun, Y.; Sun, L. A Crustin from Hydrothermal Vent Shrimp: Antimicrobial Activity and Mechanism. Mar. Drugs 2021, 19, 176. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Sun, Y.; Sun, L. The Establishment and Partial Characterization of a Continuous Fish Cell Line Fg-9307 from the Gill of the Flounder Paralichthys olivaceus. Aquaculture 1997, 156, 327–333. [Google Scholar]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2014, 12, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willy, W.; Klaus, S. Protein Domain Movements: Detection of Rigid Domains and Visualization of Hinges in Comparisons of Atomic Coordinates. Proteins 1997, 29, 1–14. [Google Scholar]
- Yu, C.; Zhang, P.; Li, X.-p.; Sun, L. Japanese flounder Paralichthys olivaceus interleukin 21 induces inflammatory response and plays a vital role in the immune defense against bacterial pathogen. Fish Shellfish Immunol. 2020, 98, 364–373. [Google Scholar] [CrossRef]
- Zheng, W.J.; Sun, L. Evaluation of housekeeping genes as references for quantitative real time RT-PCR analysis of gene expression in Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2011, 30, 638–645. [Google Scholar] [CrossRef]
- Li, H.; Sun, B.; Jiang, S.; Sun, L. Cytokines Induced by Edwardsiella tarda: Profile and Role in Antibacterial Immunity. Biomolecules 2021, 11, 1242. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, S.; Sun, L. A fish galectin-8 possesses direct bactericidal activity. Int. J. Mol. Sci. 2020, 22, 376. [Google Scholar] [CrossRef]
- Gu, H.J.; Sun, Q.L.; Jiang, S.; Zhang, J.; Sun, L. First characterization of an anti-lipopolysaccharide factor (ALF) from hydrothermal vent shrimp: Insights into the immune function of deep-sea crustacean ALF. Dev. Comp. Immunol. 2018, 84, 382–395. [Google Scholar] [CrossRef]
- Li, H.; Sun, Y.; Sun, L. A Teleost CXCL10 Is Both an Immunoregulator and an Antimicrobial. Front. Immunol. 2022, 13, 917697. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, S.; Yu, C.; Yuan, Z.; Sun, L. GSDMEa-mediated pyroptosis is bi-directionally regulated by caspase and required for effective bacterial clearance in teleost. Cell Death Dis. 2022, 13, 491. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | P35 MIC (μM) | P17 MIC (μM) | Strains | P35 MIC (μM) | P17 MIC (μM) |
|---|---|---|---|---|---|
| Gram-positive | Gram-negative | ||||
| Bacillus subtilis | 5 | — | Edwardsiella tarda | 40 | — |
| Bacillus cereus | — | — | Escherichia coli | 10 | 40 |
| Micrococcus luteus | 10 | 40 | Pseudomona fluorescens | 20 | — |
| Staphylococcus aureus | 10 | 40 | Vibrio anguillarum | 5 | 20 |
| Streptococcus iniae | 10 | 40 | Vibrio harveyi | 10 | 40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Yuan, Z.; Sun, L. A Non-Canonical Teleost NK-Lysin: Antimicrobial Activity via Multiple Mechanisms. Int. J. Mol. Sci. 2022, 23, 12722. https://doi.org/10.3390/ijms232112722
Xu H, Yuan Z, Sun L. A Non-Canonical Teleost NK-Lysin: Antimicrobial Activity via Multiple Mechanisms. International Journal of Molecular Sciences. 2022; 23(21):12722. https://doi.org/10.3390/ijms232112722
Chicago/Turabian StyleXu, Hang, Zihao Yuan, and Li Sun. 2022. "A Non-Canonical Teleost NK-Lysin: Antimicrobial Activity via Multiple Mechanisms" International Journal of Molecular Sciences 23, no. 21: 12722. https://doi.org/10.3390/ijms232112722
APA StyleXu, H., Yuan, Z., & Sun, L. (2022). A Non-Canonical Teleost NK-Lysin: Antimicrobial Activity via Multiple Mechanisms. International Journal of Molecular Sciences, 23(21), 12722. https://doi.org/10.3390/ijms232112722