
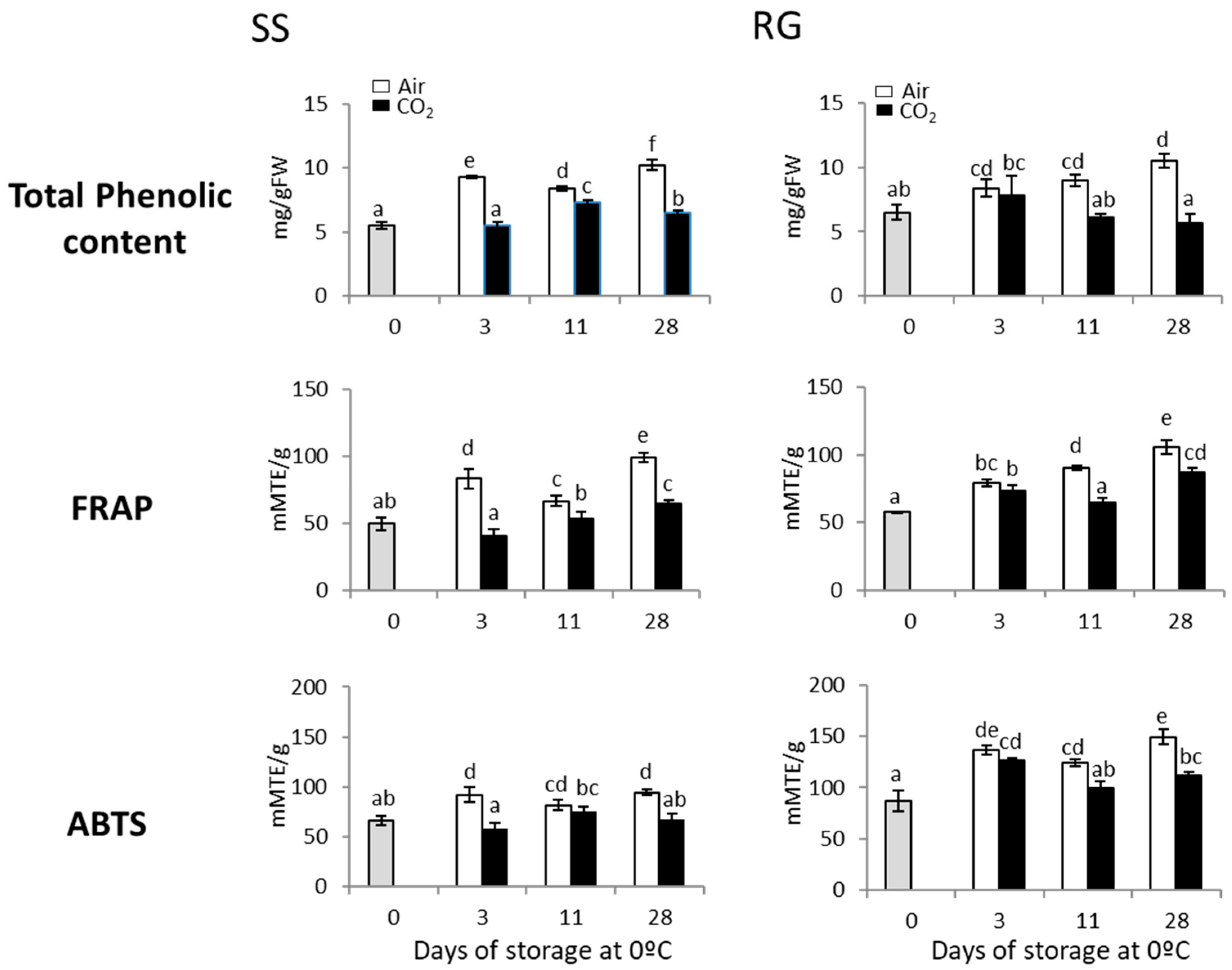
Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Romero, I.; Rosales, R.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect. Int. J. Mol. Sci. 2022, 23, 13304. https://doi.org/10.3390/ijms232113304
Romero I, Rosales R, Escribano MI, Merodio C, Sanchez-Ballesta MT. Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect. International Journal of Molecular Sciences. 2022; 23(21):13304. https://doi.org/10.3390/ijms232113304
Chicago/Turabian StyleRomero, Irene, Raquel Rosales, M. Isabel Escribano, Carmen Merodio, and M. Teresa Sanchez-Ballesta. 2022. "Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect" International Journal of Molecular Sciences 23, no. 21: 13304. https://doi.org/10.3390/ijms232113304
APA StyleRomero, I., Rosales, R., Escribano, M. I., Merodio, C., & Sanchez-Ballesta, M. T. (2022). Short-Term Gaseous Treatments Improve Rachis Browning in Red and White Table Grapes Stored at Low Temperature: Molecular Mechanisms Underlying Its Beneficial Effect. International Journal of Molecular Sciences, 23(21), 13304. https://doi.org/10.3390/ijms232113304