
A Vaccine of SARS-CoV-2 S Protein RBD Induces Protective Immunity

Abstract
:1. Introduction
2. Results
2.1. Recombinant Baculovirus Expressing Exogenous Genes Was Rescued in Sf9 Cells Using the Recombinant Bacmid
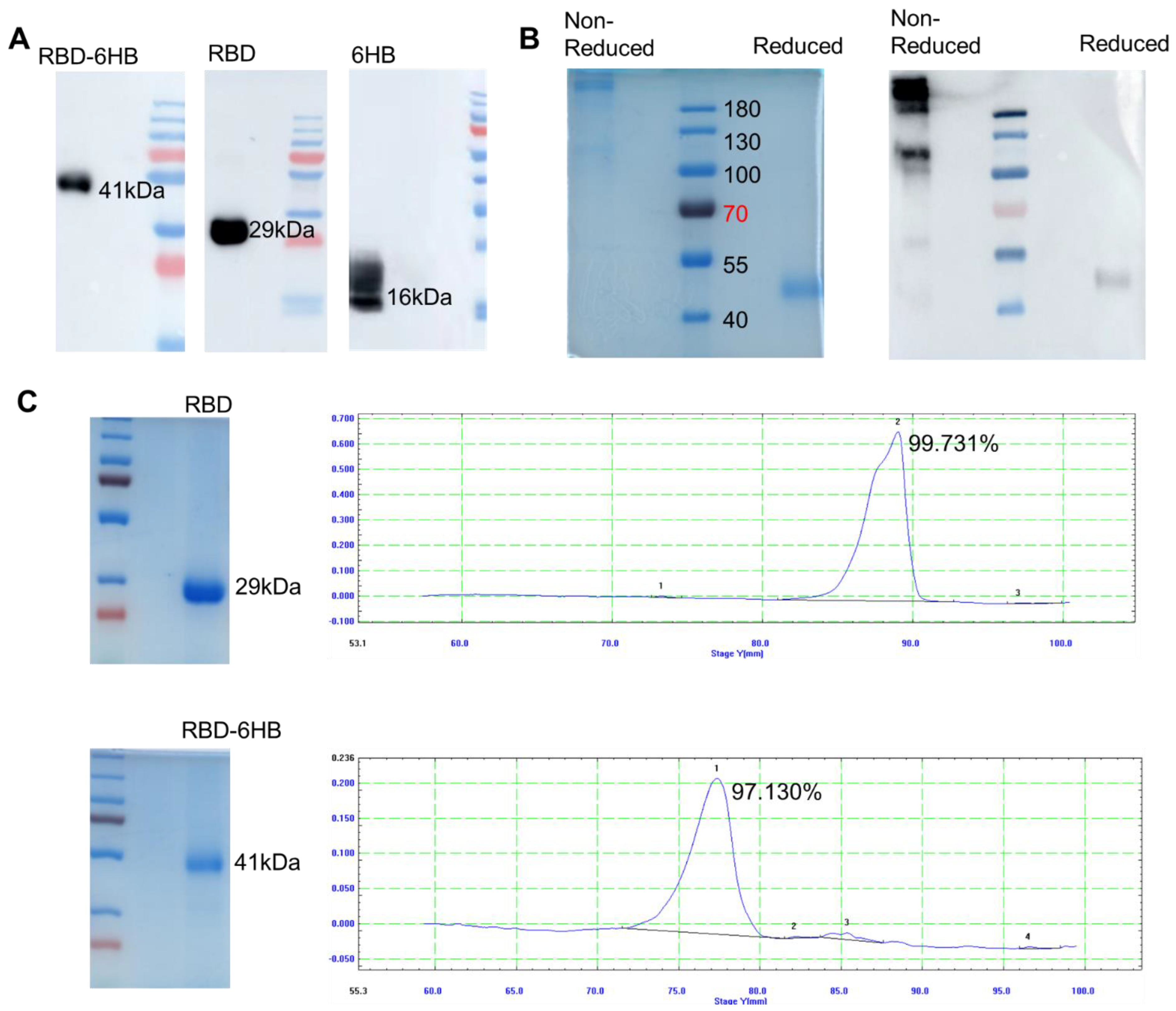
2.2. Protein Expression and Purification
2.3. Specific Antibodies in hACE2 Mice
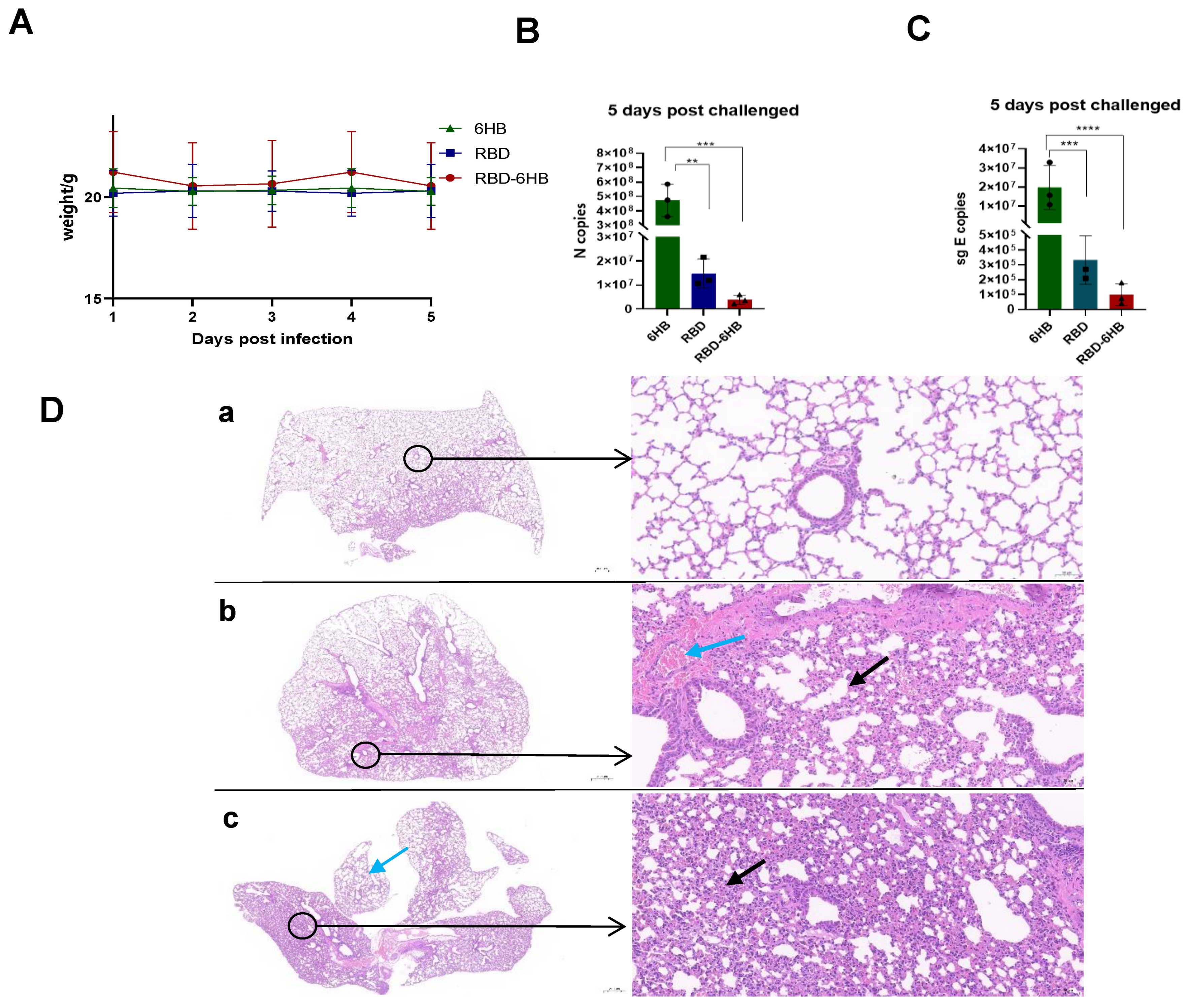
2.4. Complete Protection against Challenge by RBD-6HB Vaccination
3. Discussion
4. Materials and Methods
4.1. Plasmids, Cells, Virus
4.2. Preparation of Recombinant Bacmid
4.3. Preparation of Recombinant Baculovirus
4.4. Western Blot
4.5. Protein Expression and Purification
4.6. Recombinant RBD-6HB Protein Was Identified
4.7. Animal Immunization Experiments
4.8. Immunogenicity Analysis
4.9. Virus Challenge
4.10. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, J.; Xu, W.; Li, L.; Yi, L.; Jiang, Y.; Hao, P.; Xu, Z.; Zou, W.; Li, P.; Gao, Z.; et al. Immunogenicity and protective potential of chimeric virus-like particles containing SARS-CoV-2 spike and H5N1 matrix 1 proteins. Front. Cell. Infect. Microbiol. 2022, 12, 967493. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-Y.; Zhao, R.; Gao, L.-J.; Gao, X.-F.; Wang, D.-P.; Cao, J.-M. SARS-CoV-2: Structure, Biology, and Structure-Based Therapeutics Development. Front. Cell. Infect. Microbiol. 2020, 10, 587269. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Zhao, G.; He, Y.; Guo, Y.; Zheng, B.-J.; Jiang, S.; Zhou, Y. Receptor-binding domain of SARS-CoV spike protein induces long-term protective immunity in an animal model. Vaccine 2007, 25, 2832–2838. [Google Scholar] [CrossRef]
- Li, L.; Wang, M.; Hao, J.; Han, J.; Fu, T.; Bai, J.; Tian, M.; Jin, N.; Zhu, G.; Li, C. Mucosal IgA response elicited by intranasal immunization of Lactobacillus plantarum expressing surface-displayed RBD protein of SARS-CoV-2. Int. J. Biol. Macromol. 2021, 190, 409–416. [Google Scholar] [CrossRef]
- Yang, J.; Wang, W.; Chen, Z.; Lu, S.; Yang, F.; Bi, Z.; Bao, L.; Mo, F.; Li, X.; Huang, Y.; et al. A vaccine targeting the RBD of the S protein of SARS-CoV-2 induces protective immunity. Nature 2020, 586, 572–577. [Google Scholar] [CrossRef]
- Dai, L.; Zheng, T.; Xu, K.; Han, Y.; Xu, L.; Huang, E.; An, Y.; Cheng, Y.; Li, S.; Liu, M.; et al. A Universal Design of Betacoronavirus Vaccines against COVID-19, MERS, and SARS. Cell 2020, 182, 722–733.e11. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef]
- Ye, L.; Sun, Y.; Lin, J.; Bu, Z.; Wu, Q.; Jiang, S.; Steinhauer, D.A.; Compans, R.W.; Yang, C. Antigenic properties of a transport-competent influenza HA/HIV Env chimeric protein. Virology 2006, 352, 74–85. [Google Scholar] [CrossRef]
- Jarząb, A.; Skowicki, M.; Witkowska, D. Subunit vaccines—Antigens, carriers, conjugation methods and the role of adjuvants. Postepy Hig. Med. Dosw. 2013, 67, 1128–1143. [Google Scholar] [CrossRef] [PubMed]
- Kleanthous, H.; Silverman, J.M.; Makar, K.W.; Yoon, I.-K.; Jackson, N.; Vaughn, D.W. Scientific rationale for developing potent RBD-based vaccines targeting COVID-19. npj Vaccines 2021, 6, 128. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.G.; Su, D.; Song, T.-Z.; Zeng, Y.; Huang, W.; Wu, J.; Xu, R.; Luo, P.; Yang, X.; Zhang, X.; et al. S-Trimer, a COVID-19 subunit vaccine candidate, induces protective immunity in nonhuman primates. Nat. Commun. 2021, 12, 1346. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.-Y.; Lin, M.-Y.; Coffman, R.L.; Campbell, J.D.; Traquina, P.; Lin, Y.-J.; Liu, L.T.-C.; Cheng, J.; Wu, Y.-C.; Wu, C.-C.; et al. Development of CpG-adjuvanted stable prefusion SARS-CoV-2 spike antigen as a subunit vaccine against COVID-19. Sci. Rep. 2020, 10, 20085. [Google Scholar] [CrossRef] [PubMed]
- Weisblum, Y.; Schmidt, F.; Zhang, F.; DaSilva, J.; Poston, D.; Lorenzi, J.C.C.; Muecksch, F.; Rutkowska, M.; Hoffmann, H.-H.; Michailidis, E.; et al. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants. eLife 2020, 9, e61312. [Google Scholar] [CrossRef]
- He, X.; Hong, W.; Pan, X.; Lu, G.; Wei, X. SARS-CoV-2 Omicron variant: Characteristics and prevention. MedComm 2021, 2, 838–845. [Google Scholar] [CrossRef]
- Graham, F. Daily briefing: Omicron coronavirus variant puts scientists on alert. Nature Briefing, 26 November 2021. [Google Scholar]
- Leung, K.; Shum, M.H.H.; Leung, G.M.; Lam, T.T.Y.; Wu, J.T. Early transmissibility assessment of the N501Y mutant strains of SARS-CoV-2 in the United Kingdom, October to November 2020. Eurosurveillance 2021, 26, 2002106. [Google Scholar] [CrossRef]
- Tabynov, K.; Turubekov, N.; Babayeva, M.; Fomin, G.; Yerubayev, T.; Yespolov, T.; Li, L.; Renukaradhya, G.K.; Petrovsky, N.; Tabynov, K. An adjuvanted subunit SARS-CoV-2 spike protein vaccine provides protection against COVID-19 infection and transmission. npj Vaccines 2022, 7, 24. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef]
- Martínez-Flores, D.; Zepeda-Cervantes, J.; Cruz-Reséndiz, A.; Aguirre-Sampieri, S.; Sampieri, A.; Vaca, L. SARS-CoV-2 Vaccines Based on the Spike Glycoprotein and Implications of New Viral Variants. Front. Immunol. 2021, 12, 701501. [Google Scholar] [CrossRef]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R.; et al. Community transmission and viral load kinetics of the SARS-CoV-2 delta (B.1.617.2) variant in vaccinated and unvaccinated individuals in the UK: A prospective, longitudinal, cohort study. Lancet Infect. Dis. 2022, 22, 183–195. [Google Scholar] [CrossRef]
- McMillan, C.L.D.; Azuar, A.; Choo, J.J.Y.; Modhiran, N.; Amarilla, A.A.; Isaacs, A.; Honeyman, K.E.; Cheung, S.T.M.; Liang, B.; Wurm, M.J.; et al. Dermal Delivery of a SARS-CoV-2 Subunit Vaccine Induces Immunogenicity against Variants of Concern. Vaccines 2022, 10, 578. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L.; Zheng, M.; Zhou, F. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduct. Target. Ther. 2020, 5, 237. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Krüger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.-S.; Winkler, M.S.; et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2022, 185, 447–456.e11. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Vaccine | Adjuvant | Animals | Immune Times | Interval Times (Weeks) | Number |
|---|---|---|---|---|---|---|
| A | RBD-6HB | CPG10 μL+ aluminum hydroxide 100 μL + 20 μg | hACE2/B | 2 | 6 | 6 |
| B | RBD | CPG10 μL+ aluminum hydroxide 100 μL + 20 μg | hACE2 | 2 | 6 | 6 |
| C | 6HB | CPG10 μL+ aluminum hydroxide 100 μL + 20 μg | hACE2 | 2 | 6 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, Q.; Hao, P.; Xu, W.; Li, L.; Jiang, Y.; Xu, Z.; Chen, J.; Gao, Z.; Pang, Z.; Jin, N.; et al. A Vaccine of SARS-CoV-2 S Protein RBD Induces Protective Immunity. Int. J. Mol. Sci. 2022, 23, 13716. https://doi.org/10.3390/ijms232213716
Qu Q, Hao P, Xu W, Li L, Jiang Y, Xu Z, Chen J, Gao Z, Pang Z, Jin N, et al. A Vaccine of SARS-CoV-2 S Protein RBD Induces Protective Immunity. International Journal of Molecular Sciences. 2022; 23(22):13716. https://doi.org/10.3390/ijms232213716
Chicago/Turabian StyleQu, Qiaoqiao, Pengfei Hao, Wang Xu, Letian Li, Yuhang Jiang, Zhiqiang Xu, Jing Chen, Zihan Gao, Zhaoxia Pang, Ningyi Jin, and et al. 2022. "A Vaccine of SARS-CoV-2 S Protein RBD Induces Protective Immunity" International Journal of Molecular Sciences 23, no. 22: 13716. https://doi.org/10.3390/ijms232213716
APA StyleQu, Q., Hao, P., Xu, W., Li, L., Jiang, Y., Xu, Z., Chen, J., Gao, Z., Pang, Z., Jin, N., & Li, C. (2022). A Vaccine of SARS-CoV-2 S Protein RBD Induces Protective Immunity. International Journal of Molecular Sciences, 23(22), 13716. https://doi.org/10.3390/ijms232213716