
Transcriptome Analysis to Identify Responsive Genes under Sublethal Concentration of Bifenazate in the Diamondback Moth, Plutella xylostella (Linnaeus, 1758) (Lepidoptera: Plutellidae)

Abstract
:1. Introduction
2. Results
2.1. Susceptibility of P. xylostella Exposed to Bifenazate
2.2. Overview of the RNA-Seq Data
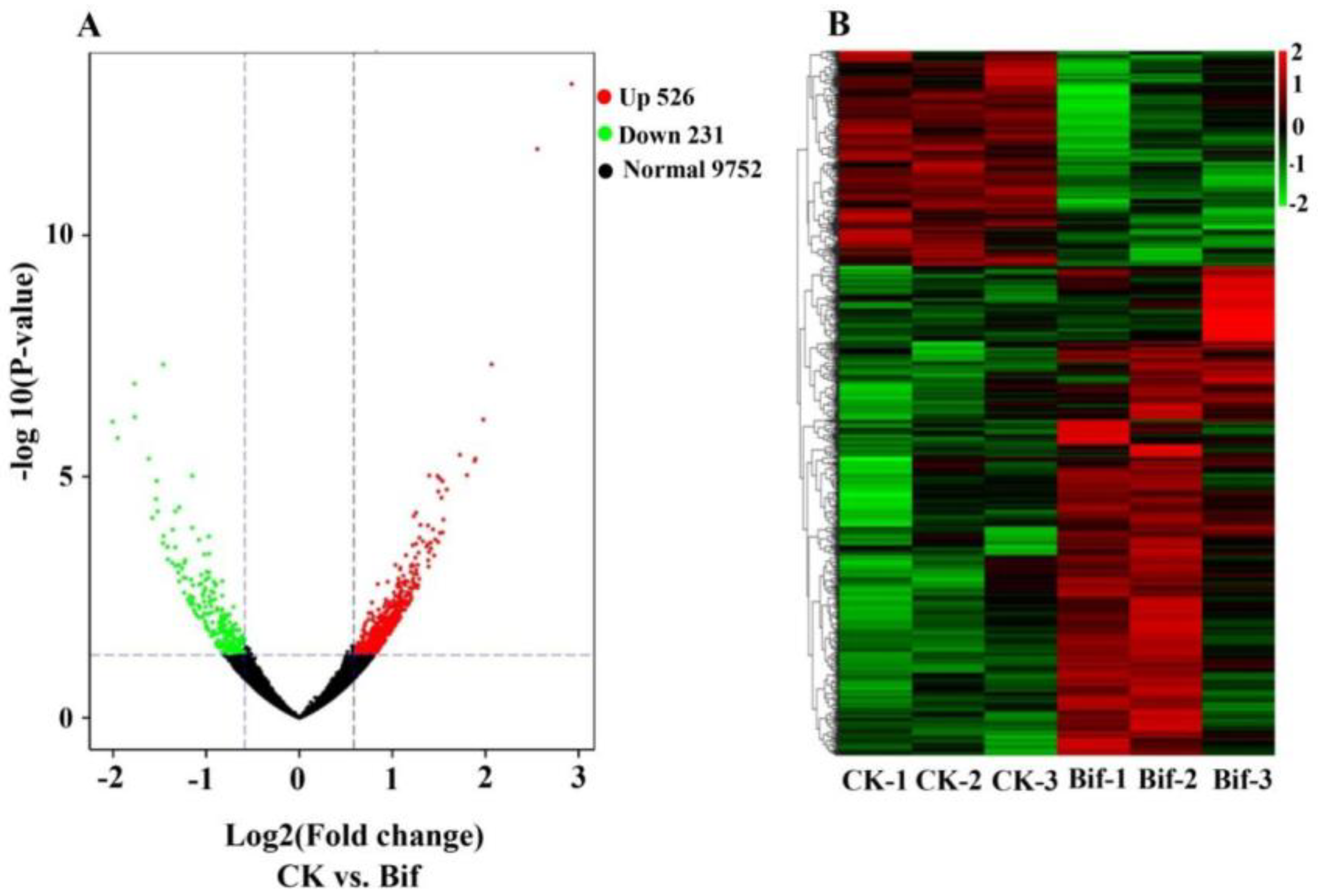
2.3. Analysis of Differentially Expressed Genes (DEGs)
2.4. Gene Functional Annotations of DEGs
2.5. KEGG Enrichment Analysis of DEGs
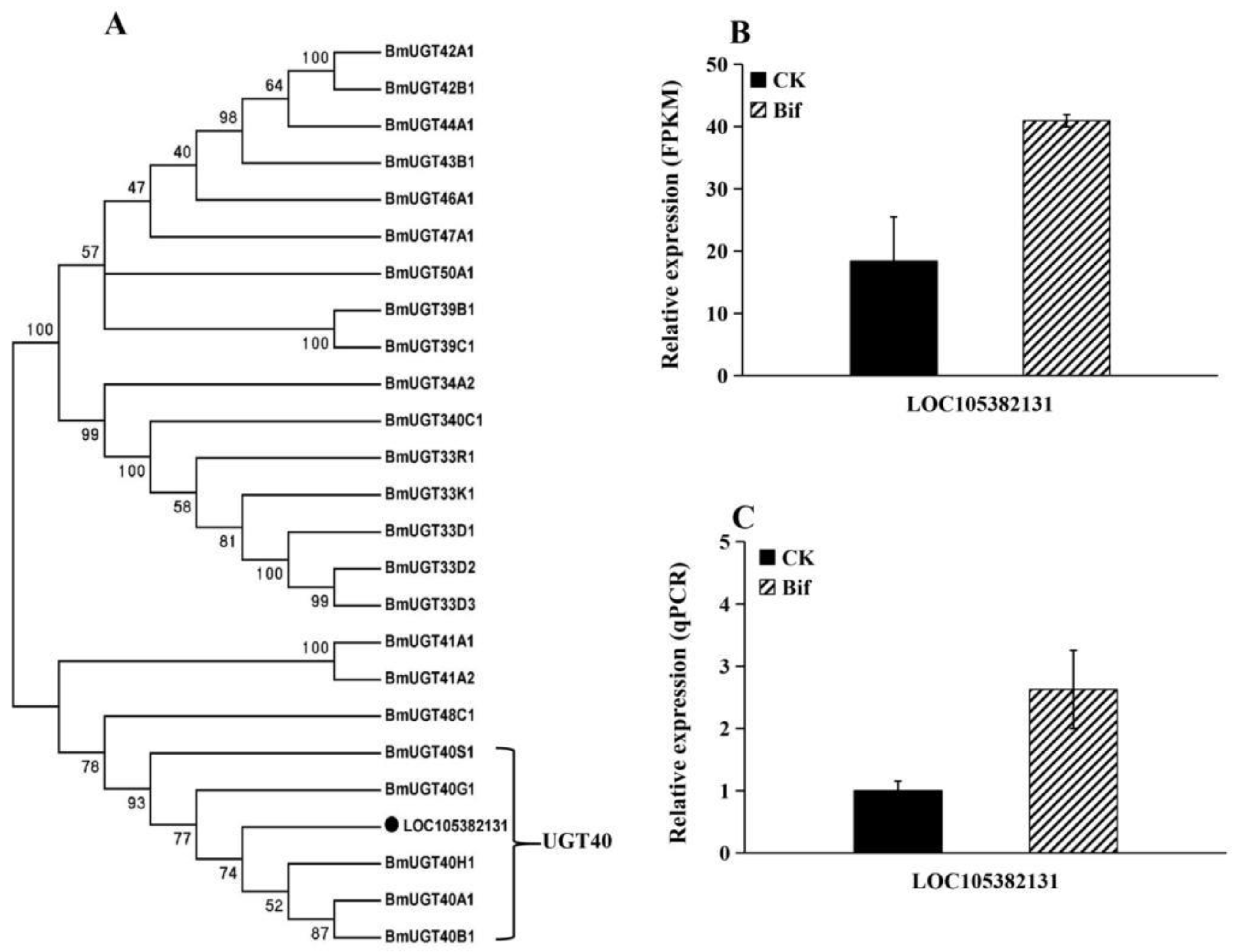
2.6. Changes in Expression Level of Detoxifying Genes
2.7. Changes in Expression Level of Cuticular Protein Genes (CPs)
2.8. Changes in Expression Level of Mitochondrial Genes
3. Discussion
4. Materials and Methods
4.1. Insect Sample and Insecticide
4.2. Insecticide Bioassay
4.3. Insecticide Treatment and Total RNA Extraction
4.4. Library Construction and Sequencing
4.5. Analysis of RNA-Seq Data
4.6. Gene Functional Annotations of DEGs
4.7. KEGG Pathway Enrichment Analysis of DEGs
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandi, A.K.; Kaur, A. Influence of temperature on spinosad toxicity in different populations of Plutella xylostella (Linnaeus). J. Agrometeorol. 2019, 21, 499–503. [Google Scholar] [CrossRef]
- Wang, M.M.; He, M.; Wang, H.; Ma, Y.F.; Dewer, Y.; Zhang, F.; He, P. A candidate aldehyde oxidase in the antennae of the diamondback moth, Plutella xylostella (L.), is potentially involved in the degradation of pheromones, plant-derived volatiles and the detoxification of xenobiotics. Pestic. Biochem. Physiol. 2021, 171, 104726. [Google Scholar] [CrossRef]
- He, W.; You, M.; Vasseur, L.; Yang, G.; Xie, M.; Cui, K.; Bai, J.; Liu, C.; Li, X.; Xu, X.; et al. Developmental and insecticide-resistant insights from the de novo assembled transcriptome of the diamondback moth, Plutella xylostella. Genomics 2012, 99, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Kim, K.; Kwon, D.H.; Jeong, I.H.; Clark, J.M.; Lee, S.H. Transcriptome-based identification and characterization of genes commonly responding to five different insecticides in the diamondback moth, Plutella xylostella. Pestic. Biochem. Physiol. 2018, 144, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mallott, M.; Hamm, S.; Troczka, B.J.; Randall, E.; Pym, A.; Grant, C.; Baxter, S.; Vogel, H.; Shelton, A.M.; Field, L.M.; et al. A flavin-dependent monooxgenase confers resistance to chlorantraniliprole in the diamondback moth, Plutella xylostella. Insect Biochem. Mol. 2019, 115, 103247. [Google Scholar] [CrossRef] [PubMed]
- Van Nieuwenhuyse, P.; Demaeght, P.; Dermauw, W.; Khalighi, M.; Stevens, C.V.; Vanholme, B.; Tirry, L.; Luemmen, P.; Van Leeuwen, T. On the mode of action of bifenazate: New evidence for a mitochondrial target site. Pestic. Biochem. Physiol. 2012, 104, 88–95. [Google Scholar] [CrossRef]
- Ochiai, N.; Mizuno, M.; Mimori, N.; Miyake, T.; Dekeyser, M.; Canlas, L.J.; Takeda, M. Toxicity of bifenazate and its principal active metabolite, diazene, to Tetranychus urticae and Panonychus citri and their relative toxicity to the predaceous mites, Phytoseiulus persimilis and Neoseiulus californicus. Exp. Appl. Acarol. 2007, 43, 181–197. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Van Pottelberge, S.; Tirry, L. Comparative acaricide susceptibility and detoxifying enzyme activities in field-collected resistant and susceptible strains of Tetranychus urticae. Pest Manag. Sci. 2005, 61, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Ninsin, K.D. Acetamiprid resistance and cross-resistance in the diamondback moth, Plutella xylostella. Pest Manag. Sci. 2004, 60, 839–841. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Liang, P.; Shi, X.; Gao, X. Effects of insecticides on the fluidity of mitochondrial membranes of the diamondback moth, Plutella xylostella, resistant and susceptible to avermectin. J. Insect Sci. 2008, 8, 3. [Google Scholar] [CrossRef]
- Wang, X.; Khakame, S.K.; Ye, C.; Yang, Y.; Wu, Y. Characterisation of field-evolved resistance to chlorantraniliprole in the diamondback moth, Plutella xylostella, from China. Pest Manag. Sci. 2013, 69, 661–665. [Google Scholar] [CrossRef]
- Chen, H.; Lin, L.; Ali, F.; Xie, M.; Zhang, G.; Su, W. Using next-generation sequencing to detect differential expression genes in Bradysia odoriphaga after exposure to insecticides. Int. J. Mol. Sci. 2017, 18, 2445. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Ma, Z.; Zhang, X.; Wu, H. Transcriptomic alterations in Sitophilus zeamais in response to allyl isothiocyanate fumigation. Pestic. Biochem. Physiol. 2017, 137, 62–70. [Google Scholar] [CrossRef]
- Sheng, S.; Wang, J.; Chu, J.; Ding, J.; Liu, Z.X.; Jiang, D.; Liang, X.; Shao, Z.; Wang, J.; Wu, F.A. Analysis of the Glyphodes pyloalis larvae immune transcriptome in response to parasitization by its endoparasitoid, Aulacococentrum confusum. Comp. Biochem. Physiol. D 2021, 38, 100803. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Lai, D.; Zhou, Y.; Xu, H.; Zhang, Z.; Kuang, S.; Shao, X. Azadirachtin A inhibits the growth and development of Bactrocera dorsalis larvae by releasing cathepsin in the midgut. Ecotoxicol. Environ. Saf. 2019, 183, 109512. [Google Scholar] [CrossRef]
- Si, F.L.; Qiao, L.; He, Q.Y.; Zhou, Y.; Yan, Z.T.; Chen, B. HSP superfamily of genes in the malaria vector Anopheles sinensis: Diversity, phylogenetics and association with pyrethroid resistance. Malar. J. 2019, 18, 132. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Wang, X. Exploring the multilevel hazards of thiamethoxam using Drosophila melanogaster. J. Hazard. Mater. 2020, 384, 121419. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Xin, X.D.; Liu, Z.X.; Wang, J.; Zhang, R.; Gui, Z.Z. Transcriptional response of detoxifying enzyme genes in Bombyx mori under chlorfenapyr exposure. Pestic. Biochem. Physiol. 2021, 177, 104899. [Google Scholar] [CrossRef]
- Shu, B.; Yu, H.; Li, Y.; Zhong, H.; Li, X.; Cao, L.; Lin, J. Identification of azadirachtin responsive genes in Spodoptera frugiperda larvae based on RNA-seq. Pestic. Biochem. Physiol. 2021, 172, 104745. [Google Scholar] [CrossRef]
- Bian, Y.; Feng, Y.; Zhang, A.; Qi, X.; Pan, J.; Han, J.; Ma, X.; Liang, L. Residue distribution and risk assessment of bifenazate and its metabolite in garlic plant. Food Chem. 2022, 379, 132013. [Google Scholar] [CrossRef]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef]
- Liu, T.; Zhan, X.; Yu, Y.; Wang, S.; Lu, C.; Lin, G.; Zhu, X.; He, W.; You, M.; You, S. Molecular and pharmacological characterization of biogenic amine receptors from the diamondback moth, Plutella xylostella. Pest Manag. Sci. 2021, 77, 4462–4475. [Google Scholar] [CrossRef]
- Xu, L.; Zhao, J.; Xu, D.; Xu, G.; Gu, Z.; Xiao, Z.; Dewer, Y.; Zhang, Y. Application of transcriptomic analysis to unveil the toxicity mechanisms of fall armyworm response after exposure to sublethal chlorantraniliprole. Ecotoxicol. Environ. Saf. 2022, 230, 113145. [Google Scholar] [CrossRef]
- Agwunobi, D.O.; Zhang, M.; Zhang, X.; Wang, T.; Yu, Z.; Liu, J. Transcriptome profile of Haemaphysalis longicornis (Acari: Ixodidae) exposed to Cymbopogon citratus essential oil and citronellal suggest a cytotoxic mode of action involving mitochondrial Ca2+ overload and depolarization. Pestic. Biochem. Physiol. 2021, 179, 104971. [Google Scholar] [CrossRef]
- Chen, E.H.; Duan, J.Y.; Song, W.; Wang, D.X.; Tang, P.A. RNA-seq analysis reveals mitochondrial and cuticular protein genes are associated with phosphine resistance in the rusty grain beetle (Coleoptera:Laemophloeidae). J. Econ. Entomol. 2021, 114, 440–453. [Google Scholar] [CrossRef]
- Yoon, K.S.; Strycharz, J.P.; Baek, J.H.; Sun, W.; Kim, J.H.; Kang, J.S.; Pittendrigh, B.R.; Lee, S.H.; Clark, J.M. Brief exposures of human body lice to sublethal amounts of ivermectin over-transcribes detoxification genes involved in tolerance. Insect Mol. Biol. 2011, 20, 687–699. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zhao, J.; Han, Z.; Chen, M. Comparative transcriptome and RNA interference reveal CYP6DC1 and CYP380C47 related to lambda-cyhalothrin resistance in Rhopalosiphum padi. Pestic. Biochem. Physiol. 2022, 183, 105088. [Google Scholar] [CrossRef]
- Cha, W.H.; Jung, C.R.; Hwang, Y.J.; Lee, D.W. Comparative transcriptome analysis of pheromone biosynthesis-related gene expressions in Plutella xylostella (L.). J. Asia-Pac. Entomol. 2017, 20, 1260–1266. [Google Scholar] [CrossRef]
- Hou, Q.L.; Chen, E.H. RNA-seq analysis of gene expression changes in cuticles during the larval-pupal metamorphosis of Plutella xylostella. Comp. Biochem. Physiol. D 2021, 39, 100869. [Google Scholar] [CrossRef]
- Gao, L.; Qiao, H.; Wei, P.; Moussian, B.; Wang, Y. Xenobiotic responses in insects. Arch. Insect Biochem. Physiol. 2022, 109, e21869. [Google Scholar] [CrossRef]
- Jing, T.X.; Wang, D.F.; Ma, Y.P.; Zeng, L.L.; Meng, L.W.; Zhang, Q.; Dou, W.; Wang, J.J. Genome-wide and expression-profiling analyses of the cytochromeP450genes in Bactrocera dorsalis (Hendel) and screening of candidateP450genes associated with malathion resistance. Pest Manag. Sci. 2020, 76, 2932–2943. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect CYP genes and P450 enzymes. Insect Mol. Biol. Biochem. 2012, 1, 236–316. [Google Scholar]
- Gao, Y.; Kim, J.H.; Jeong, I.H.; Clark, J.M.; Lee, S.H. Transcriptomic identification and characterization of genes commonly responding to sublethal concentrations of six different insecticides in the common fruit fly, Drosophila melanogaster. Pestic. Biochem. Physiol. 2021, 175, 104852. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, M.; Hilliou, F.; Fricaux, T.; Audant, P.; Feyereisen, R.; Le Goff, G. Cytochrome P450s from the fall armyworm (Spodoptera frugiperda): Responses to plant allelochemicals and pesticides. Insect Mol. Biol. 2015, 24, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Dong, F.; Qian, K.; Miao, L.; Yang, X.; Ge, H.; Wu, Z.; Wang, J. Transcriptome analysis reveals global gene expression changes of Chilo suppressalis in response to sublethal dose of chlorantraniliprole. Chemosphere 2019, 234, 648–657. [Google Scholar] [CrossRef]
- Yang, M.L.; Zhang, J.Z.; Zhu, K.Y.; Xuan, T.; Liu, X.J.; Guo, Y.P.; Ma, E.B. Mechanisms of organophosphate resistance in a field population of oriental migratory locust, Locusta migratoria manilensis (Meyen). Arch. Insect Biochem. Physiol. 2009, 71, 3–15. [Google Scholar] [CrossRef]
- Ahn, S.J.; Vogel, H.; Heckel, D.G. Comparative analysis of the UDP-glycosyltransferase multigene family in insects. Insect Biochem. Mol. 2012, 42, 133–147. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Ye, K.; Tu, X.F.; Lu, Y.J.; Thakur, K.; Jiang, L.; Wei, Z.J. Identification and expression profiles of twenty-six glutathione S-transferase genes from rice weevil, Sitophilus oryzae (Coleoptera: Curculionidae). Int. J. Biol. Macromol. 2018, 120, 1063–1071. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.L. Identification and characterisation of multiple glutathione S-transferase genes from the diamondback moth, Plutella xylostella. Pest Manag. Sci. 2015, 71, 592–600. [Google Scholar] [CrossRef]
- Li, X.; Zhu, B.; Gao, X.; Liang, P. Over-expression of UDP-glycosyltransferase gene UGT2B17 is involved in chlorantraniliprole resistance in Plutella xylostella (L.). Pest Manag. Sci. 2017, 73, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, N.T.; Tetreau, G.; Cao, X.; Jiang, H.; Wang, P.; Kanost, M.R. Annotation and expression analysis of cuticular proteins from the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. 2015, 62, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef]
- Chen, E.H.; Hou, Q.L. Identification and expression analysis of cuticular protein genes in the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Pestic. Biochem. Physiol. 2021, 178, 104943. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.H.; Hou, Q.L.; Dou, W.; Wei, D.D.; Yue, Y.; Yang, R.L.; Yang, P.J.; Yu, S.F.; De Schutter, K.; Smagghe, G.; et al. Genome-wide annotation of cuticular proteins in the oriental fruit fly (Bactrocera dorsalis), changes during pupariation and expression analysis of CPAP3 protein genes in response to environmental stresses. Insect Biochem. Mol. 2018, 97, 53–70. [Google Scholar] [CrossRef] [PubMed]
- Balabanidou, V.; Kefi, M.; Aivaliotis, M.; Koidou, V.; Girotti, J.R.; Mijailovsky, S.J.; Patricia Juarez, M.; Papadogiorgaki, E.; Chalepakis, G.; Kampouraki, A.; et al. Mosquitoes cloak their legs to resist insecticides. Proc. Biol. Sci. 2019, 286, 20191091. [Google Scholar] [CrossRef] [Green Version]
- Koganemaru, R.; Miller, D.M.; Adelman, Z.N. Robust cuticular penetration resistance in the common bed bug (Cimex lectularius L.) correlates with increased steady-state transcript levels of CPR-type cuticle protein genes. Pestic. Biochem. Physiol. 2013, 106, 190–197. [Google Scholar] [CrossRef]
- Sun, X.; Guo, J.; Ye, W.; Guo, Q.; Huang, Y.; Ma, L.; Zhou, D.; Shen, B.; Sun, Y.; Zhu, C. Cuticle genes CpCPR63 and CpCPR47 may confer resistance to deltamethrin in Culex pipiens pallens. Parasitol. Res. 2017, 116, 2175–2179. [Google Scholar] [CrossRef]
- Yahouedo, G.A.; Chandre, F.; Rossignol, M.; Ginibre, C.; Balabanidou, V.; Mendez, N.G.A.; Pigeon, O.; Vontas, J.; Cornelie, S. Contributions of cuticle permeability and enzyme detoxification to pyrethroid resistance in the major malaria vector Anopheles gambiae. Sci. Rep. 2018, 7, 11091. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liao, M.; Yang, Q.; Xiao, J.; Hu, Z.; Zhou, L.; Cao, H. Transcriptome profiling reveals differential gene expression of detoxification enzymes in Sitophilus zeamais responding to terpinen-4-ol fumigation. Pestic. Biochem. Physiol. 2018, 149, 44–53. [Google Scholar] [CrossRef]
- Wilson, D.F.; Vinogradov, S.A. Mitochondrial cytochrome c oxidase: Mechanism of action and role in regulating oxidative phosphorylation. J. Appl. Physiol. 2015, 119, 158. [Google Scholar] [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef] [PubMed]
- Fromm, S.; Senkler, J.; Eubel, H.; Peterhaensel, C.; Braun, H.P. Life without complex I: Proteome analyses of an Arabidopsis mutant lacking the mitochondrial NADH dehydrogenase complex. J. Exp. Bot. 2016, 67, 3079–3093. [Google Scholar] [CrossRef]
- Njiru, C.; Saalwaechter, C.; Gutbrod, O.; Geibel, S.; Wybouw, N.; Van Leeuwen, T. A H258Y mutation in subunit B of the succinate dehydrogenase complex of the spider mite Tetranychus urticae confers resistance to cyenopyrafen and pyflubumide, but likely reinforces cyflumetofen binding and toxicity. Insect Biochem. Mol. 2022, 144, 103761. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, D.; Ren, J.; Pu, S.; Wu, H.; Ma, Z. Target verification of allyl isothiocyanate on the core subunits of cytochrome c oxidase in Sitophilus zeamais by RNAi. Pest Manag. Sci. 2021, 77, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Liu, X.; Cooper, A.M.; Silver, K.; Merzendorfer, H.; Zhu, K.Y.; Zhang, J. Vacuolar (H+)-ATPase subunit c is essential for the survival and systemic RNA interference response in Locusta migratoria. Pest Manag. Sci. 2022, 78, 1555–1566. [Google Scholar] [CrossRef] [PubMed]
- Conte, F.; van Buuringen, N.; Voermans, N.C.; Lefeber, D.J. Galactose in human metabolism, glycosylation and congenital metabolic diseases: Time for a closer look. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129898. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.H.; Hou, Q.L.; Dou, W.; Yang, P.J.; Wang, J.J. Expression profiles of tyrosine metabolic pathway genes and functional analysis of DOPA decarboxylase in puparium tanning of Bactrocera dorsalis (Hendel). Pest Manag. Sci. 2022, 78, 344–354. [Google Scholar] [CrossRef]
- Arakane, Y.; Lomakin, J.; Beeman, R.W.; Muthukrishnan, S.; Gehrke, S.H.; Kanost, M.R.; Kramer, K.J. Molecular and functional analyses of amino acid decarboxylases involved in cuticle tanning in Tribolium castaneum. J. Biol. Chem. 2009, 284, 16584–16594. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.H.; Hou, Q.L.; Wei, D.D.; Dou, W.; Liu, Z.; Yang, P.J.; Smagghe, G.; Wang, J.J. Tyrosine hydroxylase coordinates larval-pupal tanning and immunity in oriental fruit fly (Bactrocera dorsalis). Pest Manag. Sci. 2018, 74, 569–578. [Google Scholar] [CrossRef]
- Andersen, S.O. Insect cuticular sclerotization: A review. Insect Biochem. Mol. 2010, 40, 166–178. [Google Scholar] [CrossRef]
- Noh, M.Y.; Muthukrishnan, S.; Kramer, K.J.; Arakane, Y. Cuticle formation and pigmentation in beetles. Curr. Opin. Insect Sci. 2016, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.; Noh, M.Y.; Kramer, K.J.; Muthukrishnan, S.; Arakane, Y. Gene functions in adult cuticle pigmentation of the yellow mealworm, Tenebrio molitor. Insect Biochem. Mol. 2020, 117, 103291. [Google Scholar] [CrossRef] [PubMed]
- Pedley, A.M.; Benkovic, S.J. A new view into the regulation of purine metabolism: The purinosome. Trends Biochem. Sci. 2017, 42, 141–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsac, R.; Pinson, B.; Saint-Marc, C.; Olmedo, M.; Artal-Sanz, M.; Daignan-Fornier, B.; Gomes, J.E. Purine homeostasis is necessary for developmental timing, germline maintenance and muscle integrity in Caenorhabditis elegans. Genetics 2019, 211, 1297–1313. [Google Scholar] [CrossRef]
- Tian, R.; Yang, C.; Chai, S.M.; Guo, H.; Seim, I.; Yang, G. Evolutionary impacts of purine metabolism genes on mammalian oxidative stress adaptation. Zool. Res. 2022, 43, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Kakino, K.; Tanaka, M.; Lee, J.M.; Kusakabe, T.; Banno, Y. A defect in purine nucleotide metabolism in the silkworm, Bombyx mori, causes a translucent larval integument and male infertility. Insect Biochem. Mol. 2020, 126, 103458. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Lu, M.; Han, G.; Du, Y.; Wang, J. Sublethal effects of chlorantraniliprole on development, reproduction and vitellogenin gene (CsVg) expression in the rice stem borer, Chilo Suppressalis. Pest Manag. Sci. 2016, 72, 2280–2286. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Qian, K.; Zhang, N.; Miao, L.; Cai, J.; Lu, M.; Du, Y.; Wang, J. Sublethal effects of chlorantraniliprole on juvenile hormone levels and mRNA expression of JHAMT and FPPS genes in the rice stem borer, Chilo suppressalis. Pest Manag. Sci. 2017, 73, 2111–2117. [Google Scholar] [CrossRef]
- Chen, J.; Lu, Z.; Li, M.; Mao, T.; Wang, H.; Li, F.; Sun, H.; Dai, M.; Ye, W.; Li, B. The mechanism of sublethal chlorantraniliprole exposure causing silkworm pupation metamorphosis defects. Pest Manag. Sci. 2020, 76, 2838–2842. [Google Scholar] [CrossRef]
- Pu, X.; Yang, Y.; Wu, S.; Wu, Y. Characterisation of abamectin resistance in a field-evolved multiresistant population of Plutella xylostella. Pest Manag. Sci. 2010, 66, 371–378. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, S.K.; Kanwar, R.; Goelzenleuchter, M.; Therneau, T.M.; Beutler, A.S. SERE: Single-parameter quality control and sample comparison for RNA-Seq. BMC Genom. 2012, 13, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of exon skipping events differentiate among splicing patterns in sixteen human tissues. F1000Research 2013, 2, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.Z.; Cai, T.; Olyarchuk, J.G.; Wei, L.P. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Guo, Z.; Kang, S.; Qin, J.; Gong, L.; Sun, D.; Guo, L.; Zhu, L.; Bai, Y.; Zhang, Z.; et al. Reduced expression of the P-glycoprotein gene PxABCB1 is linked to resistance to Bacillus thuringiensis Cry1Ac toxin in Plutella xylostella (L.). Pest Manag. Sci. 2020, 76, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regression Equation | LC50 (95% CI) (mg/L) | LC30 (95% CI) (mg/L) | χ2 |
|---|---|---|---|
| y = −2.97 + 2.35x | 18.38 (3.55–32.14) | 11.63 (1.97–17.36) | 10.73 |
| Samples | Clean Reads | GC Content | Q20 (%) | Q30 (%) | Genome |
|---|---|---|---|---|---|
| CK-1 | 21,023,434 | 49.79% | 97.52 | 93.22 | 74.03 |
| CK-2 | 20,899,860 | 50.33% | 97.80 | 93.89 | 74.44 |
| CK-3 | 20,764,706 | 51.39% | 97.71 | 93.77 | 73.80 |
| Bif-1 | 19,706,331 | 50.53% | 97.64 | 93.47 | 76.81 |
| Bif-2 | 22,127,357 | 52.05% | 98.06 | 94.53 | 75.65 |
| Bif-3 | 20,023,653 | 50.48% | 97.75 | 93.85 | 73.88 |
| Gene ID | p-Value | Log2FC | KEGG Pathway ID | Nr Annotation |
|---|---|---|---|---|
| LOC105381553 | 4.24 × 10−6 | −1.62 | ko00350 | juvenile hormone-suppressible protein |
| LOC105382593 | 0.023 | −0.80 | ko00350 | aromatic-L-amino-acid decarboxylase |
| LOC105386870 | 0.016 | −0.92 | ko00350 | prophenoloxidase |
| LOC105388111 | 0.0028 | −1.04 | ko00350 | acidic juvenile hormone-suppressible protein |
| LOC105388114 | 0.014 | −0.84 | ko00350 | arylphorin subunit beta-like precursor |
| LOC119690367 | 5.82 × 10−7 | −1.77 | ko00350 | juvenile hormone-suppressible protein |
| LOC105381730 | 0.030 | −0.65 | ko00230 | adenosine deaminase CECR1-A-like |
| LOC105382860 | 0.027 | −0.60 | ko00230 | xanthine dehydrogenase |
| LOC105383429 | 0.025 | −0.64 | ko00230 | apyrase-like |
| LOC105383538 | 0.0017 | −1.075 | ko00230 | indole-3-acetaldehyde oxidase-like |
| LOC105384208 | 0.022 | −0.69 | ko00230 | xanthine dehydrogenase |
| LOC105386323 | 0.024 | −0.86 | ko00230 | uricase |
| LOC105393481 | 0.043 | −0.59 | ko00230 | purine nucleoside phosphorylase-like |
| LOC105393606 | 0.041 | −0.66 | ko00230 | phosphoribosylformylglycinamidine synthase |
| LOC105398645 | 0.0075 | −0.83 | ko00230 | amidophosphoribosyltransferase-like |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Q.; Zhang, H.; Zhu, J.; Liu, F. Transcriptome Analysis to Identify Responsive Genes under Sublethal Concentration of Bifenazate in the Diamondback Moth, Plutella xylostella (Linnaeus, 1758) (Lepidoptera: Plutellidae). Int. J. Mol. Sci. 2022, 23, 13173. https://doi.org/10.3390/ijms232113173
Hou Q, Zhang H, Zhu J, Liu F. Transcriptome Analysis to Identify Responsive Genes under Sublethal Concentration of Bifenazate in the Diamondback Moth, Plutella xylostella (Linnaeus, 1758) (Lepidoptera: Plutellidae). International Journal of Molecular Sciences. 2022; 23(21):13173. https://doi.org/10.3390/ijms232113173
Chicago/Turabian StyleHou, Qiuli, Hanqiao Zhang, Jiani Zhu, and Fang Liu. 2022. "Transcriptome Analysis to Identify Responsive Genes under Sublethal Concentration of Bifenazate in the Diamondback Moth, Plutella xylostella (Linnaeus, 1758) (Lepidoptera: Plutellidae)" International Journal of Molecular Sciences 23, no. 21: 13173. https://doi.org/10.3390/ijms232113173