
The Cysteine Protease Giardipain-1 from Giardia duodenalis Contributes to a Disruption of Intestinal Homeostasis
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Giardipain-1 Induces Apoptosis in the Jird Duodenum
2.2. Giardipain-pAC-Transfected Trophozoites Enhance Epithelial Cell Damage in the Duodenum
2.3. Trophozoites Increase Epithelial Cell Proliferation in the Duodenum
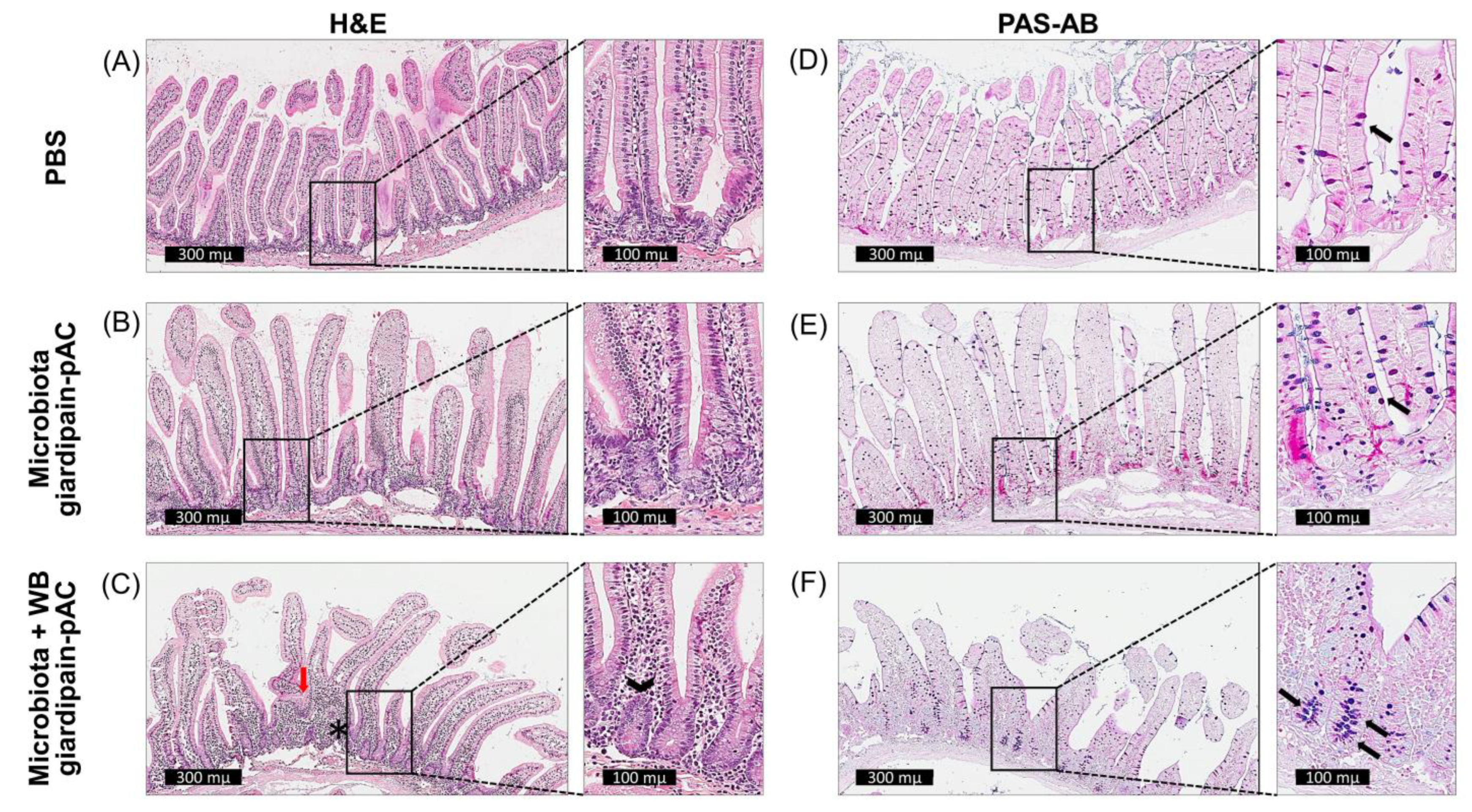
2.4. Giardia Induces Goblet Cell Hyperplasia
2.5. Giardia Triggers a Chronic Inflammatory Response in the Duodenum
2.6. Faecal Microbiota Transplanted to Jirds Infected with Giardia Triggers Epithelial Dysfunction and Changes in the Microbiome Composition
3. Discussion
4. Material and Methods
4.1. Parasite Culture
4.2. Purification, Cloning and Expression of Giardipain-1
4.3. Plasmid Detection in WB Giardipain-pAC Trophozoites by Puromycin-Selection
4.4. Assessing the Effect of Purified Giardipain-1 on Intestinal Loops
4.5. Analysis of Giardipain-1 Expression in WB Giardipain-pAC, WB+ and WB− Trophozoites by Western Blot, and Detection of Giardipain-1 Protease Activity by Zymogram Analysis
4.6. Infection of Jirds and Faecal Microbiota Transplantation (FMT)
4.7. Monitoring the Number of Trophozoites in the Small Intestine of Jirds Infected with WB− or with WB+ or with WB Giardipain-pAC Trophozoites
4.8. Histopathological Examination
4.9. Immunofluorescence
4.10. Immunohistochemistry
4.11. Bacterial DNA Isolation and Illumina-Based Sequencing of 16S Amplicons
4.12. Inferring Microbial Operational Taxonomic Units (OTUs)
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Allaire, J.; Vors, C.; Harris, W.S.; Jackson, K.H.; Tchernof, A.; Couture, P.; Lamarche, B. Comparing the serum TAG response to high-dose supplementation of either DHA or EPA among individuals with increased cardiovascular risk: The ComparED Study. Br. J. Nutr. 2019, 121, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Kapczuk, P.; Kosik-Bogacka, D.; Kupnicka, P.; Metryka, E.; Simińska, D.; Rogulska, K.; Skórka-Majewicz, M.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. The influence of selected gastrointestinal parasites on apoptosis in intestinal epithelial cells. Biomolecules 2020, 10, 674. [Google Scholar] [CrossRef]
- Svärd, S.G.; Hagblom, P.; Palm, J.E.D. Giardia lamblia—A model organism for eukaryotic cell differentiation. FEMS Microbiol. Lett. 2003, 218, 3–7. [Google Scholar] [CrossRef]
- Ringqvist, E.; Palm, J.E.D.; Skarin, H.; Hehl, A.B.; Weiland, M.; Davids, B.J.; Reiner, D.S.; Griffiths, W.J.; Eckmann, L.; Gillin, F.D.; et al. Release of metabolic enzymes by Giardia in response to interaction with intestinal epithelial cells. Mol. Biochem. Parasitol. 2008, 159, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Ma’ayeh, S.Y.; Brook-Carter, P.T. Representational difference analysis identifies specific genes in the interaction of Giardia duodenalis with the murine intestinal epithelial cell line, IEC-6. Int. J. Parasitol. 2012, 42, 501–509. [Google Scholar] [CrossRef]
- Ferella, M.; Davids, B.J.; Cipriano, M.J.; Birkeland, S.R.; Palm, D.; Gillin, F.D.; McArthur, A.G.; Svärd, S. Gene expression changes during Giardia-host cell interactions in serum-free medium. Mol. Biochem. Parasitol. 2014, 197, 21–23. [Google Scholar] [CrossRef]
- Emery, S.J.; Mirzaei, M.; Vuong, D.; Pascovici, D.; Chick, J.M.; Lacey, E.; Haynes, P.A. Induction of virulence factors in Giardia duodenalis independent of host attachment. Sci. Rep. 2016, 6, 20765. [Google Scholar] [CrossRef] [Green Version]
- Dubourg, A.; Xia, D.; Winpenny, J.P.; Al Naimi, S.; Bouzid, M.; Sexton, D.W.; Wastling, J.M.; Hunter, P.R.; Tyler, K.M. Giardia Secretome highlights secreted tenascins as a key component of pathogenesis. Gigascience 2018, 7, giy003. [Google Scholar] [CrossRef]
- Ortega-Pierres, G.; Argüello-García, R.; Laredo-Cisneros, M.S.; Fonseca-Linán, R.; Gómez-Mondragón, M.; Inzunza-Arroyo, R.; Flores-Benítez, D.; Raya-Sandino, A.; Chavez-Munguía, B.; Ventura-Gallegos, J.L.; et al. Giardipain-1, a protease secreted by Giardia duodenalis trophozoites, causes junctional, barrier and apoptotic damage in epithelial cell monolayers. Int. J. Parasitol. 2018, 48, 621–639. [Google Scholar] [CrossRef]
- Argüello-García, R.; Carrero, J.C.; Ortega-Pierres, G. Host immune responses against intestinal unicellular parasites and their role in pathogenesis and protection. In Encyclopedia of Infection and Immunity; Rezaei, N., Ed.; Elsevier: Oxford, UK, 2022; pp. 580–601. ISBN 978-0-323-90303-5. [Google Scholar]
- Cotton, J.A.; Bhargava, A.; Ferraz, J.G.; Yates, R.M.; Beck, P.L.; Buret, A.G. Giardia duodenalis cathepsin b proteases degrade intestinal epithelial interleukin-8 and attenuate interleukin-8-induced neutrophil chemotaxis. Infect. Immun. 2014, 82, 2772–2787. [Google Scholar] [CrossRef] [Green Version]
- Bhargava, A.; Cotton, J.A.; Dixon, B.R.; Gedamu, L.; Yates, R.M.; Buret, A.G. Giardia duodenalis surface cysteine proteases induce cleavage of the intestinal epithelial cytoskeletal protein villin via Myosin light chain kinase. PLoS ONE 2015, 10, e0136102. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ma’ayeh, S.; Peirasmaki, D.; Lundström-Stadelmann, B.; Hellman, L.; Svärd, S.G. Secreted Giardia intestinalis cysteine proteases disrupt intestinal epithelial cell junctional complexes and degrade chemokines. Virulence 2018, 9, 879–894. [Google Scholar] [CrossRef] [Green Version]
- Beatty, J.K.; Akierman, S.V.; Motta, J.-P.; Muise, S.; Workentine, M.L.; Harrison, J.J.; Bhargava, A.; Beck, P.L.; Rioux, K.P.; McKnight, G.W.; et al. Giardia duodenalis induces pathogenic dysbiosis of human intestinal microbiota biofilms. Int. J. Parasitol. 2017, 47, 311–326. [Google Scholar] [CrossRef]
- Gerbaba, T.K.; Green-Harrison, L.; Buret, A.G. Modeling host-microbiome interactions in Caenorhabditis elegans. J. Nematol. 2017, 49, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Fekete, E.; Allain, T.; Siddiq, A.; Sosnowski, O.; Buret, A.G. Giardia Spp. and the gut microbiota: Dangerous liaisons. Front. Microbiol. 2020, 11, 618106. [Google Scholar] [CrossRef]
- DuBois, K.N.; Abodeely, M.; Sajid, M.; Engel, J.C.; McKerrow, J.H. Giardia lamblia cysteine proteases. Parasitol. Res. 2006, 99, 313–316. [Google Scholar] [CrossRef]
- Belosevic, M.; Faubert, G.M.; MacLean, J.D.; Law, C.; Croll, N.A. Giardia lamblia infections in Mongolian gerbils: An Animal Model. J. Infect. Dis. 1983, 147, 222–226. [Google Scholar] [CrossRef]
- Astiazarán-García, H.; Espinosa-Cantellano, M.; Castañón, G.; Chávez-Munguía, B.; Martínez-Palomo, A. Giardia lamblia: Effect of infection with symptomatic and asymptomatic isolates on the growth of gerbils (Meriones unguiculatus). Exp. Parasitol. 2000, 95, 128–135. [Google Scholar] [CrossRef]
- Bouza, M.; Maciques, I.; Torres, D.; Núñez, F.A. Giardia lamblia in Mongolian gerbils: Characteristics of infection using different human isolates. Exp. Parasitol. 2000, 96, 43–46. [Google Scholar] [CrossRef]
- Hernández-Sánchez, J.; Liñan, R.F.; Salinas-Tobón, M.d.R.; Ortega-Pierres, G. Giardia duodenalis: Adhesion-deficient clones have reduced ability to establish infection in Mongolian gerbils. Exp. Parasitol. 2008, 119, 364–372. [Google Scholar] [CrossRef]
- Parker, A.; Maclaren, O.J.; Fletcher, A.G.; Muraro, D.; Kreuzaler, P.A.; Byrne, H.M.; Maini, P.K.; Watson, A.J.M.; Pin, C. Cell proliferation within small intestinal crypts is the principal driving force for cell migration on villi. FASEB J. 2017, 31, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Bergstrom, K.S.B.; Guttman, J.A.; Rumi, M.; Ma, C.; Bouzari, S.; Khan, M.A.; Gibson, D.L.; Vogl, A.W.; Vallance, B.A. Modulation of intestinal goblet cell function during infection by an attaching and effacing bacterial pathogen. Infect. Immun. 2008, 76, 796–811. [Google Scholar] [CrossRef] [Green Version]
- Ponce-Macotela, M.; González-Maciel, A.; Reynoso-Robles, R.; Martínez-Gordillo, M.N. Goblet Cells: Are They an unspecific barrier against Giardia intestinalis or a gate? Parasitol. Res. 2008, 102, 509–513. [Google Scholar] [CrossRef]
- Heyworth, M.F.; Carlson, J.R.; Ermak, T.H. Clearance of Giardia muris infection requires helper/inducer T lymphocytes. J. Exp. Med. 1987, 165, 1743–1748. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, K.; Kanai, T. The Gut Microbiota and inflammatory bowel disease. Semin. Immunopathol. 2015, 37, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Slattum, G.M.; Rosenblatt, J. Tumour Cell Invasion: An emerging role for basal epithelial cell extrusion. Nat. Rev. Cancer 2014, 14, 495–501. [Google Scholar] [CrossRef]
- Heller, F.; Florian, P.; Bojarski, C.; Richter, J.; Christ, M.; Hillenbrand, B.; Mankertz, J.; Gitter, A.H.; Bürgel, N.; Fromm, M.; et al. Interleukin-13 is the key effector th2 cytokine in ulcerative colitis that affects epithelial tight junctions, apoptosis, and cell restitution. Gastroenterology 2005, 129, 550–564. [Google Scholar] [CrossRef]
- Schulzke, J.-D.; Bojarski, C.; Zeissig, S.; Heller, F.; Gitter, A.H.; Fromm, M. Disrupted barrier function through epithelial cell apoptosis. Ann. N. Y. Acad. Sci. 2006, 1072, 288–299. [Google Scholar] [CrossRef]
- Delgado, M.E.; Grabinger, T.; Brunner, T. Cell death at the intestinal epithelial front line. FEBS J. 2016, 283, 2701–2719. [Google Scholar] [CrossRef]
- Barash, N.R.; Maloney, J.G.; Singer, S.M.; Dawson, S.C. Giardia alters commensal microbial diversity throughout the murine gut. Infect. Immun. 2017, 85, e00948-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.K.; Green, P.H.R. Endoscopy in celiac disease. Curr. Opin. Gastroenterol. 2005, 21, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.; Balsitis, M.; Gallivan, S.; Dixon, M.F.; Gilmour, H.M.; Shepherd, N.A.; Theodossi, A.; Williams, G.T. Guidelines for the Initial Biopsy Diagnosis of suspected chronic idiopathic inflammatory bowel disease. the british society of gastroenterology initiative. J. Clin. Pathol. 1997, 50, 93–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, C.E.; Brandborg, L.L.; Phelps, P.C.; Taylor, H.C. Studies of celiac disease. I. The apparent identical and specific nature of the duodenal and proximal jejunal lesion in celiac disease and idiopathic sprue. Gastroenterology 1960, 38, 28–49. [Google Scholar] [CrossRef]
- Khan, W.I.; Collins, S.M. Immune-Mediated alteration in gut physiology and its role in host defence in nematode infection. Parasite Immunol. 2004, 26, 319–326. [Google Scholar] [CrossRef]
- Ishikawa, N.; Wakelin, D.; Mahida, Y.R. Role of T helper 2 cells in intestinal goblet cell hyperplasia in mice infected with Trichinella spiralis. Gastroenterology 1997, 113, 542–549. [Google Scholar] [CrossRef]
- Buret, A.; Gall, D.G.; Nation, P.N.; Olson, M.E. Intestinal protozoa and epithelial cell kinetics, structure and function. Parasitol. Today 1990, 6, 375–380. [Google Scholar] [CrossRef]
- Miller, H.R.; Nawa, Y. Nippostrongylus Brasiliensis: Intestinal goblet-cell response in adoptively immunized rats. Exp. Parasitol. 1979, 47, 81–90. [Google Scholar] [CrossRef]
- Khan, W.I.; Abe, T.; Ishikawa, N.; Nawa, Y.; Yoshimura, K. Reduced amount of intestinal mucus by treatment with anti-CD4 antibody interferes with the spontaneous cure of Nippostrongylus Brasiliensis-Infection in mice. Parasite Immunol. 1995, 17, 485–491. [Google Scholar] [CrossRef]
- Hasnain, S.Z.; Wang, H.; Ghia, J.; Haq, N.; Deng, Y.; Velcich, A.; Grencis, R.K.; Thornton, D.J.; Khan, W.I. Mucin gene deficiency in mice impairs host resistance to an enteric parasitic infection. Gastroenterology 2010, 138, 1763–1771.e5. [Google Scholar] [CrossRef]
- Hasnain, S.Z.; Evans, C.M.; Roy, M.; Gallagher, A.L.; Kindrachuk, K.N.; Barron, L.; Dickey, B.F.; Wilson, M.S.; Wynn, T.A.; Grencis, R.K.; et al. Muc5ac: A critical component mediating the rejection of enteric nematodes. J. Exp. Med. 2011, 208, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Takano, J.; Yardley, J.H. Jejunal lesions in patients with Giardiasis and malabsorption. An electron microscopic study. Bull. Johns Hopkins Hosp. 1965, 116, 413–429. [Google Scholar] [PubMed]
- Williamson, A.L.; O’Donoghue, P.J.; Upcroft, J.A.; Upcroft, P. Immune and pathophysiological responses to different strains of Giardia duodenalis in neonatal mice. Int. J. Parasitol. 2000, 30, 129–136. [Google Scholar] [CrossRef]
- Buret, A.; Gall, D.G.; Olson, M.E. Effects of murine Giardiasis on growth, intestinal morphology, and disaccharidase activity. J. Parasitol. 1990, 76, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Amat, C.B.; Motta, J.-P.; Fekete, E.; Moreau, F.; Chadee, K.; Buret, A.G. Cysteine protease-dependent mucous and differential mucin gene expression in Giardia duodenalis infection. Am. J. Pathol. 2017, 187, 2486–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fekete, E.; Allain, T.; Amat, C.B.; Mihara, K.; Saifeddine, M.; Hollenberg, M.D.; Chadee, K.; Buret, A.G. Giardia duodenalis cysteine proteases cleave proteinase-activated receptor-2 to regulate intestinal goblet cell mucin gene expression. Int. J. Parasitol. 2022, 52, 285–292. [Google Scholar] [CrossRef]
- Gerbaba, T.K.; Gupta, P.; Rioux, K.; Hansen, D.; Buret, A.G. Giardia duodenalis-induced alterations of commensal bacteria kill Caenorhabditis elegans: A new model to study microbial-microbial interactions in the gut. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G550–G561. [Google Scholar] [CrossRef] [Green Version]
- Barash, N.R.; Nosala, C.; Pham, J.K.; McInally, S.G.; Gourguechon, S.; McCarthy-Sinclair, B.; Dawson, S.C. Giardia Colonizes and encysts in high-density foci in the murine small intestine. mSphere 2017, 2, e00343-16. [Google Scholar] [CrossRef] [Green Version]
- Bäckhed, F.; Fraser, C.M.; Ringel, Y.; Sanders, M.E.; Sartor, R.B.; Sherman, P.M.; Versalovic, J.; Young, V.; Finlay, B.B. Defining a healthy human gut microbiome: Current concepts, future directions, and clinical applications. Cell Host Microbe 2012, 12, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.I. Honor Thy gut symbionts redux. Science 2012, 336, 1251–1253. [Google Scholar] [CrossRef]
- Buret, A.G.; Motta, J.-P.; Allain, T.; Ferraz, J.; Wallace, J.L. Pathobiont release from dysbiotic gut microbiota biofilms in intestinal inflammatory diseases: A role for iron? J. Biomed. Sci. 2019, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Graziani, F.; Pujol, A.; Nicoletti, C.; Dou, S.; Maresca, M.; Giardina, T.; Fons, M.; Perrier, J. Ruminococcus Gnavus E1 modulates mucin expression and intestinal glycosylation. J. Appl. Microbiol. 2016, 120, 1403–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gophna, U.; Konikoff, T.; Nielsen, H.B. Oscillospira and related bacteria—From metagenomic species to metabolic features. Environ. Microbiol. 2017, 19, 835–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiippala, K.; Kainulainen, V.; Kalliomäki, M.; Arkkila, P.; Satokari, R. Mucosal prevalence and interactions with the epithelium indicate commensalism of Sutterella spp. Front. Microbiol. 2016, 7, 1706. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.C.; Bessman, N.J.; Hepworth, M.R.; Kumar, N.; Shibata, N.; Kobuley, D.; Wang, K.; Ziegler, C.G.K.; Goc, J.; Shima, T.; et al. Lymphoid-tissue-resident commensal bacteria promote members of the IL-10 cytokine family to establish mutualism. Immunity 2016, 44, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.D.; Gillin, F.D.; Spira, W.M.; Nash, T.E. Chronic giardiasis: Studies on drug sensitivity, toxin production, and host immune response. Gastroenterology 1982, 83, 797–803. [Google Scholar] [CrossRef]
- Keister, D.B. Axenic Culture of Giardia lamblia in TYI-S-33 medium supplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 487–488. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ohishi, I.; Sakaguchi, G. Fluid accumulation in mouse ligated intestine inoculated with Clostridium perfringens enterotoxin. Appl. Environ. Microbiol. 1979, 37, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef]
- Nava, P.; Koch, S.; Laukoetter, M.G.; Lee, W.Y.; Kolegraff, K.; Capaldo, C.T.; Beeman, N.; Addis, C.; Gerner-Smidt, K.; Neumaier, I.; et al. Interferon-gamma regulates intestinal epithelial homeostasis through converging beta-catenin signaling pathways. Immunity 2010, 32, 392–402. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012; ISBN 978-1-936113-41-5. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Stevenes, M.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0-2. 2012. Available online: http://cran.r-project.org/package=vegan (accessed on 21 August 2022).









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quezada-Lázaro, R.; Vázquez-Cobix, Y.; Fonseca-Liñán, R.; Nava, P.; Hernández-Cueto, D.D.; Cedillo-Peláez, C.; López-Vidal, Y.; Huerta-Yepez, S.; Ortega-Pierres, M.G. The Cysteine Protease Giardipain-1 from Giardia duodenalis Contributes to a Disruption of Intestinal Homeostasis. Int. J. Mol. Sci. 2022, 23, 13649. https://doi.org/10.3390/ijms232113649
Quezada-Lázaro R, Vázquez-Cobix Y, Fonseca-Liñán R, Nava P, Hernández-Cueto DD, Cedillo-Peláez C, López-Vidal Y, Huerta-Yepez S, Ortega-Pierres MG. The Cysteine Protease Giardipain-1 from Giardia duodenalis Contributes to a Disruption of Intestinal Homeostasis. International Journal of Molecular Sciences. 2022; 23(21):13649. https://doi.org/10.3390/ijms232113649
Chicago/Turabian StyleQuezada-Lázaro, Rodrigo, Yessica Vázquez-Cobix, Rocío Fonseca-Liñán, Porfirio Nava, Daniel Dimitri Hernández-Cueto, Carlos Cedillo-Peláez, Yolanda López-Vidal, Sara Huerta-Yepez, and M. Guadalupe Ortega-Pierres. 2022. "The Cysteine Protease Giardipain-1 from Giardia duodenalis Contributes to a Disruption of Intestinal Homeostasis" International Journal of Molecular Sciences 23, no. 21: 13649. https://doi.org/10.3390/ijms232113649
APA StyleQuezada-Lázaro, R., Vázquez-Cobix, Y., Fonseca-Liñán, R., Nava, P., Hernández-Cueto, D. D., Cedillo-Peláez, C., López-Vidal, Y., Huerta-Yepez, S., & Ortega-Pierres, M. G. (2022). The Cysteine Protease Giardipain-1 from Giardia duodenalis Contributes to a Disruption of Intestinal Homeostasis. International Journal of Molecular Sciences, 23(21), 13649. https://doi.org/10.3390/ijms232113649