
The Role of Telomerase in Breast Cancer’s Response to Therapy



{kind=link}
Abstract
1. Introduction
2. Breast Cancer—A Challenge
3. Obstacles in Therapeutic Strategies
4. Immortality of Cancer Cells
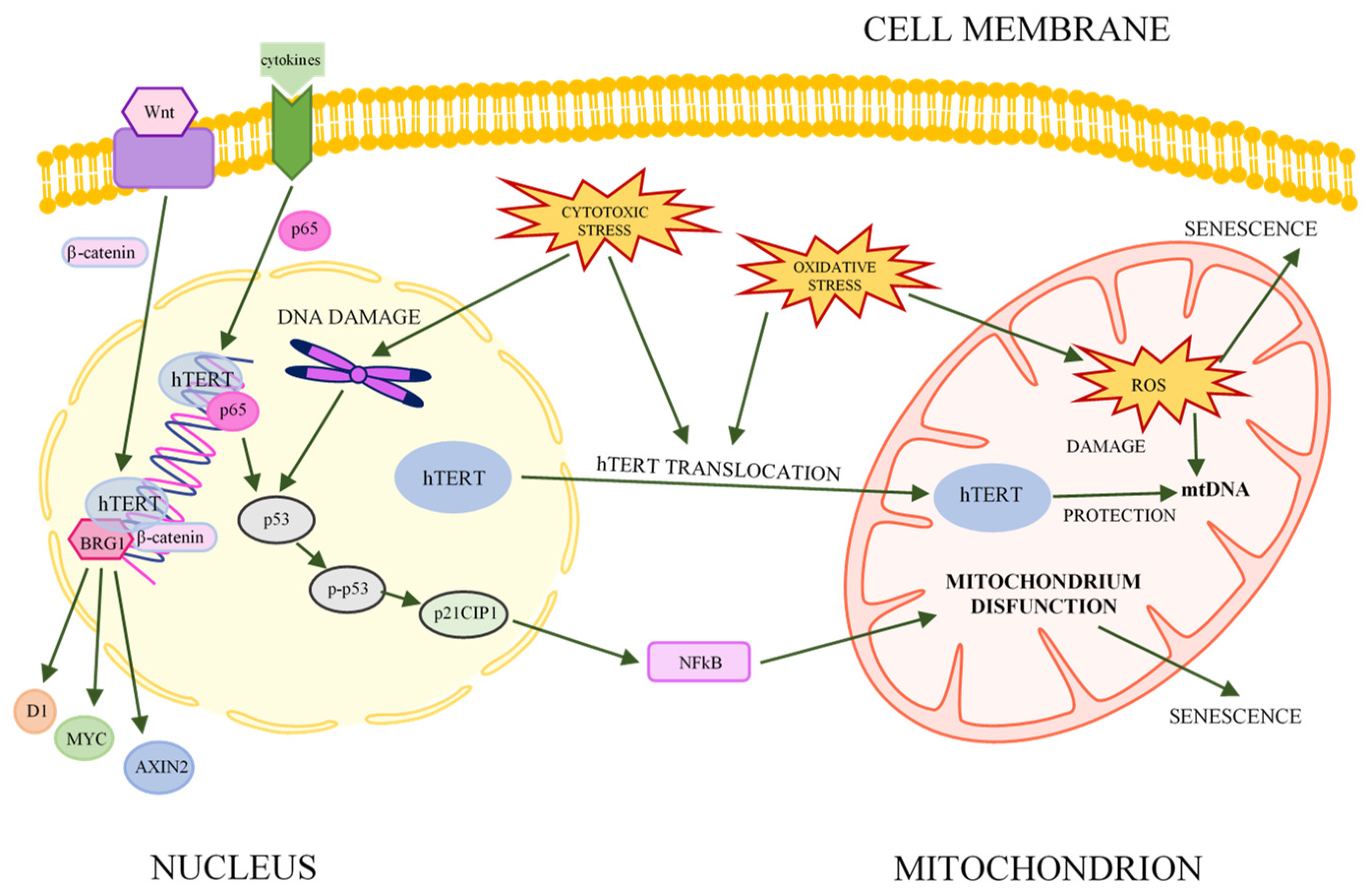
5. Telomerase—Key Factor in Immortality
6. Canonical and Noncanonical Functions of Telomerase
7. Therapeutic Potential of Telomerase Subunits
7.1. Telomerase as a Target for Cancer Therapy
7.2. Targeting Telomeres
7.3. Genetic Profiling in Different Cancers
7.4. Immunotherapy
7.5. New Generation Vaccines
8. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- GLOBOCAN 2020. Estimated Number of Incident Cases and Deaths Worldwide, Females, All Ages (excl. NMSC). Available online: https://gco.iarc.fr/today/online-analysis-multi-bars?v=2020&mode=cancer&mode_population=countries&population=900&populations=&key=total&sex=2&cancer=39&type=0&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&nb_items=10&group_cancer=1&include_nmsc=0&include_nmsc_other=1&type_multiple=%257B%2522inc%2522%253Atrue%252C%2522mort%2522%253Atrue%252C%2522prev%2522%253Afalse%257D&orientation=horizontal&type_sort=0&type_nb_items=%257B%2522top%2522%253Atrue%252C%2522bottom%2522%253Afalse%257D (accessed on 14 April 2022).
- Bryś, M.; Laskowska, M.; Forma, E.; Krześlak, A. Telomeraza—Struktura i Funkcja Oraz Regulacja Ekspresji Genu. Folia Med. Lodz. 2012, 39, 293–326. [Google Scholar]
- Xu, Y.; Goldkorn, A. Telomere and Telomerase Therapeutics in Cancer. Genes 2016, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Krzakowski, K.; Krzemieniecki, K. Wybrane Nowotwory: Rak Piersi. In Interna Szczeklika; Gajewski, P., Szczeklik, A., Eds.; Medycyna Praktyczna: Kraków, Poland, 2017; pp. 2325–2329. [Google Scholar]
- Balcerak, A.; Wakuła, M.; Trȩbińska, A.; Grzybowska, E.A. Migracja i Inwazyjność Komórek Nowotworowych; Rola Plastyczności Komórek i Udział Macierzy Zewnątrzkomórkowej w Tworzeniu Przerzutów. Nowotwory 2016, 66, 45–52. [Google Scholar] [CrossRef][Green Version]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-Mesenchymal Transition in Cancer: A Historical Overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, C.; Conceição, F.; Jerónimo, C.; Lamghari, M.; Sousa, D.M. Stress in Metastatic Breast Cancer: To the Bone and Beyond. Cancers 2022, 14, 1881. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, S.; Wang, X. The Metabolic Mechanisms of Breast Cancer Metastasis. Front. Oncol. 2020, 10, 602416. [Google Scholar] [CrossRef]
- Blanco, M.A.; Kang, Y. Signaling pathways in breast cancer metastasis—Novel insights from functional genomics. Breast Cancer Res. 2011, 13, 206. [Google Scholar] [CrossRef]
- Gupta, G.P.; Nguyen, D.X.; Chiang, A.C.; Bos, P.D.; Kim, J.Y.; Nadal, C.; Gomis, R.R.; Manova-Todorova, K.; Massagué, J. Mediators of vascular remodelling co-opted for sequential steps in lung metastasis. Nature 2007, 446, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef]
- Romaniuk-Drapała, A.; Totoń, E.; Konieczna, N.; Machnik, M.; Barczak, W.; Kowal, D.; Kopczyński, P.; Kaczmarek, M.; Rubiś, B. hTERT Downregulation Attenuates Resistance to DOX, Impairs FAK-Mediated Adhesion, and Leads to Autophagy Induction in Breast Cancer Cells. Cells 2021, 10, 867. [Google Scholar] [CrossRef]
- Sardanelli, F.; Aase, H.S.; Álvarez, M.; Azavedo, E.; Baarslag, H.J.; Balleyguier, C.; Baltzer, P.A.; Beslagic, V.; Bick, U.; Bogdanovic-Stojanovic, D.; et al. Position Paper on Screening for Breast Cancer by the European Society of Breast Imaging (EUSOBI) and 30 National Breast Radiology Bodies from Austria, Belgium, Bosnia and Herzegovina, Bulgaria, Croatia, Czech Republic, Denmark, Estonia, Finland, France, Germany, Greece, Hungary, Iceland, Ireland, Italy, Israel, Lithuania, Moldova, The Netherlands, Norway, Poland, Portugal, Romania, Serbia, Slovakia, Spain, Sweden, Switzerland and Turkey. Eur. Radiol. 2017, 27, 2737–2743. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Tabriz Univ. Med. Sci. 2017, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Totoń, E.; Jacczak, B.; Barczak, W.; Jagielski, P.; Gryczka, R.; Hołysz, H.; Grodecka-Gazdecka, S.; Rubiś, B. No Association between ABCB1 G2677T/A or C3435T Polymorphisms and Survival of Breast Cancer Patients-A 10-Year Follow-Up Study in the Polish Population. Genes 2022, 13, 729. [Google Scholar] [CrossRef]
- Siedlecki, J.A. Biologia Molekularna Nowotworów. In Podstawy Onkologii Klinicznej; Meder, J., Ed.; Centrum Medyczne Kształcenia Podyplomowego w Warszawie: Warszawa, Poland, 2011; Volume 6, pp. 41–46. [Google Scholar]
- Ścibior-Bentkowska, D.; Czeczot, H. Komórki Nowotworowe a Stres Oksydacyjny Cancer Cells and Oxidative Stress. Postep. Hig. Med. Dosw. 2009, 63, 58–72. [Google Scholar]
- Olszewski, W. Podstawy Patologii Nowotworów. In Podstawy Onkologii Klinicznej; Meder, J., Ed.; Centrum Medyczne Kształcenia Podyplomowego w Warszawie: Warszawa, Poland, 2011; Volume 5, pp. 29–40. [Google Scholar]
- Dominiak, A.; Nowicki, T.; Lacheta, D.; Nowicka, G. Cancer-Associated Fibroblasts as a Potential Target in Oncology Therapy. In Postepy Higieny i Medycyny Doswiadczalnej; Polska Akademia Nauk: Warszawa, Poland, 2019; pp. 536–548. [Google Scholar] [CrossRef]
- Romaniuk, A.; Paszel-Jaworska, A.; Totoń, E.; Lisiak, N.; Hołysz, H.; Królak, A.; Grodecka-Gazdecka, S.; Rubiś, B. The Non-Canonical Functions of Telomerase: To Turn Off or Not to Turn Off. Mol. Biol. Rep. 2019, 46, 1401–1411. [Google Scholar] [CrossRef]
- Lipinska, N.; Romaniuk, A.; Paszel-Jaworska, A.; Toton, E.; Kopczynski, P.; Rubis, B. Telomerase and Drug Resistance in Cancer. Cell. Mol. Life Sci. 2017, 74, 4121–4132. [Google Scholar] [CrossRef]
- Blackburn, E.H. Telomeres and Telomerase: Their Mechanisms of Action and the Effects of Altering Their Functions. FEBS Lett. 2005, 579, 859–862. [Google Scholar] [CrossRef]
- Smoleń, E.; Dobrowolska, B. Praktyka Samobadania Piersi i Wykonywanie Mammografii w Grupie pielęgniareka Zmienne Socjodemograficzne. Med. Sr. -Environ. Med. 2017, 20, 56–65. [Google Scholar] [CrossRef]
- Park, J.-I.; Venteicher, A.S.; Yeon Hong, J.; Choi, J.; Jun, S.; Shkreli, M.; Chang, W.; Meng, Z.; Cheung, P.; Ji, H.; et al. Telomerase Modulates Wnt Signalling by Association with Target Gene Chromatin. Nature 2009, 460, 66–72. [Google Scholar] [CrossRef]
- Moreno, I.; Alborán, D.; Hartl, M.A.N.; Flores, I.; Khattar, E.; Tergaonkar, V. Transcriptional Regulation of Telomerase Reverse Transcriptase (TERT) by MYC. Front. Cell Dev. Biol. 2017, 5, 1. [Google Scholar] [CrossRef]
- Koh, C.M.; Khattar, E.; Leow, S.C.; Liu, C.Y.; Muller, J.; Ang, W.X.; Li, Y.; Franzoso, G.; Li, S.; Guccione, E.; et al. Telomerase Regulates MYC-Driven Oncogenesis Independent of Its Reverse Transcriptase Activity. J. Clin. Investig. 2015, 125, 2109–2122. [Google Scholar] [CrossRef] [PubMed]
- Akincilar, S.C.; Chan, C.H.T.; Ng, Q.F.; Fidan, K.; Tergaonkar, V. Non-Canonical Roles of Canonical Telomere Binding Proteins in Cancers. Cell. Mol. Life Sci. 2021, 78, 4235–4257. [Google Scholar] [CrossRef] [PubMed]
- Aklncllar, S.C.; Wu, L.; Ng, Q.F.; Chua, J.Y.H.; Unal, B.; Noda, T.; Chor, W.H.J.; Ikawa, M.; Tergaonkar, V. NAIL: An Evolutionarily Conserved LncRNA Essential for Licensing Coordinated Activation of P38 and NFκB in Colitis. Gut 2021, 70, 1857–1871. [Google Scholar] [CrossRef]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-ΚB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Gaynor, R.B. IκB Kinases: Key Regulators of the NF-ΚB Pathway. Trends Biochem. Sci. 2004, 29, 72–79. [Google Scholar] [CrossRef]
- Ghosh, A.; Saginc, G.; Leow, S.C.; Khattar, E.; Shin, E.M.; Yan, T.D.; Wong, M.; Zhang, Z.; Li, G.; Sung, W.K.; et al. Telomerase Directly Regulates NF-B-Dependent Transcription. Nat. Cell Biol. 2012, 14, 1270–1281. [Google Scholar] [CrossRef]
- Ahmed, S.; Passos, J.F.; Birket, M.J.; Beckmann, T.; Brings, S.; Peters, H.; Birch-Machin, M.A.; von Zglinicki, T.; Saretzki, G. Telomerase Does Not Counteract Telomere Shortening but Protects Mitochondrial Function under Oxidative Stress. J. Cell Sci. 2008, 121, 1046–1053. [Google Scholar] [CrossRef]
- Indran, I.R.; Hande, M.P.; Pervaiz, S. HTERT Overexpression Alleviates Intracellular ROS Production, Improves Mitochondrial Function, and Inhibits ROS-Mediated Apoptosis in Cancer Cells. Cancer Res. 2011, 71, 266–276. [Google Scholar] [CrossRef]
- Ségal-Bendirdjian, E.; Geli, V. Non-Canonical Roles of Telomerase: Unraveling the Imbroglio. Front. Cell Dev. Biol. 2019, 7, 332. [Google Scholar] [CrossRef]
- Gordon, D.M.; Santos, J.H. The Emerging Role of Telomerase Reverse Transcriptase in Mitochondrial DNA Metabolism. J. Nucleic Acids 2010, 2010, 390791. [Google Scholar] [CrossRef] [PubMed]
- Miao, G.-Y.; Zhou, X.; Zhang, X.; Xie, Y.; Sun, C.; Liu, Y.; Gan, L.; Zhang, H. Telomere-Mitochondrion Links Contribute to Induction of Senescence in MCF-7 Cells after Carbon-Ion Irradiation. Asian Pac. J. Cancer Prev. 2016, 17, 1993–1998. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhou, Y.; Chen, D.; Li, L.; Yang, X.; You, Y.; Ling, X. Impact of Mitochondrial Telomerase Over-Expression on Drug Resistance of Hepatocellular Carcinoma. Am. J. Transl. Res. 2015, 7, 88–99. [Google Scholar] [PubMed]
- Jacczak, B.; Rubiś, B.; Totoń, E. Potential of Naturally Derived Compounds in Telomerase and Telomere Modulation in Skin Senescence and Aging. Int. J. Mol. Sci. 2021, 22, 6381. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhou, Y.; Chen, D.X.; Li, L.L.; Yang, X.; You, Y.; Ling, X. Effects of Mitochondrial Translocation of Telomerase on Drug Resistance in Hepatocellular Carcinoma Cells. J. Cancer 2015, 6, 151. [Google Scholar] [CrossRef]
- Ling, X.; Wen, L.; Zhou, Y. Role of Mitochondrial Translocation of Telomerase in Hepatocellular Car-Cinoma Cells with Multidrug Resistance. Int. J. Med. Sci. 2012, 9, 545–554. [Google Scholar] [CrossRef][Green Version]
- Bagheri, S.; Nosrati, M.; Li, S.; Fong, S.; Torabian, S.; Rangel, J.; Moore, D.H.; Federman, S.; LaPosa, R.R.; Baehner, F.L.; et al. Genes and Pathways Downstream of Telomerase in Melanoma Metastasis. Proc. Natl. Acad. Sci. USA 2006, 103, 11306–11311. [Google Scholar] [CrossRef]
- Li, S.; Crothers, J.; Haqq, C.M.; Blackburn, E.H. Cellular and Gene Expression Responses Involved in the Rapid Growth Inhibition of Human Cancer Cells by RNA Interference-Mediated Depletion of Telomerase RNA. J. Biol. Chem. 2005, 280, 23709–23717. [Google Scholar] [CrossRef]
- Liu, H.; Yang, Y.; Ge, Y.; Liu, J.; Zhao, Y. TERC Promotes Cellular Inflammatory Response Independent of Telomerase. Nucleic Acids Res. 2019, 47, 8084–8095. [Google Scholar] [CrossRef]
- Ivancich, M.; Schrank, Z.; Wojdyla, L.; Leviskas, B.; Kuckovic, A.; Sanjali, A.; Puri, N. Treating Cancer by Targeting Telomeres and Telomerase. Antioxidants 2017, 6, 15. [Google Scholar] [CrossRef]
- Broccoli, D.; Young, J.W.; De Lange, T. Telomerase Activity in Normal and Malignant Hematopoietic Cells. Proc. Natl. Acad. Sci. USA 1995, 92, 9082–9086. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, R.D.; Wright, W.E.; Shay, J.W.; Taylor, R.S. Telomerase Activity Concentrates in the Mitotically Active Segments of Human Hair Follicles. J. Investig. Dermatol. 1997, 108, 113–117. [Google Scholar] [CrossRef]
- Shay, J.W.; Bacchetti, S. A Survey of Telomerase Activity in Human Cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Wright, W.E.; Piatyszek, M.A.; Rainey, W.E.; Byrd, W.; Shay, J.W. Telomerase Activity in Human Germline and Embryonic Tissues and Cells. Dev. Genet. 1996, 18, 173–179. [Google Scholar] [CrossRef]
- Kazanowska, B.; Mikołajewska, A.; Reich, A.; Reich, M.; Chybicka, A. Telomery i Aktywność Telomerazy w Komórkach Prawidłowych Oraz w Komórkach Nowotworowych. Adv. Clin. Exp. Med. 2003, 12, 87–95. [Google Scholar]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of Telomeres and Telomerase in Cancer, and Advances in Telomerase-Targeted Therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef]
- Gruber, H.-J.; Semeraro, M.D.; Renner, W.; Herrmann, M. Telomeres and Age-Related Diseases. Biomedicines 2021, 9, 1355. [Google Scholar] [CrossRef]
- Gridelli, C.; Ciuleanu, T.; Domine, M.; Szczesna, A.; Bover, I.; Cobo, M.; Kentepozidis, N.; Zarogoulidis, K.; Kalofonos, C.; Kazarnowisz, A.; et al. Clinical Activity of a Htert (vx-001) Cancer Vaccine as Post-Chemotherapy Maintenance Immunotherapy in Patients with Stage IV Non-Small Cell Lung Cancer: Final Results of a Randomised Phase 2 Clinical Trial. Br. J. Cancer 2020, 122, 1461–1466. [Google Scholar] [CrossRef]
- Relitti, N.; Saraswati, A.P.; Federico, S.; Khan, T.; Brindisi, M.; Zisterer, D.; Brogi, S.; Gemma, S.; Butini, S.; Campiani, G. Telomerase-Based Cancer Therapeutics: A Review on Their Clinical Trials. Curr. Top. Med. Chem. 2020, 20, 433–457. [Google Scholar] [CrossRef]
- Wang, L.; Li, P.F.; Geng, M.; Cao, Y.C.; Yin, Y.C. Correlation between Chemosensitivity to Anticancer Drugs and Telomerase Reverse Transcriptase MRNA Expression in Gastric Cancer. Diagn. Pathol. 2013, 8, 33. [Google Scholar] [CrossRef]
- Poynter, K.R.; Sachs, P.C.; Bright, A.T.; Breed, M.S.; Nguyen, B.N.; Elmore, L.W.; Holt, S.E. Genetic Inhibition of Telomerase Results in Sensitization and Recovery of Breast Tumor Cells. Mol. Cancer Ther. 2009, 8, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Massard, C.; Zermati, Y.; Pauleau, A.L.; Larochette, N.; Métivier, D.; Sabatier, L.; Kroemer, G.; Soria, J.C. HTERT: A Novel Endogenous Inhibitor of the Mitochondrial Cell Death Pathway. Oncogene 2006, 25, 4505–4514. [Google Scholar] [CrossRef]
- Gellert, G.C.; Dikmen, Z.G.; Wright, W.E.; Gryaznov, S.; Shay, J.W. Effects of a Novel Telomerase Inhibitor, GRN163L, in Human Breast Cancer. Breast Cancer Res. Treat. 2006, 96, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Hochreiter, A.E.; Xiao, H.; Goldblatt, E.M.; Gryaznov, S.M.; Miller, K.D.; Badve, S.; Sledge, G.W.; Herbert, B.S. Telomerase Template Antagonist GRN163L Disrupts Telomere Maintenance, Tumor Growth, and Metastasis of Breast Cancer. Clin. Cancer Res. 2006, 12, 3184–3192. [Google Scholar] [CrossRef]
- Koziel, J.E.; Herbert, B.S. The telomerase inhibitor imetelstat alone, and in combination with trastuzumab, decreases the cancer stem cell population and self-renewal of HER2+ breast cancer cells. Breast Cancer Res. Treat. 2015, 149, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qian, X.; Wang, B.; Xia, Y.; Zheng, Y.; Du, L.; Xu, D.; Xing, D.; DePinho, R.A.; Lu, Z. Programmable base editing of mutated TERT promoter inhibits brain tumor growth. Nat. Cell Biol. 2020, 22, 282–288. [Google Scholar] [CrossRef]
- Bochman, M.L.; Paeschke, K.; Zakian, V.A. DNA Secondary Structures: Stability and Function of G-Quadruplex Structures. Nat. Rev. Genet. 2012, 13, 770–780. [Google Scholar] [CrossRef]
- Moye, A.L.; Porter, K.C.; Cohen, S.B.; Phan, T.; Zyner, K.G.; Sasaki, N.; Lovrecz, G.O.; Beck, J.L.; Bryan, T.M. Telomeric G-Quadruplexes Are a Substrate and Site of Localization for Human Telomerase. Nat. Commun. 2015, 6, 7643. [Google Scholar] [CrossRef]
- Riou, J.F.; Guittat, L.; Mailliet, P.; Laoui, A.; Renou, E.; Petitgenet, O.; Mégnin-Chanet, F.; Hélène, C.; Mergny, J.L. Cell Senescence and Telomere Shortening Induced by a New Series of Specific G-Quadruplex DNA Ligands. Proc. Natl. Acad. Sci. USA 2002, 99, 2672–2677. [Google Scholar] [CrossRef]
- Amato, R.; Valenzuela, M.; Berardinelli, F.; Salvati, E.; Maresca, C.; Leone, S.; Antoccia, A.; Sgura, A. G-Quadruplex Stabilization Fuels the ALT Pathway in ALT-Positive Osteosarcoma Cells. Genes 2020, 11, 304. [Google Scholar] [CrossRef]
- Liu, Y.C.; Yang, D.Y.; Sheu, S.Y. Insights into the Free Energy Landscape and Salt-Controlled Mechanism of the Conformational Conversions between Human Telomeric G-Quadruplex Structures. Int. J. Biol. Macromol. 2021, 191, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Barczak, W.; Suchorska, W.M.; Sobecka, A.; Bednarowicz, K.; Machczynski, P.; Golusinski, P.; Rubis, B.; Masternak, M.M.; Golusinski, W. HTERT C250T Promoter Mutation and Telomere Length as a Molecular Markers of Cancer Progression in Patients with Head and Neck Cancer. Mol. Med. Rep. 2017, 16, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Hilton, J.; Gelmon, K.; Bedard, P.L.; Tu, D.; Xu, H.; Tinker, A.V.; Goodwin, R.; Laurie, S.A.; Jonker, D.; Hansen, A.R.; et al. Results of the Phase I CCTG IND.231 Trial of CX-5461 in Patients with Advanced Solid Tumors Enriched for DNA-Repair Deficiencies. Nat. Commun. 2022, 13, 3607. [Google Scholar] [CrossRef]
- Kulak, O.; Chen, H.; Holohan, B.; Wu, X.; He, H.; Borek, D.; Otwinowski, Z.; Yamaguchi, K.; Garofalo, L.A.; Ma, Z.; et al. Disruption of Wnt/β-Catenin Signaling and Telomeric Shortening Are Inextricable Consequences of Tankyrase Inhibition in Human Cells. Mol. Cell. Biol. 2015, 35, 2425–2435. [Google Scholar] [CrossRef]
- Marko Ivancich, L.W. Targeting the Telomere with T-Oligo, G-Quadruplex Stabilizers, and Tankyrase Inhibitors. J. Cancer Sci. Ther. 2014, 6, 429–432. [Google Scholar] [CrossRef]
- Seimiya, H. Crossroads of Telomere Biology and Anticancer Drug Discovery. Cancer Sci. 2020, 111, 3089–3099. [Google Scholar] [CrossRef]
- Yu, M.; Yang, Y.; Sykes, M.; Wang, S. Small-Molecule Inhibitors of Tankyrases as Prospective Therapeutics for Cancer. J. Med. Chem. 2022, 65, 5244–5273. [Google Scholar] [CrossRef]
- Schrank, Z.; Khan, N.; Osude, C.; Singh, S.; Miller, R.J.; Merrick, C.; Mabel, A.; Kuckovic, A.; Puri, N. Oligonucleotides Targeting Telomeres and Telomerase in Cancer. Molecules 2018, 23, 2267. [Google Scholar] [CrossRef]
- Chen, X.; Tang, W.J.; Shi, J.B.; Liu, M.M.; Liu, X.H. Therapeutic Strategies for Targeting Telomerase in Cancer. Med. Res. Rev. 2020, 40, 532–585. [Google Scholar] [CrossRef]
- Chhabra, G.; Wojdyla, L.; Frakes, M.; Schrank, Z.; Leviskas, B.; Ivancich, M.; Vinay, P.; Ganapathy, R.; Ramirez, B.E.; Puri, N. Mechanism of Action of G-Quadruplex–Forming Oligonucleotide Homologous to the Telomere Overhang in Melanoma. J. Investig. Dermatol. 2018, 138, 903–910. [Google Scholar] [CrossRef]
- Puri, N.; Pitman, R.T.; Mulnix, R.E.; Erickson, T.; Iness, A.N.; Vitali, C.; Zhao, Y.; Salgia, R. Non-Small Cell Lung Cancer Is Susceptible to Induction of DNA Damage Responses and Inhibition of Angiogenesis by Telomere Overhang Oligonucleotides. Cancer Lett. 2014, 343, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Eller, M.S.; Panova, I.; Kubera, J.; Wee, L.H.; Cowan, K.H.; Gilchrest, B.A. Telomeric DNA Induces Apoptosis and Senescence of Human Breast Carcinoma Cells. Breast Cancer Res. 2007, 9, R13. [Google Scholar] [CrossRef] [PubMed]
- Puri, N.; Eller, M.S.; Byers, H.R.; Dykstra, S.; Kubera, J.; Gilchrest, B.A. Telomere-based DNA Damage Responses: A New Approach to Melanoma. FASEB J. 2004, 18, 1373–1381. [Google Scholar] [CrossRef]
- Li, G.Z.; Eller, M.S.; Hanna, K.; Gilchrest, B.A. Signaling Pathway Requirements for Induction of Senescence by Telomere Homolog Oligonucleotides. Exp. Cell Res. 2004, 301, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Jue, M.; Gregory, X.; Chinand, L.; Garraway, L.A. Highly Recurrent TERT Promoter Mutations in Human Melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef]
- Killela, P.J.; Reitman, Z.J.; Jiao, Y.; Bettegowda, C.; Agrawal, N.; Diaz, L.A., Jr.; Friedman, A.H.; Friedman, H.; Gallia, G.L.; Giovanella, B.C.; et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. USA 2013, 110, 6021–6026. [Google Scholar] [CrossRef]
- Fredriksson, N.J.; Ny, L.; Nilsson, J.A.; Larsson, E. Systematic analysis of noncoding somatic mutations and gene expression alterations across 14 tumor types. Nat. Genet. 2014, 46, 1258–1263. [Google Scholar] [CrossRef]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef]
- Sieverling, L.; Hong, C.; Koser, S.D.; Ginsbach, P.; Kleinheinz, K.; Hutter, B.; Braun, D.M.; Cortés-Ciriano, I.; Xi, R.; Kabbe, R.; et al. Genomic footprints of activated telomere maintenance mechanisms in cancer. Nat. Commun. 2020, 11, 733. [Google Scholar] [CrossRef]
- Noureen, N.; Wu, S.; Lv, Y.; Yang, J.; Yung, W.K.A.; Gelfond, J.; Wang, X.; Koul, D.; Ludlow, A.; Zheng, S. Integrated analysis of telomerase enzymatic activity unravels an association with cancer stemness and proliferation. Nat. Commun. 2021, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Akincilar, S.C.; Low, K.C.; Liu, C.Y.; Yan, T.D.; Oji, A.; Ikawa, M.; Li, S.; Tergaonkar, V. Quantitative Assessment of Telomerase Components in Cancer Cell Lines. FEBS Lett. 2015, 589, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Leão, R.; Apolónio, J.D.; Lee, D.; Figueiredo, A.; Tabori, U.; Castelo-Branco, P. Mechanisms of human telomerase reverse transcriptase (hTERT) regulation: Clinical impacts in cancer. J. Biomed. Sci. 2018, 25, 22. [Google Scholar] [CrossRef]
- Eller, M.S.; Li, G.; Firoozabadi, R.; Puri, N.; Gilchrest, B.A. Induction of a P95/Nbs1-mediated S Phase Checkpoint by Telomere 3′ Overhang Specific DNA. FASEB J. 2003, 17, 152–162. [Google Scholar] [CrossRef]
- Greten, T.F.; Forner, A.; Korangy, F.; N’Kontchou, G.; Barget, N.; Ayuso, C.; Ormandy, L.A.; Manns, M.P.; Beaugrand, M.; Bruix, J. A Phase II Open Label Trial Evaluating Safety and Efficacy of a Telomerase Peptide Vaccination in Patients with Advanced Hepatocellular Carcinoma. BMC Cancer 2010, 10, 209. [Google Scholar] [CrossRef] [PubMed]
- Mizukoshi, E.; Kaneko, S. Telomerase-Targeted Cancer Immunotherapy. Int. J. Mol. Sci. 2019, 20, 1823. [Google Scholar] [CrossRef]
- Brower, V. Telomerase-Based Therapies Emerging Slowly. J. Natl. Cancer Inst. 2010, 102, 520–521. [Google Scholar] [CrossRef]
- Kailashiya, C.; Sharma, H.B.; Kailashiya, J. Telomerase Based Anticancer Immunotherapy and Vaccines Approaches. Vaccine 2017, 35, 5768–5775. [Google Scholar] [CrossRef]
- DiPersio, J.F.; Collins, R.H.; Blum, W.; Devetten, M.P.; Stiff, P.; Elias, L.; Reddy, A.; Smith, J.A.; Khoury, H.J. Immune Responses in AML Patients Following Vaccination with GRNVAC1, Autologous RNA Transfected Dendritic Cells Expressing Telomerase Catalytic Subunit HTERT. Blood 2009, 114, 633. [Google Scholar] [CrossRef]
- Negrini, S.; De Palma, R.; Filaci, G. Anti-Cancer Immunotherapies Targeting Telomerase. Cancers 2020, 12, 2260. [Google Scholar] [CrossRef]
- Yamano, T.; Kaneda, Y.; Hiramatsu, S.H.; Huang, S.; Tran, A.N.; Giuliano, A.E.; Hoon, D.S.B. Immunity against breast cancer by TERT DNA vaccine primed with chemokine CCL21. Cancer Gene Ther. 2007, 14, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Ellingsen, E.B.; Aamdal, E.; Guren, T.; Lilleby, W.; Brunsvig, P.F.; Mangsbo, S.M.; Steinar, A.; Hovig, E.; Mensali, N.; Gaudernack, G.; et al. Durable and dynamic hTERT immune responses following vaccination with the long-peptide cancer vaccine UV1: Long-term follow-up of three phase I clinical trials. J. Immunother. Cancer 2022, 10, e004345. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Medioni, J.; Garibal, J.; Adotevi, O.; Doucet, L.; Durey, M.D.; Ghrieb, Z.; Kiladjian, J.J.; Brizard, M.; Laheurte, C.; et al. A First-in-Human Phase I Study of INVAC-1, an Optimized Human Telomerase DNA Vaccine in Patients with Advanced Solid Tumors. Clin. Cancer Res. 2020, 26, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://clinicaltrials.gov (accessed on 10 October 2022).
- Pulaski, B.A.; Ostrand-Rosenberg, S. Mouse 4T1 breast tumor model. Curr. Protoc. Immunol. 2001, 20, 20-2. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Noguchi, M.; Mine, T.; Komatsu, N.; Yutani, S.; Ueno, T. Characteristics of severe adverse events after peptide vaccination for advanced cancer patients: Analysis of 500 cases. Oncol. Rep. 2011, 25, 57–62. [Google Scholar]
- Cappelletti, M.; Zampaglione, I.; Rizzuto, G.; Ciliberto, G.; La Monica, N.; Fattori, E. Gene electro-transfer improves transduction by modifying the fate of intramuscular plasmid DNA. J. Gene Med. 2003, 5, 324–332. [Google Scholar] [CrossRef]
- Aurisicchio, L.; Fridman, A.; Mauro, D.; Sheloditna, R.; Chiappori, A.; Bagchi, A.; Ciliberto, G. Safety, tolerability and immunogenicity of V934/V935 hTERT vaccination in cancer patients with selected solid tumors: A phase I study. J. Transl. Med. 2020, 18, 39. [Google Scholar] [CrossRef]
- Bajaj, S.; Kumar, M.S.; Peters, G.J.; Mayur, Y.C. Targeting Telomerase for Its Advent in Cancer Therapeutics. Med. Res. Rev. 2020, 40, 1871–1919. [Google Scholar] [CrossRef]
- Trybek, T.; Kowalik, A.; Góźdź, S.; Kowalska, A. Telomeres and Telomerase in Oncogenesis (Review). Oncol. Lett. 2020, 20, 1015–1027. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Judasz, E.; Lisiak, N.; Kopczyński, P.; Taube, M.; Rubiś, B. The Role of Telomerase in Breast Cancer’s Response to Therapy. Int. J. Mol. Sci. 2022, 23, 12844. https://doi.org/10.3390/ijms232112844
Judasz E, Lisiak N, Kopczyński P, Taube M, Rubiś B. The Role of Telomerase in Breast Cancer’s Response to Therapy. International Journal of Molecular Sciences. 2022; 23(21):12844. https://doi.org/10.3390/ijms232112844
Chicago/Turabian StyleJudasz, Eliza, Natalia Lisiak, Przemysław Kopczyński, Magdalena Taube, and Błażej Rubiś. 2022. "The Role of Telomerase in Breast Cancer’s Response to Therapy" International Journal of Molecular Sciences 23, no. 21: 12844. https://doi.org/10.3390/ijms232112844
APA StyleJudasz, E., Lisiak, N., Kopczyński, P., Taube, M., & Rubiś, B. (2022). The Role of Telomerase in Breast Cancer’s Response to Therapy. International Journal of Molecular Sciences, 23(21), 12844. https://doi.org/10.3390/ijms232112844