
Comprehensive and Quantitative Analysis of the Changes in Proteomic and Phosphoproteomic Profiles during Stimulation and Repression of Steroidogenesis in MA-10 Leydig Cells †



Abstract
:1. Introduction
2. Results
2.1. Validation of MA-10 Leydig Cell Responsiveness
2.2. Treatment of MA-10 Leydig Cells with Fsk or Fsk+AICAR Significantly Affects the Levels of 20 Proteins
2.3. Identification of Proteins Differentially Phosphorylated in MA-10 Leydig Cells in Response to Fsk and AICAR
2.3.1. Proteins Differentially Phosphorylated in Response to Fsk
2.3.2. Proteins Differentially Phosphorylated in Response to AMPK Activation
2.3.3. Proteins Differentially Phosphorylated in Response to Combined Fsk+AICAR
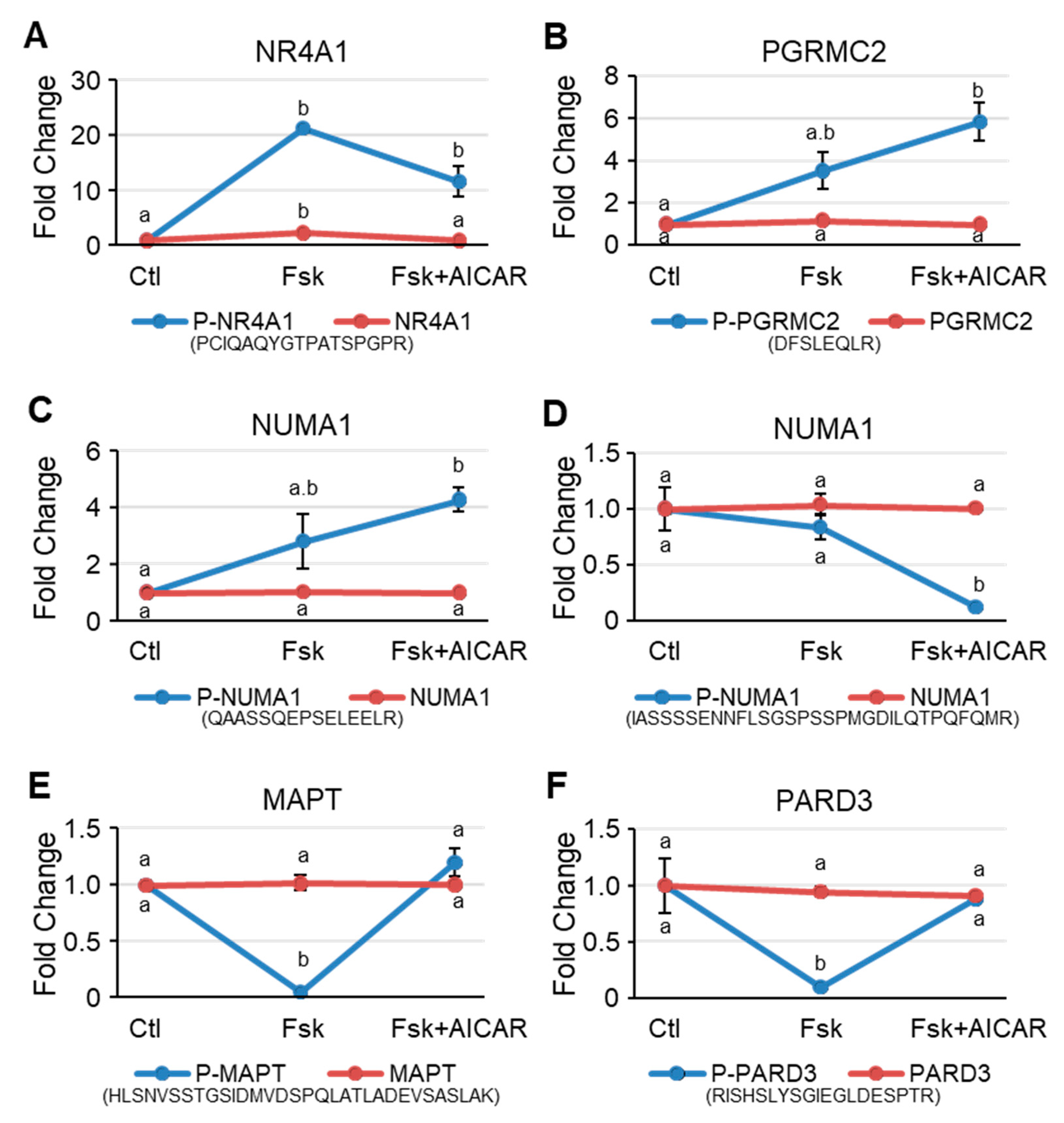
2.4. Phosphorylation Trajectory of Representative Proteins
3. Discussion
3.1. Global Variation in Protein Levels in MA-10 Leydig Cells 1 h after Fsk or Fsk+AICAR Treatment
3.2. Comprehensive Analysis of the Phosphoproteome during Stimulation and Inhibition of Steroidogenesis in MA-10 Leydig Cells
4. Materials and Methods
4.1. Cell Culture
4.2. Chemicals
4.3. RNA Isolation, Reverse Transcription, and Quantitative PCR
4.4. Sample Preparation for LC-MS/MS
4.5. Quantitative Sample Analysis by LC-MS/MS
4.6. Quantitative Data Analyses
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leydig, F. Zur anatomie der mannlichen geschlechtsorgane und analdrusen der saugethiere. Z. Wiss. Zool. 1850, 2, 1–57. [Google Scholar]
- Bouin, P.; Ancel, P. Recherches sur les cellules interstitielles du testicule des mammiferes. Arch. Zool. Exp. Gen. 1903, 1, 437–523. [Google Scholar]
- Hall, P.F.; Irby, D.C.; De Kretser, D.M. Conversion of cholesterol to androgens by rat testes: Comparison of interstitial cells and seminiferous tubules. Endocrinology 1969, 84, 488–496. [Google Scholar] [CrossRef]
- Hughes, I.A. Consensus statement on management of intersex disorders. Arch. Dis. Child. 2005, 91, 554–563. [Google Scholar] [CrossRef]
- Ewing, L.L.; Eik-Nes, K.B. On the formation of testosterone by the perfused rabbit testis. Can. J. Biochem. 1966, 44, 1327–1344. [Google Scholar] [CrossRef] [PubMed]
- Dufau, M.L.; Tsuruhara, T.; Horner, K.A.; Podesta, E.; Catt, K.J. Intermediate role of adenosine 3′:5′-cyclic monophosphate and protein kinase during gonadotropin-induced steroidogenesis in testicular interstitial cells. Proc. Natl. Acad. Sci. USA 1977, 74, 3419–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, J.J. Molecular regulation of steroidogenesis in endocrine Leydig cells. Steroids 2015, 103, 3–10. [Google Scholar] [CrossRef]
- de Mattos, K.; Viger, R.S.; Tremblay, J.J. Transcription factors in the regulation of Leydig cell gene expression and function. Front. Endocrinol. 2022, 13, 881309. [Google Scholar] [CrossRef]
- Martin, L.J.; Tremblay, J.J. The human 3β-hydroxysteroid dehydrogenase/Δ5-Δ4 isomerase type 2 promoter is a novel target for the immediate early orphan nuclear receptor Nur77 in steroidogenic cells. Endocrinology 2005, 146, 861–869. [Google Scholar] [CrossRef]
- Martin, L.J.; Boucher, N.; Brousseau, C.; Tremblay, J.J. The orphan nuclear receptor NUR77 regulates hormone-induced StAR transcription in Leydig cells through cooperation with Ca2+/calmodulin-dependent protein kinase I. Mol. Endocrinol. 2008, 22, 2021–2037. [Google Scholar] [CrossRef] [Green Version]
- Darney, K.J.; Ewing, L. Autoregulation of testosterone secretion in perfused rat testes. Endocrinology 1981, 109, 993–995. [Google Scholar] [CrossRef] [PubMed]
- Shimizu-Albergine, M.; Tsai, L.-C.L.; Patrucco, E.; Beavo, J.A. cAMP-specific phosphodiesterases 8A and 8B, essential regulators of Leydig cell steroidogenesis. Mol. Pharmacol. 2012, 81, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Sanders, M.J.; Underwood, E.; Heath, R.; Mayer, F.V.; Carmena, D.; Jing, C.; Walker, P.A.; Eccleston, J.F.; Haire, L.F.; et al. Structure of mammalian AMPK and its regulation by ADP. Nature 2011, 472, 230–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdou, H.S.; Bergeron, F.; Tremblay, J.J. A cell-autonomous molecular cascade initiated by AMP-activated protein kinase represses steroidogenesis. Mol. Cell. Biol. 2014, 34, 4257–4271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, B.J.; Wells, J.; King, S.R.; Stocco, D.M. The purification, cloning, and expression of a novel luteinizing hormone-induced mitochondrial protein in MA-10 mouse Leydig tumor cells. Characterization of the steroidogenic acute regulatory protein (StAR). J. Biol. Chem. 1994, 269, 28314–28322. [Google Scholar] [CrossRef]
- Abdou, H.S.; Villeneuve, G.; Tremblay, J.J. The calcium signaling pathway regulates Leydig cell steroidogenesis through a transcriptional cascade involving the nuclear receptor NR4A1 and the steroidogenic acute regulatory protein. Endocrinology 2013, 154, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, N.; Fafouri, A.; Bayot, A.; Kumar, M.; Lecharpentier, T.; Ball, G.; Edwards, D.; Bernard, V.; Dournaud, P.; Drunat, S.; et al. Dymeclin deficiency causes postnatal microcephaly, hypomyelination and reticulum-to-Golgi trafficking defects in mice and humans. Hum. Mol. Genet. 2015, 24, 2771–2783. [Google Scholar] [CrossRef]
- Dimitrov, A.; Paupe, V.; Gueudry, C.; Sibarita, J.-B.; Raposo, G.; Vielemeyer, O.; Gilbert, T.; Csaba, Z.; Attie-Bitach, T.; Cormier-Daire, V.; et al. The gene responsible for Dyggve-Melchior-Clausen syndrome encodes a novel peripheral membrane protein dynamically associated with the Golgi apparatus. Hum. Mol. Genet. 2009, 18, 440–453. [Google Scholar] [CrossRef] [Green Version]
- Osipovich, A.B.; Jennings, J.L.; Lin, Q.; Link, A.J.; Ruley, H.E. Dyggve–Melchior–Clausen syndrome: Chondrodysplasia resulting from defects in intracellular vesicle traffic. Proc. Natl. Acad. Sci. USA 2008, 105, 16171–16176. [Google Scholar] [CrossRef] [Green Version]
- Dalla Rosa, I.; Durigon, R.; Pearce, S.F.; Rorbach, J.; Hirst, E.M.A.; Vidoni, S.; Reyes, A.; Brea-Calvo, G.; Minczuk, M.; Woellhaf, M.W.; et al. MPV17L2 is required for ribosome assembly in mitochondria. Nucleic Acids Res. 2014, 42, 8500–8515. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.L.; Bose, H.S. Early steps in steroidogenesis: Intracellular cholesterol trafficking. J. Lipid Res. 2011, 52, 2111–2135. [Google Scholar] [CrossRef] [Green Version]
- Dejgaard, S.; Presley, J. Interactions of lipid droplets with the intracellular transport machinery. Int. J. Mol. Sci. 2021, 22, 2776. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kang, H.; Park, E.; Park, H.-S.; Lee, K. The expression of CKLFSF2B is regulated by GATA1 and CREB in the Leydig cells, which modulates testicular steroidogenesis. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Orlando, U.; Cooke, M.; Maciel, F.C.; Papadopoulos, V.; Podestá, E.J.; Maloberti, P. Characterization of the mouse promoter region of the acyl-CoA synthetase 4 gene: Role of Sp1 and CREB. Mol. Cell. Endocrinol. 2013, 369, 15–26. [Google Scholar] [CrossRef]
- Manna, P.R.; Stocco, D.M. Crosstalk of CREB and Fos/Jun on a single cis-element: Transcriptional repression of the steroidogenic acute regulatory protein gene. J. Mol. Endocrinol. 2007, 39, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Clem, B.F.; Hudson, E.A.; Clark, B.J. Cyclic adenosine 3′,5′-monophosphate (cAMP) enhances cAMP-responsive element binding (CREB) protein phosphorylation and phospho-CREB interaction with the mouse steroidogenic acute regulatory protein gene promoter. Endocrinology 2005, 146, 1348–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, P.; Eubank, D.; Lalli, E.; Sassone-Corsi, P.; Stocco, D. Transcriptional regulation of the mouse steroidogenic acute regulatory protein gene by the cAMP response-element binding protein and steroidogenic factor 1. J. Mol. Endocrinol. 2003, 30, 381–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, P.R.; Dyson, M.T.; Eubank, D.W.; Clark, B.J.; Lalli, E.; Sassone-Corsi, P.; Zeleznik, A.J.; Stocco, D.M. Regulation of Steroidogenesis and the Steroidogenic Acute Regulatory Protein by a Member of the cAMP Response-Element Binding Protein Family. Mol. Endocrinol. 2002, 16, 184–199. [Google Scholar] [CrossRef]
- Ofir, R.; Dwarki, V.J.; Rashid, D.; Verma, I.M. CREB represses transcription of fos promoter: Role of phosphorylation. Gene Expr. 1991, 1, 55–60. [Google Scholar]
- Walker, W.H.; Girardet, C.; Habener, J.F. Alternative exon splicing controls a translational switch from activator to repressor isoforms of transcription factor CREB during spermatogenesis. J. Biol. Chem. 1996, 271, 20145–21050. [Google Scholar] [CrossRef]
- Yi, W.-R.; Tu, M.-J.; Yu, A.-X.; Lin, J.; Yu, A.-M. Bioengineered miR-34a modulates mitochondrial inner membrane protein 17 like 2 (MPV17L2) expression toward the control of cancer cell mitochondrial functions. Bioengineered 2022, 13, 12489–12503. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gao, L.; Chen, J.; Zhang, W.-W.; Zhang, X.-Y.; Wang, B.; Zhang, C.; Wang, Y.; Huang, Y.-C.; Wang, H.; et al. Mitochondrial ROS-mediated ribosome stalling and GCN2 activation are partially involved in 1-nitropyrene-induced steroidogenic inhibition in testes. Environ. Int. 2022, 167, 107393. [Google Scholar] [CrossRef] [PubMed]
- Kone, M.; Pullen, T.J.; Sun, G.; Ibberson, M.; Martinez-Sanchez, A.; Sayers, S.; Nguyen-Tu, M.S.; Kantor, C.; Swisa, A.; Dor, Y.; et al. LKB1 and AMPK differentially regulate pancreatic β-cell identity. FASEB J. 2014, 28, 4972–4985. [Google Scholar] [CrossRef] [PubMed]
- Golkowski, M.; Shimizu-Albergine, M.; Suh, H.W.; Beavo, J.A.; Ong, S.-E. Studying mechanisms of cAMP and cyclic nucleotide phosphodiesterase signaling in Leydig cell function with phosphoproteomics. Cell. Signal. 2016, 28, 764. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Merrill, R.A.; Strack, S. A-kinase anchoring protein 1: Emerging roles in regulating mitochondrial form and function in health and disease. Cells 2020, 9, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyson, M.T.; Jones, J.K.; Kowalewski, M.P.; Manna, P.R.; Alonso, M.; Gottesman, M.E.; Stocco, U.M. Mitochondrial A-kinase anchoring protein 121 binds type II protein kinase A and enhances steroidogenic acute regulatory protein-mediated steroidogenesis in MA-10 mouse Leydig tumor cells. Biol. Reprod. 2008, 78, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakane, F.; King, S.R.; Du, Y.; Kallen, C.B.; Walsh, L.P.; Watari, H.; Stocco, D.M.; Strauss, J.F. Phosphorylation of steroidogenic acute regulatory protein (StAR) modulates its steroidogenic activity. J. Biol. Chem. 1997, 272, 32656–32662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pryde, K.R.; Smith, H.L.; Chau, K.-Y.; Schapira, A.H. PINK1 disables the anti-fission machinery to segregate damaged mitochondria for mitophagy. J. Cell Biol. 2016, 213, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakkaraju, A.K.; Mary, C.; Scherrer, A.; Johnson, A.E.; Strub, K. SRP keeps polypeptides translocation-competent by slowing translation to match limiting ER-targeting sites. Cell 2008, 133, 440–451. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-Y.; Ko, C.-Y.; Kao, T.-J.; Yang, W.-B.; Tsai, Y.-T.; Chuang, J.-Y.; Hu, S.-L.; Yang, P.-Y.; Lo, W.-L.; Hsu, T.-I. CYP17A1 maintains the survival of glioblastomas by regulating SAR1-mediated endoplasmic reticulum health and redox homeostasis. Cancers 2019, 11, 1378. [Google Scholar] [CrossRef] [Green Version]
- Anton, K.; Sinclair, J.; Ohoka, A.; Kajita, M.; Ishikawa, S.; Benz, P.M.; Renne, T.; Balda, M.; Jorgensen, C.; Matter, K.; et al. PKA-regulated VASP phosphorylation promotes extrusion of transformed cells from the epithelium. J. Cell Sci. 2014, 127, 3425–3433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, C.; Bright, N.J.; Sastre, M.; Muckett, P.J.; Carling, D. AMP-activated protein kinase (AMPK) is a tau kinase, activated in response to amyloid β-peptide exposure. Biochem. J. 2011, 434, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Alquezar, C.; Arya, S.; Kao, A.W. Tau Post-translational modifications: Dynamic transformers of Tau function, degradation, and aggregation. Front. Neurol. 2020, 11, 595532. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Macara, I.G. Par-3 controls tight junction assembly through the Rac exchange factor Tiam1. Nat. Cell Biol. 2005, 7, 262–269. [Google Scholar] [CrossRef]
- Zhang, L.; Li, J.; Young, L.H.; Caplan, M.J. AMP-activated protein kinase regulates the assembly of epithelial tight junctions. Proc. Natl. Acad. Sci. USA 2006, 103, 17272–17277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, X.; Liu, C.; Wang, Z.; Sun, Y.; Xiong, Y.; Lei, Q.; Guan, K. PARD3 induces TAZ activation and cell growth by promotingLATS1 andPP1 interaction. EMBO Rep. 2015, 16, 975–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Lee, G.; Jeong, M.G.; Kim, H.K.; Won, H.Y.; Choi, Y.; Lee, J.; Nam, M.; Choi, C.S.; Hwang, G.; et al. Transcriptional coactivator with PDZ-binding motif suppresses the expression of steroidogenic enzymes by nuclear receptor 4 A1 in Leydig cells. FASEB J. 2020, 34, 5332–5347. [Google Scholar] [CrossRef] [Green Version]
- Horsley, V.; Pavlath, G.K. Nfat: Ubiquitous regulator of cell differentiation and adaptation. J. Cell Biol. 2002, 156, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.; Luo, C.; Hogan, P.G. Transcription factors of the NFAT family: Regulation and Function. Annu. Rev. Immunol. 1997, 15, 707–747. [Google Scholar] [CrossRef]
- Okamura, H.; Aramburu, J.; Garcia-Rodriguez, C.; Viola, J.; Raghavan, A.; Tahiliani, M.; Zhang, X.; Qin, J.; Hogan, P.G.; Rao, A. concerted dephosphorylation of the transcription factor NFAT1 induces a conformational switch that regulates transcriptional activity. Mol. Cell 2000, 6, 539–550. [Google Scholar] [CrossRef]
- Okamura, H.; Garcia-Rodriguez, C.; Martinson, H.; Qin, J.; Virshup, D.M.; Rao, A. A conserved docking motif for CK1 binding controls the nuclear localization of NFAT1. Mol. Cell. Biol. 2004, 24, 4184–4195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radulescu, A.E.; Cleveland, D.W. NuMA after 30 years: The matrix revisited. Trends Cell Biol. 2010, 20, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhshi, T.; Trimble, W.S. A non-canonical Hedgehog pathway initiates ciliogenesis and autophagy. J. Cell Biol. 2021, 220, e202004179. [Google Scholar] [CrossRef]
- Schaffer, B.E.; Levin, R.S.; Hertz, N.T.; Maures, T.J.; Schoof, M.L.; Hollstein, P.E.; Benayoun, B.A.; Banko, M.R.; Shaw, R.J.; Shokat, K.M.; et al. Identification of AMPK phosphorylation sites reveals a network of proteins involved in cell invasion and facilitates large-scale substrate prediction. Cell Metab. 2015, 22, 907–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, B.C.; Okuyama, M.W.; Müller, K.; Dehnhard, M.; Jewgenow, K. Steroidogenic enzymes, their products and sex steroid receptors during testis development and spermatogenesis in the domestic cat (Felis catus). J. Steroid Biochem. Mol. Biol. 2018, 178, 135–149. [Google Scholar] [CrossRef]
- Ponikwicka-Tyszko, D.; Chrusciel, M.; Pulawska, K.; Bernaczyk, P.; Sztachelska, M.; Guo, P.; Li, X.; Toppari, J.; Huhtaniemi, I.T.; Wolczynski, S.; et al. Mifepristone treatment promotes testicular Leydig cell tumor progression in transgenic mice. Cancers 2020, 12, E3263. [Google Scholar] [CrossRef]
- Wendler, A.; Wehling, M. PGRMC2, a yet uncharacterized protein with potential as tumor suppressor, migration inhibitor, and regulator of cytochrome P450 enzyme activity. Steroids 2013, 78, 555–558. [Google Scholar] [CrossRef]
- Ascoli, M. Characterization of several clonal lines of cultured Leydig tumor cells: Gonadotropin receptors and steroidogenic responses. Endocrinology 1981, 108, 88–95. [Google Scholar] [CrossRef]
- Rahman, N.A.; Huhtaniemi, I.T. Testicular cell lines. Mol. Cell. Endocrinol. 2004, 228, 53–65. [Google Scholar] [CrossRef]
- Selvaraj, V.; Stocco, D.M.; Clark, B.J. Current knowledge on the acute regulation of steroidogenesis. Biol. Reprod. 2018, 99, 13–26. [Google Scholar] [CrossRef] [Green Version]
- King, S.R.; Ronen-Fuhrmann, T.; Timberg, R.; Clark, B.J.; Orly, J.; Stocco, D.M. Steroid production after in vitro transcription, translation, and mitochondrial processing of protein products of complementary deoxyribonucleic acid for steroidogenic acute regulatory protein. Endocrinology 1995, 136, 5165–5176. [Google Scholar] [CrossRef] [PubMed]
- Engeli, R.T.; Fürstenberger, C.; Kratschmar, D.V.; Odermatt, A. Currently available murine Leydig cell lines can be applied to study early steps of steroidogenesis but not testosterone synthesis. Heliyon 2018, 4, e00527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehanovic, S.; E Mendoza-Villarroel, R.; de Mattos, K.; Talbot, P.; Viger, R.S.; Tremblay, J.J. Identification of novel genes and pathways regulated by the orphan nuclear receptor COUP-TFII in mouse MA-10 Leydig cells. Biol. Reprod. 2021, 105, 1283–1306. [Google Scholar] [CrossRef]
- Hébert-Mercier, P.-O.; Bergeron, F.; Robert, N.M.; Mehanovic, S.; Pierre, K.J.; Mendoza-Villarroel, R.E.; de Mattos, K.; Brousseau, C.; Tremblay, J.J. Growth hormone-induced STAT5B regulates Star gene expression through a cooperation with cJUN in mouse MA-10 Leydig cells. Endocrinology 2022, 163, bqab267. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DMSO vs. Fsk | |||||||
|---|---|---|---|---|---|---|---|
| Upregulated by Forskolin | |||||||
| Protein name | Protein | Gene name | Fsk | DMSO | Fsk/DMSO | q-Value | Fsk/DMSO |
| Dymeclin | Q8CHY3 | Dym | 3.64 × 107 | 1.57 × 106 | 23.2 | 5.08 × 10-6 | Up |
| Homologous-pairing protein 2 homolog | O35047 | Psmc3ip | 7.93 × 106 | 1.57 × 106 | 5.06 | 1.07 × 10-4 | Up |
| Acyl-Coenzyme A dehydrogenase family member 12 | D3Z7X0 | Acad12 | 8.76 × 106 | 2.22 × 106 | 3.94 | 2.56 × 10-2 | Up |
| N-acetyltransferase domain containing 1 | Q9DBW3 | Natd1 | 5.61 × 106 | 1.57 × 106 | 3.58 | 1.58 × 10-4 | Up |
| Nuclear receptor subfamily 4 group A member 1 | P12813 | Nr4a1 | 3.76 × 106 | 1.57 × 106 | 2.4 | 1.41 × 10-3 | Up |
| Downregulated by Forskolin | |||||||
| Protein name | Protein | Gene name | Fsk | DMSO | DMSO/Fsk | q-Value | Fsk/DMSO |
| H/ACA ribonucleoprotein complex subunit 3 | Q9CQS2 | Nop10 | 1.21 × 106 | 7.91 × 106 | 6.55 | 1.44 × 10-2 | Down |
| Mpv17-like protein 2 | Q8VIK2 | Mpv17l2 | 1.21 × 106 | 6.46 × 106 | 5.35 | 8.13 × 10-3 | Down |
| Overexpressed in colon carcinoma 1 protein homolog | P0C913 | Occ1 | 1.21 × 106 | 6.36 × 106 | 5.27 | 8.32 × 10-3 | Down |
| Glutathione peroxidase 7 | Q99LJ6 | Gpx7 | 1.21 × 106 | 4.79 × 106 | 3.97 | 3.60 × 10-2 | Down |
| Fsk vs. Fsk+AICAR | |||||||
| Upregulated by AICAR | |||||||
| Protein name | Protein | Gene name | Fsk+AICAR | Fsk | Fsk+AICAR/Fsk | q-Value | Fsk+AICAR/Fsk |
| Cyclic AMP-responsive element-binding protein 1 | Q01147 | Creb1 | 2.19 × 107 | 1.21 × 106 | 18.15 | 3.83 × 10-4 | Up |
| Mpv17-like protein 2 | Q8VIK2 | Mpv17l2 | 6.32 × 106 | 1.21 × 106 | 5.23 | 5.72 × 10-3 | Up |
| Overexpressed in colon carcinoma 1 protein homolog | P0C913 | Occ1 | 5.95 × 106 | 1.21 × 106 | 4.93 | 7.06 × 10-3 | Up |
| HAUS augmin-like complex subunit 3 | Q8QZX2 | Haus3 | 3.98 × 106 | 1.21 × 106 | 3.3 | 3.10 × 10-2 | Up |
| Downregulated by AICAR | |||||||
| Protein name | Protein | Gene name | Fsk+AICAR | Fsk | Fsk/Fsk+AICAR | q-Value | Fsk+AICAR/Fsk |
| Dymeclin | Q8CHY3 | Dym | 1.61 × 106 | 3.64 × 107 | 22.64 | 8.11x 10-6 | Down |
| Ubinuclein-2 | Q80WC1 | Ubn2 | 1.61 × 106 | 1.56 × 107 | 9.7 | 1.71 × 10-5 | Down |
| Ferrochelatase | P22315 | Fech | 1.61 × 106 | 6.69 × 106 | 4.16 | 4.02 × 10-4 | Down |
| Serine/threonine-protein kinase LMTK2 | Q3TYD6 | Lmtk2 | 1.61 × 106 | 4.56 × 106 | 2.84 | 6.67 × 10-3 | Down |
| Armadillo-like helical domain-containing protein 3 | Q6PD19 | Armh3 | 1.61 × 106 | 4.52 × 106 | 2.81 | 8.76 × 10-4 | Down |
| Ribokinase | Q8R1Q9 | Rbks | 1.61 × 106 | 4.05 × 106 | 2.52 | 1.13 × 10-3 | Down |
| Nuclear receptor subfamily 4 group A member 1 | P12813 | Nr4a1 | 1.61 × 106 | 3.76 × 106 | 2.34 | 1.48 × 10-3 | Down |
| DMSO vs. Fsk+AICAR | |||||||
| Upregulated by Forskolin + AICAR | |||||||
| Protein name | Protein | Gene name | Fsk+AICAR | DMSO | Fsk+AICAR/DMSO | q-Value | Fsk+AICAR/DMSO |
| Glutamyl-tRNA(Gln) amidotransferase subunit B. mitochondrial | Q99JT1 | Gatb | 3.56 × 106 | 1.57 × 106 | 2.27 | 8.86 × 10-4 | Up |
| Decapping and exoribonuclease protein | O70348 | Dxo | 2.89 × 106 | 1.57 × 106 | 1.84 | 3.70 × 10-2 | Up |
| Downregulated by Forskolin + AICAR | |||||||
| Protein name | Protein | Gene name | Fsk+AICAR | DMSO | DMSO/Fsk+AICAR | q-Value | Fsk+AICAR/DMSO |
| SAGA-associated factor 29 | Q9DA08 | Sgf29 | 1.61 × 106 | 1.39 × 109 | 864.22 | 3.00 × 10-5 | Down |
| Coiled-coil domain-containing protein 187 | Q8C5V8 | Ccdc187 | 1.61 × 106 | 1.40 × 107 | 8.71 | 3.87 × 10-4 | Down |
| Ferrochelatase | P22315 | Fech | 1.61 × 106 | 6.73 × 106 | 4.19 | 1.34 × 10-4 | Down |
| Armadillo-like helical domain-containing protein 3 | Q6PD19 | Armh3 | 1.61 × 106 | 5.59 × 106 | 3.48 | 3.67 × 10-2 | Down |
| Fsk vs. DMSO | ||||||||
|---|---|---|---|---|---|---|---|---|
| Phosphorylation decreased by Forskolin | ||||||||
| Phosphopeptide Sequence | Protein | Protein Name | Gene Name | Fsk | DMSO | DMSO/ Fsk | q-Value | Fsk/ DMSO |
| IQTSNVTNKNDPK | Q64012 | RNA-binding protein Raly | Raly | 2.12 × 106 | 3.47 × 108 | 163.6 | 4.54 × 10-4 | Down |
| DLPPFEDESEGLLGTEGPMEEEEDGEELIGDGMER | P97310 | DNA replication licensing factor MCM2 | Mcm2 | 2.12 × 106 | 5.46 × 107 | 25.7 | 1.63 × 10-3 | Down |
| IDEPNTPYHNMIGDDEDAYSDSEGNEVMTPDILAK | Q9DCL8 | Protein phosphatase inhibitor 2 | Ppp1r2 | 2.12 × 106 | 5.14 × 107 | 24.19 | 1.53 × 10-3 | Down |
| HLSNVSSTGSIDMVDSPQLATLADEVSASLAK | P10637 | Microtubule-associated protein tau | Mapt | 2.12 × 106 | 4.06 × 107 | 19.14 | 1.53 × 10-3 | Down |
| SSSPSASLTEHEVSDSPGDEPSESPYESADETQTEASVSSK | Q8BVE8 | Histone-lysine N-methyltransferase NSD2 | Whsc1 | 2.12 × 106 | 3.51 × 107 | 16.54 | 8.63 × 10-3 | Down |
| ETGGTYPPSPPPHSSPTPAATVAATVSTAVPGEPLLPR | Q80U72 | Protein scribble homolog | Scrib | 2.12 × 106 | 2.36 × 107 | 11.13 | 1.55 × 10-2 | Down |
| RISHSLYSGIEGLDESPTR | Q99NH2 | Partitioning defective 3 homolog | Pard3 | 2.12 × 106 | 2.19 × 107 | 10.33 | 2.37 × 10-2 | Down |
| KSMYSRVPECQVTTYYYVGFAYLMMR | Q8QZY1 | Eukaryotic translation initiation factor 3 subunit L | Eif3l | 2.12 × 106 | 2.12 × 107 | 9.99 | 3.82 × 10-3 | Down |
| ALEETPPDSPAAEQENSVNCVDPLR | Q8K1K3 | TERF1-interacting nuclear factor 2 | Tinf2 | 2.12 × 106 | 2.03 × 107 | 9.54 | 3.36 × 10-3 | Down |
| LAAQESSEAEDVTVDR | Q6PGL7 | WASH complex subunit FAM21 | Fam21 | 2.12 × 106 | 1.99 × 107 | 9.36 | 4.73 × 10-2 | Down |
| Phosphorylation increased by Forskolin | ||||||||
| Phosphopeptide Sequence | Protein | Protein Name | Gene Name | Fsk | DMSO | Fsk/ DMSO | q-Value | Fsk/ DMSO |
| KSSVEGLEPAENK | P16254 | Signal recognition particle 14 kDa protein | Srp14 | 2.63 × 108 | 2.90 × 106 | 90.6 | 4.54 × 10-4 | Up |
| STSQGSINSPVYSR | Q8K4G5 | Actin-binding LIM protein 1 | Ablim1 | 7.08 × 107 | 2.12 × 106 | 33.48 | 1.63 × 10-3 | Up |
| QNPEQSADEDAEKNEEDSEGSSDEDEDEDGVGNTTFLK | Q8R1B4 | Eukaryotic translation initiation factor 3 subunit C | Eif3c | 7.06 × 107 | 2.12 × 106 | 33.39 | 1.53 × 10-3 | Up |
| PCIQAQYGTPATSPGPR | P12813 | Nuclear receptor subfamily 4 group A member 1 | Nr4a1 | 4.52 × 107 | 2.12 × 106 | 21.36 | 1.53 × 10-3 | Up |
| NTFTAWSEEDSDYEIDDR | Q6ZQ58 | La-related protein 1 | Larp1 | 1.66 × 108 | 1.02 × 107 | 16.28 | 9.61 × 10-3 | Up |
| EKVESAGPGGDSEPTGSTGALAHTPR | O08550 | Histone-lysine N-methyltransferase 2B | Kmt2b | 2.42 × 107 | 2.12 × 106 | 11.44 | 3.17 × 10-3 | Up |
| SPDLSNQNSDQANEEWETASESSDFASER | Q7TSC1 | Protein PRRC2A | Prrc2a | 1.77 × 107 | 2.12 × 106 | 8.35 | 8.52 × 10-3 | Up |
| KVSVEPQDSHQDAQPR | Q8BXB6 | Solute carrier organic anion transporter family member 2B1 | Slco2b1 | 1.97 × 108 | 2.47 × 107 | 7.98 | 1.30 × 10-2 | Up |
| KSSAAAAAAAAAEGALLPQTPPSPR | Q9ERD6 | Ras-specific guanine nucleotide-releasing factor RalGPS2 | Ralgps2 | 1.64 × 107 | 2.12 × 106 | 7.77 | 9.61 × 10-3 | Up |
| SHSESASPSALSSSPNNLSPTGWSQPK | Q99N57 | RAF proto-oncogene serine/threonine-protein kinase | Raf1 | 1.61 × 107 | 2.12 × 106 | 7.63 | 1.30 × 10-2 | Up |
| Fsk+AICAR vs. Fsk | ||||||||
| Phosphorylation decreased by AICAR | ||||||||
| Phosphopeptide Sequence | Protein | Protein Name | Gene Name | Fsk + AICAR | Fsk | Fsk/ Fsk+AICAR | q-Value | Fsk+AICAR/Fsk |
| YDEKTGLALQTEEFIPNYYCTDERR | K7N6T2 | Vomeronasal 2 receptor 67 | Vmn2r67 | 2.15 × 106 | 1.56 × 109 | 727.23 | 1.81 × 10-4 | Down |
| GASAATGIPLESDEDSNDNDNDLENENCMHTN | Q8R5H1 | Ubiquitin carboxyl-terminal hydrolase 15 | Usp15 | 2.15 × 106 | 6.62 × 107 | 30.83 | 2.50 × 10-3 | Down |
| KGSDDDGGDSPVQDIDTPEVDLYQLQVNTLR | O88574 | Histone deacetylase complex subunit SAP30 | Sap30 | 2.15 × 106 | 3.75 × 107 | 17.43 | 1.12 × 10-2 | Down |
| WGQPPSPTPVPRPPDADPNTPSPK | Q6PDQ2 | Chromodomain-helicase-DNA-binding protein 4 | Chd4 | 2.15 × 106 | 3.33 × 107 | 15.52 | 8.64 × 10-3 | Down |
| TNSMGSATGPLPGTK | Q8BZ47 | Zinc finger protein 609 | Znf609 | 2.15 × 106 | 3.21 × 107 | 14.94 | 4.99 × 10-3 | Down |
| NLATSADTPPSTIPGTGK | Q8BKX6 | Serine/threonine-protein kinase SMG1 | Smg1 | 2.15 × 106 | 1.63 × 107 | 7.57 | 4.56 × 10-3 | Down |
| SPHDSKSPLDHRSPLER | E9PZM4 | Chromodomain-helicase-DNA-binding protein 2 | Chd2 | 2.15 × 106 | 1.58 × 107 | 7.35 | 4.63 × 10-3 | Down |
| IASSSSENNFLSGSPSSPMGDILQTPQFQMR | E9Q7G0 | Nuclear mitotic apparatus protein 1 | Numa1 | 2.15 × 106 | 1.40 × 107 | 6.52 | 1.14 × 10-2 | Down |
| NQENVSHLSVSSASPTSSVASAAGSVTSSSLQK | O54826 | Protein AF-10 | Mllt10 | 2.15 × 106 | 1.39 × 107 | 6.45 | 2.00 × 10-2 | Down |
| GSSGEGLPFAEEGNLTIK | Q8VHK1 | Caskin-2 | Caskin2 | 2.15 × 106 | 1.23 × 107 | 5.75 | 1.02 × 10-2 | Down |
| Phosphorylation increased by AICAR | ||||||||
| Phosphopeptide Sequence | Protein | Protein Name | Gene Name | Fsk + AICAR | Fsk | Fsk+AICAR/ Fsk | q-Value | Fsk+AICAR/Fsk |
| ARPAQAPVSEELPPSPKPGK | Q6PGL7 | WASH complex subunit FAM21 | Fam21 | 6.22 × 107 | 2.12 × 106 | 29.29 | 1.54 × 10-2 | Up |
| HLSNVSSTGSIDMVDSPQLATLADEVSASLAK | P10637 | Microtubule-associated protein tau | Mapt | 4.88 × 107 | 2.12 × 106 | 22.99 | 2.50 × 10-3 | Up |
| APSPEPPTEEVAAETNSTPDDLEAQDALSPETTEEK | Q9ESJ4 | NCK-interacting protein with SH3 domain | Nckipsd | 4.73 × 107 | 2.12 × 106 | 22.26 | 2.50 × 10-3 | Up |
| SSSPSASLTEHEVSDSPGDEPSESPYESADETQTEASVSSK | Q8BVE8 | Histone-lysine N-methyltransferase NSD2 | Whsc1 | 3.40 × 107 | 2.12 × 106 | 16.00 | 4.63 × 10-3 | Up |
| VLLTHEVMCSR | O08792 | Transcription factor COE2 | Ebf2 | 3.04 × 107 | 2.12 × 106 | 14.33 | 2.50 × 10-3 | Up |
| VTETEDDSDSDSDDDEDDVHVTIGDIK | Q9D824 | Pre-mRNA 3’-end-processing factor FIP1 | Fip1l1 | 2.40 × 107 | 2.12 × 106 | 11.31 | 2.50 × 10-3 | Up |
| SRSVEDDEEGHLICQSGDVLSAR | P22518 | Dual specificity protein kinase CLK1 | Clk1 | 2.04 × 107 | 2.12 × 106 | 9.63 | 8.25 × 10-3 | Up |
| RISHSLYSGIEGLDESPTR | Q99NH2 | Partitioning defective 3 homolog | Pard3 | 1.94 × 107 | 2.12 × 106 | 9.16 | 3.87 × 10-3 | Up |
| DAEDLSPCLPSSSQEDTAVPSSPGPSDEVSNTEAEAR | G5E8P0 | Gamma-tubulin complex component 6 | Tubgcp6 | 1.92 × 107 | 2.12 × 106 | 9.06 | 3.47 × 10-2 | Up |
| SVDLKTASPESGRSGFQDEESFR | E9Q4Y4 | Centrosomal protein 192 | Cep192 | 1.89 × 107 | 2.12 × 106 | 8.91 | 4.99 × 10-3 | Up |
| Fsk+AICAR vs. DMSO | ||||||||
| Phosphorylation decreased by Forskolin + AICAR | ||||||||
| Phosphopeptide Sequence | Protein | Protein Name | Gene Name | Fsk + AICAR | DMSO | DMSO/Fsk+AICAR | q-Value | Fsk+AICAR/DMSO |
| AADPPAENSSAPEAEQGGAE | P62960 | Nuclease-sensitive element-binding protein 1 | Ybx1 | 2.15 × 106 | 1.60 × 108 | 74.33 | 2.97 × 10-3 | Down |
| DLPPFEDESEGLLGTEGPMEEEEDGEELIGDGMER | P97310 | DNA replication licensing factor MCM2 | Mcm2 | 2.15 × 106 | 5.46 × 107 | 25.4 | 2.27 × 10-3 | Down |
| SLTISVDSASTSR | Q99MZ6 | Unconventional myosin-VIIb | Myo7b | 2.15 × 106 | 4.54 × 107 | 21.12 | 3.65 × 10-2 | Down |
| LDGESDKEQFDDDQK | Q8C1B1 | Calmodulin-regulated spectrin-associated protein 2 | Camsap2 | 2.15 × 106 | 3.27 × 107 | 15.23 | 6.41 × 10-3 | Down |
| ALEETPPDSPAAEQENSVNCVDPLR | Q8K1K3 | TERF1-interacting nuclear factor 2 | Tinf2 | 2.15 × 106 | 2.03 × 107 | 9.43 | 4.79 × 10-3 | Down |
| QATESPAYGIPLKDGSEQTDEEAEGPFSDDEMVTHK | Q99KK1 | Receptor expression-enhancing protein 3 | Reep3 | 2.15 × 106 | 1.89 × 107 | 8.8 | 2.18 × 10-2 | Down |
| HSPIKEEPCGSISETVCK | Q6PDM1 | Male-specific lethal 1 homolog | Msl1 | 2.15 × 106 | 1.88 × 107 | 8.75 | 2.63 × 10-2 | Down |
| QDVETCRPSSPFGR | Q6PG16 | Holliday junction recognition protein | Hjurp | 2.15 × 106 | 1.78 × 107 | 8.27 | 1.96 × 10-2 | Down |
| SSPDPAVNPVPK | Q924W7 | Suppression of tumorigenicity 5 protein | St5 | 2.15 × 106 | 1.73 × 107 | 8.03 | 6.41 × 10-3 | Down |
| SEVNSEDSDIQEVLPVPK | Q7TQI8 | Testis-specific Y-encoded-like protein 2 | Tspyl2 | 2.83 × 106 | 2.20 × 107 | 7.8 | 1.83 × 10-2 | Down |
| Phosphorylation increased by Forskolin + AICAR | ||||||||
| Sequence | Protein | Protein Name | Gene Name | Fsk + AICAR | DMSO | Fsk+AICAR/ DMSO | q-Value | Fsk+AICAR/DMSO |
| KSSVEGLEPAENK | P16254 | Signal recognition particle 14 kDa protein | Srp14 | 2.76 × 108 | 2.90 × 106 | 95.06 | 1.47 × 10-3 | Up |
| TPLGASLDEQSSGTPK | Q922B9 | Sperm-specific antigen 2 homolog | Ssfa2 | 1.37 × 108 | 2.35 × 106 | 58.37 | 1.47 × 10-3 | Up |
| TVSTQHSTESQDNDQPDYDSVASDEDTDVETR | Q9JLQ2 | ARF GTPase-activating protein GIT2 | Git2 | 1.84 × 108 | 3.32 × 106 | 55.33 | 6.87 × 10-3 | Up |
| QNPEQSADEDAEKNEEDSEGSSDEDEDEDGVGNTTFLK | Q8R1B4 | Eukaryotic translation initiation factor 3 subunit C | Eif3c | 8.66 × 107 | 2.12 × 106 | 40.93 | 1.74 × 10-3 | Up |
| STSQGSINSPVYSR | Q8K4G5 | Actin-binding LIM protein 1 | Ablim1 | 8.20 × 107 | 2.12 × 106 | 38.77 | 3.41 × 10-3 | Up |
| LKNDSDLFGLGLEEMGPKESSDEDR | Q5U3K5 | Rab-like protein 6 | Rabl6 | 4.16 × 107 | 2.12 × 106 | 19.67 | 1.76 × 10-3 | Up |
| NTFTAWSEEDSDYEIDDR | Q6ZQ58 | La-related protein 1 | Larp1 | 1.88 × 108 | 1.02 × 107 | 18.39 | 6.41 × 10-3 | Up |
| AELEEMEEVHPSDEEEEETKAESFYQK | Q9QYR6 | Microtubule-associated protein 1A | Map1a | 3.86 × 107 | 2.12 × 106 | 18.26 | 1.76 × 10-3 | Up |
| QSNASSDVEVEEKETNVSKEDTDQEEK | Q99JF8 | PC4 and SFRS1-interacting protein | Psip1 | 3.81 × 107 | 2.12 × 106 | 18.01 | 6.41 × 10-3 | Up |
| PCIQAQYGTPATSPGPR | P12813 | Nuclear receptor subfamily 4 group A member 1 | Nr4a1 | 2.46 × 107 | 2.12 × 106 | 11.62 | 2.39 × 10-2 | Up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demmouche, Z.B.; Tremblay, J.J. Comprehensive and Quantitative Analysis of the Changes in Proteomic and Phosphoproteomic Profiles during Stimulation and Repression of Steroidogenesis in MA-10 Leydig Cells. Int. J. Mol. Sci. 2022, 23, 12846. https://doi.org/10.3390/ijms232112846
Demmouche ZB, Tremblay JJ. Comprehensive and Quantitative Analysis of the Changes in Proteomic and Phosphoproteomic Profiles during Stimulation and Repression of Steroidogenesis in MA-10 Leydig Cells. International Journal of Molecular Sciences. 2022; 23(21):12846. https://doi.org/10.3390/ijms232112846
Chicago/Turabian StyleDemmouche, Zoheir B., and Jacques J. Tremblay. 2022. "Comprehensive and Quantitative Analysis of the Changes in Proteomic and Phosphoproteomic Profiles during Stimulation and Repression of Steroidogenesis in MA-10 Leydig Cells" International Journal of Molecular Sciences 23, no. 21: 12846. https://doi.org/10.3390/ijms232112846
APA StyleDemmouche, Z. B., & Tremblay, J. J. (2022). Comprehensive and Quantitative Analysis of the Changes in Proteomic and Phosphoproteomic Profiles during Stimulation and Repression of Steroidogenesis in MA-10 Leydig Cells. International Journal of Molecular Sciences, 23(21), 12846. https://doi.org/10.3390/ijms232112846