

Cathepsin K in Pathological Conditions and New Therapeutic and Diagnostic Perspectives



 and
and 
Abstract
1. Introduction
2. CatK Structure, Mechanism of Action, Genotype, and Polymorphism
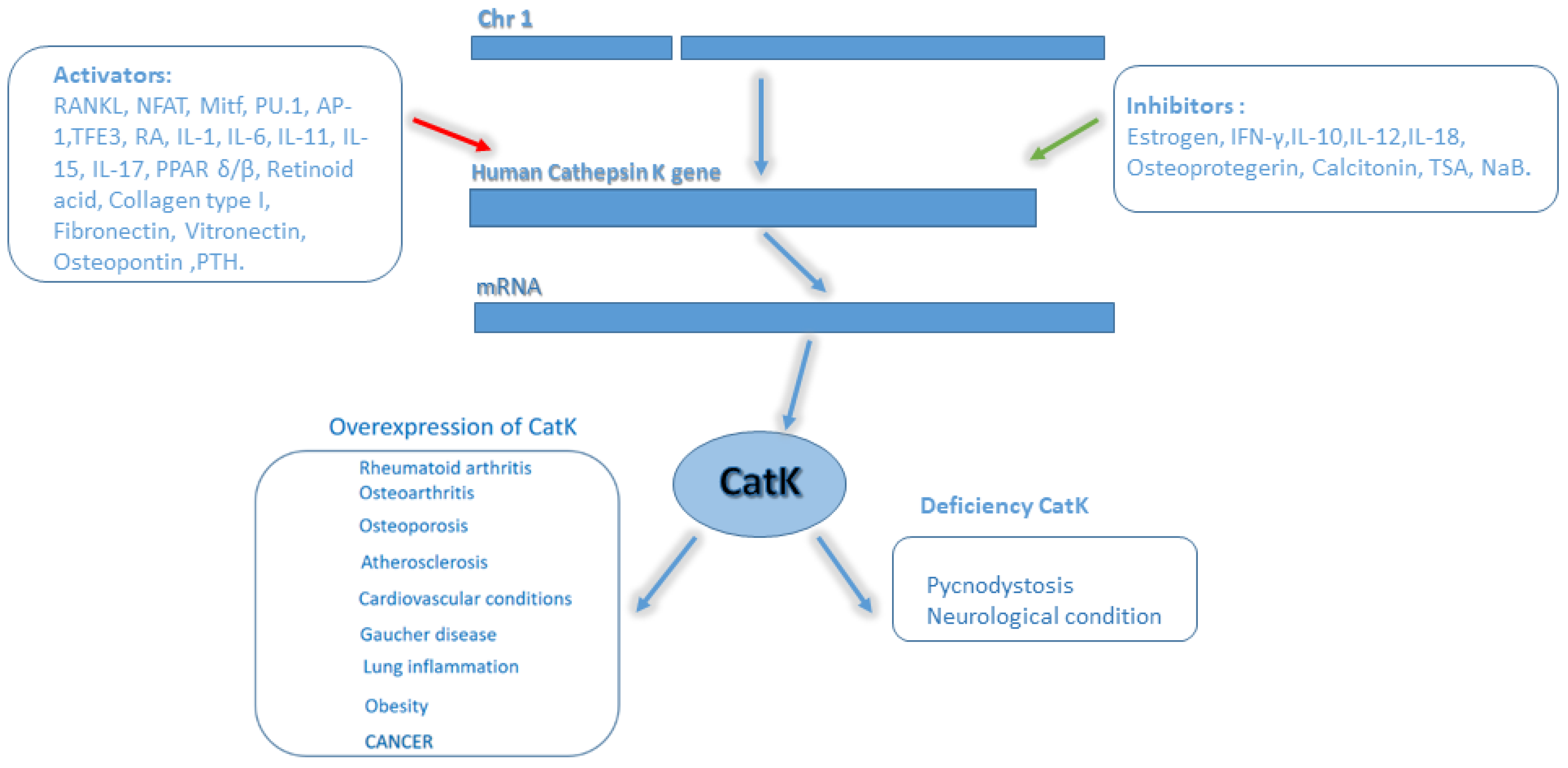
2.1. Regulation of CatK Transcription
3. Role of CatK in Diseases
3.1. Bone
3.2. Heart and Vascular System
3.3. Immune System and Lungs
3.4. Central Nervous System (CNS)
3.5. Skin
3.6. Cancer
4. CatK Inhibitors for Bone and Other Diseases
5. Experimental Perspectives of CatK in Disease Diagnosis and Therapy
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- MEROPS—The Pepotidase Database. EMBL-EBI. 2017. Available online: https://www.ebi.ac.uk/merops/ (accessed on 8 October 2022).
- Ezkurdia, I.; Juan, D.; Rodriguez, J.M.; Frankish, A.; Diekhans, M.; Harrow, J.; Vazquez, J.; Valencia, A.; Tress, M.L. Multiple evidence strands suggest that there may be as few as 19,000 human protein-coding genes. Hum. Mol. Genet. 2014, 23, 5866–5878. [Google Scholar] [CrossRef] [PubMed]
- Trindade, F.; Ferreira, R.; Amado, F.; Vitorino, R. Biofluid proteases profiling in diabetes mellitus. Adv. Clin. Chem. 2015, 69, 161–207. [Google Scholar] [PubMed]
- Troen, B.R. The regulation of cathepsin K gene expression. Ann. N. Y. Acad. Sci. 2006, 1068, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Doucet, A.; Butler, G.S.; Rodriáguez, D.; Prudova, A.; Overall, C.M. Metadegradomics: Toward in vivo quantitative degradomics of proteolytic post-translational modifications of the cancer proteome. Mol. Cell. Proteom. 2008, 7, 1925–1951. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Farquhar, M. Lysosome function in the regulation of the secretory process in cells of the anterior pituitary gland. J. Cell Biol. 1966, 1, 319–347. [Google Scholar] [CrossRef] [PubMed]
- Aits, S.; Jäättelä, M. Lysosomal cell death at a glance. J. Cell Sci. 2013, 126, 1905–1912. [Google Scholar] [CrossRef]
- Mason, S.; Joyce, J. Proteolytic networks in cancer. Trends. Cell Biol. 2011, 21, 228–237. [Google Scholar] [CrossRef]
- Mohamed, M.; Sloane, B. Cysteine cathepsins: Mulfunctional enzymes in cancer. Nat. Rev. Cancer 2006, 6, 764–775. [Google Scholar] [CrossRef]
- Bühling, B.; Röcken, C.; Brasch, F.; Hartig, R.; Yasuda, Y.; Saftig, P.; Brömme, D.; Welte, T. Pivotal role of cathepsin K in lung fibrosis. Am. J. Pathol. 2004, 164, 2203–2216. [Google Scholar] [CrossRef]
- Lecaille, F.; Brömme, D.; Lalmanach, G. Biochemical properties and regulation of cathepsin K activity. Biochimie 2008, 90, 208–226. [Google Scholar] [CrossRef]
- Verbovšek, U.; Van Noorden, C.; Lah, T. Complexity of cancer protease biology: Cathepsin K expression and function in cancer progression. Semin. Cancer Biol. 2015, 35, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, M.; Wang, Z.; Fu, Z.; Lu, A.; Zhang, G. Advances in the discovery of cathepsin K inhibitors on bone resorption. J. Enzyme Inhib. Med. Chem. 2018, 33, 890–904. [Google Scholar] [CrossRef]
- Rocho, F.R.; Bonatto, V.; Lameiro, R.F.; Lameira, J.; Leitão, A.; Montanari, C.A. A patent review on cathepsin K inhibitors to treat osteoporosis (2011–2021). Expert Opin. Ther. Pat. 2022, 32, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yasuda, Y.; Li, W.; Bogyo, M.; Katz, N.; Gordon, R.E.; Fields, G.B.; Brömme, D. Regulation of collagenase activities of human cathepsins by glycosaminoglycans. J. Biol. Chem. 2004, 279, 5470–5479. [Google Scholar] [CrossRef] [PubMed]
- Novinec, M.; Kovacic, L.; Lenarcic, B.; Baici, A. Conformational flexibility and allosteric regulation of cathepsin K. Biochem. J. 2010, 429, 379–389. [Google Scholar] [CrossRef]
- McQueney, M.S.; Amegadzie, B.Y.; D’Alessio, K.; Hanning, C.R.; McLaughlin, M.M.; McNulty, D.; Carr, S.A.; Ijames, C.; Kurdyla, J.; Jones, C.S. Autocatalytic activation of human cathepsin K. J. Biol. Chem. 1997, 272, 13955–13960. [Google Scholar] [CrossRef]
- Yadati, T.; Houben, T.; Bitorina, A.; Shiri-Sverdlov, R. The Ins and Outs of Cathepsins: Physiological Function and Role in Disease Management. Cells 2020, 9, 1679. [Google Scholar] [CrossRef]
- Calio, A.; Brunelli, M.; Gobbo, S.; Argani, P.; Munari, E.; Netto, G.; Martignoni, G. Cathepsin K: A Novel Diagnostic and Predictive Biomarker for Renal Tumors. Cancers 2021, 13, 2441. [Google Scholar] [CrossRef]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef]
- Shalhoub, V.; Faust, J.; Boyle, W.; Dunstan, C.R.; Kelley, M.; Kaufman, S.; Lacey, D.L. Osteoprotegerin and osteoprotegerin ligand effects on osteoclast formation from human peripheral blood mononuclear cell precursors. J. Cell Biochem. 1997, 72, 251–261. [Google Scholar] [CrossRef]
- Hsu, H.; Lacey, D.L.; Dunstan, C.R.; Solovyev, I.; Colombero, A.; Timms, E.; Tan, H.L.; Elliott, G.; Kelley, M.J.; Sarosi, I.; et al. Tumor necrosis factor receptor family member RANK mediates osteoclast differentiation and activation induced by osteoprotegerin ligand. Proc. Natl. Acad. Sci. USA 1999, 96, 3540–3545. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Martinez, A.F.; Fernandez, I.; Balkan, W.; Troen, B.R. AP-1 stimulates the cathepsin K promoter in RAW 264.7 cells. Gene 2007, 403, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Lerner, U.H. Bone remodeling in post-menopausal osteoporosis. J Dent Res. 2006, 85, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Komine, M.; Kukita, A.; Kukita, T.; Ogata, Y.; Hotokebuchi, T.; Kohashi, O. Tumor necrosis factor-alpha cooperates with receptor activator of nuclear factor kappaB ligand in generation of osteoclasts in stromal cell-depleted rat bone marrow cell culture. Bone 2001, 28, 474–483. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kogawa, M.; Wada, S.; Takayanagi, H.; Tsujimoto, M.; Katayama, S.; Nogi, Y. Essential role of p38 mitogen-activated protein kinase n cathepsin K gene expression during osteoclastogenesis through association of NFATc1 and PU.1. J. Biol. Chem. 2004, 279, 45969–45979. [Google Scholar] [CrossRef]
- Mansky, K.; Sankar, U.; Han, J.; Ostrowski, M.C. Microphthalmia transcription factor is a target of the p38 MAPK pathway in response to receptor activator of NF-kappa B ligand signaling. J. Biol. Chem. 2002, 277, 11077–11083. [Google Scholar] [CrossRef]
- George, A.; Zand, D.J.; Hufnagel, R.B.; Sharma, R.; Sergeev, Y.V.; Legare, J.M.; Rice, G.M.; Scott Schwoerer, J.A.; Rius, M.; Tetri, L.; et al. Biallelic Mutations in MITF Cause Coloboma, Osteopetrosis, Microphthalmia, Macrocephaly, Albinism, and Deafness. Am. J. Hum. Genet. 2016, 99, 1388–1394. [Google Scholar] [CrossRef]
- Motyckova, G.; Weilbaecher, K.; Horstmann, M.; Rieman, D.; Fisher, D.Z.; Fisher, F.D. Linking osteopetrosis and pycnodysostosis: Regulation of cathepsin K expression by the microphthalmia transcription factor family. Proc. Natl. Acad. Sci. USA 2001, 98, 5798–5803. [Google Scholar] [CrossRef]
- Saneshige, S.; Mano, H.; Tezuka, K.; Kakudo, S.; Mori, Y.; Honda, Y.; Itabashi, A.; Yamada, T.; Miyata, K.; Hakeda, Y.; et al. Retinoic acid directly stimulates osteoclastic bone resorption and gene expression of cathepsin K/OC-2. Biochem. J. 1995, 309, 721–724. [Google Scholar] [CrossRef]
- Uemura, T.L.; Kuboki, Y. Preliminary communication. MRNA expression of MT1-MMP, MMP-9, cathepsin K, and TRAP in highly enriched osteoclasts cultured on several matrix proteins and ivory surfaces. Biosci. Biotechnol. Biochem. 2000, 64, 1771–1773. [Google Scholar] [CrossRef]
- Wittrant, Y.; Couillaud, S.; Theoleyre, S.; Dunstan, C.; Heymann, D.; Rédini, F. Osteoprotegerin differentially regulates protease expression in osteoclasts cultures. Biochem. Biophys. Res. Commun. 2002, 293, 38–44. [Google Scholar] [CrossRef]
- Mano, H.E. Mammalian mature osteoclasts as estrogen target cells. Biochem. Biophys. Res. Commun. 1996, 223, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Kamolmatyakul, S.W.; Li, J. Interferon-gamma down-regulates gene expression of cathepsin K in osteoclasts and inhibits osteoclast formation. J. Dent. Res. 2001, 80, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Cawley, K.M.; Bustamante-Gomez, N.C.; Guha, A.G.; MacLeod, R.S.; Xiong, J.; Gubrij, I.; Liu, Y.; Mulkey, R.; Palmier, M.; Thostenson, J.D.; et al. Local Production of Osteoprotegerin by Osteoblasts Suppresses Bone Resorption. Cell Rep. 2020, 32, 108052. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Wu, Z.; Chu, H.; Lu, J.; Lyi, A.; Liu, J.; Zhang, G. Cathepsin K: The actionin and beyond bone. Front. Cell Dev. Biol. 2020, 8, 433. [Google Scholar] [CrossRef]
- Bojarski, K.K.; Sage, J.; Lalmanach, G.; Lecaille, F.; Samsonov, S.A. In silico and in vitro mapping of specificity patterns of glycosaminoglycans towards cysteine cathepsins B, L, K, S and V. J. Mol. Graph. Model. 2022, 113, 108153. [Google Scholar] [CrossRef]
- Novinec, M.; Pavšič, M.; Lenarčič, B. A simple and efficient protocol for the production of recombinant cathepsin V and other cysteine cathepsins in soluble form in Escherichia coli. Protein Expr. Purif. 2012, 82, 2012. [Google Scholar] [CrossRef]
- Drake, M.T.; Clarke, B.L.; Oursler, M.J.; Khosla, S. Cathepsin K Inhibitors for Osteoporosis: Biology, Potential Clinical Utility, and Lessons Learned. Endocr. Rev. 2017, 38, 325–350. [Google Scholar] [CrossRef]
- Liang, W.; Wang, F.; Chen, Q.; Dai, J.; Escara-Wilke, J.; Keller, E.T.; Zimmermann, J.; Hong, N.; Lu, Y.; Zhang, J. Targeting cathepsin K diminishes prostate cancer establishment and growth in murine bone. J. Cancer Res. Clin. Oncol. 2019, 145, 1999–2012. [Google Scholar] [CrossRef]
- Hou, W.S.; Li, Z.; Büttner, F.H.; Bartnik, E.; Brömme, D. Cleavage site specificity of cathepsin K toward cartilage proteoglycans and protease complex formation. Biol. Chem. 2003, 384, 891–897. [Google Scholar] [CrossRef]
- Everts, V.; Jansen, I.D.C.; Vries, T.J. Mechanisms of bone resorption. Bone 2022, 163, 116499. [Google Scholar] [CrossRef] [PubMed]
- Aguda, A.H.; Panwar, P.; Du, X.; Nguyen, N.T.; Brayer, G.D.; Brömme, D. Structural basis of collagen fiber degradation by cathepsin K. Proc. Natl. Acad. Sci. USA 2014, 111, 17474–17479. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, J.P.; Suominen, M.I.; Peng, Z.; Halleen, J.M. Secreted tartrate-resistant acid phosphatase 5b is a Marker of osteoclast number in human osteoclast cultures and the rat ovariectomy model. Calcif. Tissue Int. 2008, 82, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Paiva, K.B.; Granjeiro, J.M. Bone tissue remodeling and development: Focus on matrix metalloproteinase functions. Arch. Biochem. Biophys. 2014, 561, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Röcken, C.; Stix, B.; Brömme, D.; Ansorge, S.; Roessner, A.; Bühling, F.A. Putative role for cathepsin K in degradation of AA and AL amyloidosis. Am. J. Pathol. 2001, 158, 1029–1038. [Google Scholar] [CrossRef]
- Motyckova, G.; Fisher, D.E. Pycnodysostosis: Role and regulation of cathepsin K in osteoclast function and human disease. Curr. Mol. Med. 2002, 2, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Panwar, P.; Xue, L.; Søe, K.; Srivastava, K.; Law, S.; Delaisse, J.M.; Brömme, D. An Ectosteric Inhibitor of cathepsin K Inhibits Bone Resorption in Ovariectomized Mice. J. Bone Miner. Res. 2017, 32, 2415–2430. [Google Scholar] [CrossRef] [PubMed]
- Huidekoper, H.H.; Oussoren, E. Mucolipidoses, Multiple Sulfatase Deficiency, and Cathepsin K and C Deficiency. In Physician’s Guide to the Diagnosis, Treatment, and Follow-Up of Inherited Metabolic Diseases; Blau, N., Dionisi Vici, C., Ferreira, C.R., Vianey-Saban, C., van Karnebeek, C.D.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1235–1247. [Google Scholar]
- Zhang, W.; Dong, Z.; Li, D.; Li, B.; Liu, Y.; Zheng, X.; Liu, H.; Zhou, H.; Hu, K.; Xue, Y. Cathepsin K deficiency promotes alveolar bone regeneration by promoting jaw bone marrow mesenchymal stem cells proliferation and differentiation via glycolysis pathway. Cell Prolif. 2021, 54, e13058. [Google Scholar] [CrossRef]
- Migliore, A.; Paoletta, M.; Moretti, A.; Liguori, S.; Iolascon, G. The perspectives of intra-articular therapy in the management of osteoarthritis. Expert Opin. Drug Deliv. 2020, 17, 1213–1226. [Google Scholar] [CrossRef]
- Khoshdel, A.; Forootan, M.; Afsharinasab, M.; Rezaian, M.; Abbasifard, M. Assessment of the circulatory concentrations of cathepsin D, cathepsin K, and alpha-1 antitrypsin in patients with knee osteoarthritis. Ir. J. Med. Sci. 2022. [Google Scholar] [CrossRef]
- Soki, F.N.; Yoshida, R.; Paglia, D.N.; Duong, L.T.; Hansen, M.F.; Drissi, H. Articular cartilage protection in Ctsk−/− mice is associated with cellular and molecular changes in subchondral bone and cartilage matrix. J. Cell. Physiol. 2018, 233, 8666–8676. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Chattopadhyay, N. Pharmacological inhibition of cathepsin K: A promising novel approach for postmenopausal osteoporosis therapy. Biochem. Pharmacol. 2016, 117, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Bolignano, D.; Greco, M.; Arcidiacono, V.; Tripolino, O.; Vita, C.; Provenzano, M.; Donato, C.; Chiarella, S.; Fuiano, G.; De Sarro, G.; et al. Increased circulating Cathepsin-K levels reflect PTH control in chronic hemodialysis patients. J. Nephrol. 2020, 34, 451–458. [Google Scholar] [CrossRef]
- Tantawy, A. Cytokines in Gaucher disease: Role in the pathogenesis of bone and pulmonary disease. Egypt. J. Med. Hum. Genet. 2015, 16, 207–213. [Google Scholar] [CrossRef]
- Quintanilla-Dieck, M.J.; Codriansky, K.; Keady, M.; Bhawan, J.; Rünger, T.M. Cathepsin K in melanoma invasion. J. Investig. Dermatol. 2008, 128, 2281–2288. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Cheng, W.; Shi, G.; Hu, L.; Inoue, A.; Wu, H.; Takeshita, K.; Li, X.; Huang, Z.; Song, H.; et al. Cathepsin K-mediated Notch1 activation contributes to neovascularization in response to hypoxia. Nat. Commun. 2014, 5, 3838. [Google Scholar] [CrossRef]
- Yue, X.; Jiang, H.; Xu, Y.; Xia, M.; Cheng, X.W. Cathepsin K Deficiency Impaired Ischemia-Induced Neovascularization in Aged Mice. Stem Cells Int. 2020, 2020, 6938620. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Jin, J.; Jin, D.; Cui, L.; Li, X.; Rei, Y.; Jiang, H.; Zhao, G.; Yang, G.; et al. Increased serum cathepsin K in patients with coronary artery disease. Yonsei Med. J. 2014, 55, 912–919. [Google Scholar] [CrossRef]
- Guo, R.; Hua, Y.; Rogers, O.; Brown, T.E.; Ren, J.; Nair, S. Cathepsin K knockout protects against cardiac dysfunction in diabetic mice. Sci. Rep. 2017, 7, 8703. [Google Scholar] [CrossRef]
- Lu, P.; Moreland, T.; Christian, C.J.; Lund, T.C.; Steet, R.A.; Steet, H.F. Inappropriate cathepsin K secretion promotes its enzymatic activation driving heart and valve malformation. JCI Insight 2020, 5, e133019. [Google Scholar] [CrossRef]
- Wu, H.; Du, Q.; Dai, Q.; Ge, J.; Cheng, X. Cysteine Protease Cathepsins in Atherosclerotic Cardiovascular Diseases. J. Atheroscler. Thromb. 2018, 25, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Li, C.C.; Qiu, X.T.; Sun, Q.; Zhou, J.P.; Yang, H.J.; Wu, W.Z.; He, L.F.; Tang, K.E.; Zhang, G.G.; Bai, Y.P. Endogenous reduction of miR-185 accelerates cardiac function recovery in mice following myocardial infarction via targeting of cathepsin K. J. Cell. Mol. Med. 2019, 23, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T.; Takashi, K.; Sato, K.; Takaishi, M.; Nakajima, K.; Yamamoto, M.; Kamijima, R.; DiGiovanni, J.; Sano, S. Cathepsin K is involved in development of psoriasis-like skin lesions through TLR-dependent Th17 activation. Immunology 2013, 190, 4805–4811. [Google Scholar]
- Zhao, R.; He, X.W.; Shi, Y.H.; Liu, Y.S.; Liu, F.D.; Hu, Y.; Zhuang, M.T.; Feng, X.Y.; Zhao, L.; Zhao, B.Q.; et al. Cathepsin K knockout exacerbates haemorrhagic transformation induced by recombinant tissue plasminogen activator after focal cerebral ischaemia in mice. Cell. Mol. Neurobiol. 2019, 39, 823–831. [Google Scholar] [CrossRef]
- Aoki, T.; Kataoka, H.; Ishibashi, R.; Nozaki, K.; Hashimoto, N. Cathepsin B, K, and S Are Expressed in Cerebral Aneurysms and Promote the Progression of Cerebral Aneurysms. Stroke 2008, 39, 2603–2610. [Google Scholar] [CrossRef]
- Bernstein, H.; Uwe, L. Cathepsin K and Metabolic Abnormalities in Schizophrenia. Arterioscler. Thromb. Vasc. Biol. 2008, 28, e163. [Google Scholar] [CrossRef][Green Version]
- Bernstein, H.G.; Bukowska, A.; Dobrowolny, H.; Bogerts, B.; Lendeckel, U. Cathepsin K and schizophrenia. Synapse 2007, 61, 252–253. [Google Scholar] [CrossRef]
- Nakajima, K.; Sano, S. Mouse models of psoriasis and their relevance. J. Dermatol. 2018, 45, 252–263. [Google Scholar] [CrossRef]
- Hira, V.; Verbovšek, U.; Breznik, B.; Srdič, M.; Novinec, M.; Kakar, H.; Wormer, J.; Swaan, B.; Lenarčič, B.; Juliano, L.; et al. Cathepsin K cleavage of SDF-1α inhibits its chemotactic activity towards glioblastoma stem-like cells. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 594–603. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Osteoporosis: Current and Emerging Therapies Targeted to Immunological Checkpoints. Curr. Med. Chem. 2020, 27, 6356–6372. [Google Scholar] [CrossRef]
- Qian, D.; He, L.; Zhang, Q.; Li, W.; Tang, D.; Wu, C.; Yang, F.; Li, K.; Zhang, H. Cathepsin K: A Versatile Potential Biomarker and Therapeutic Target for Various Cancers. Curr. Oncol. 2022, 29, 5963–5987. [Google Scholar] [CrossRef] [PubMed]
- Kleer, C.G.; Bloushtain-Qimron, N.; Chen, Y.H.; Carrasco, D.; Hu, M.; Yao, J.; Kraeft, S.K.; Collins, L.C.; Sabel, M.S.; Argani, P.; et al. Epithelial and stromal cathepsin K and CXCL14 expression in breast tumor progression. Clin. Cancer Res. 2008, 17, 5357–5367. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, J.; Hanemaaijer, R.; Mulder, A.; Dijkstra, P.; Szuhai, K.; Brömme, D.; Verheijen, J.H.; Hogendoorn, P.W.C. Cathepsin K is the principal protease in giant cell tumor of bone. Am. J. Pathol. 2004, 165, 593–600. [Google Scholar] [CrossRef]
- Husmann, K.; Muff, R.; Bolander, M.; Sarkar, G.; Born, W.; Fuchs, B. Cathepsins and osteosarcoma: Expression analysis identifies cathepsin K as an indicator of metastasis. Mol. Carcinog. 2008, 73, 66–73. [Google Scholar] [CrossRef]
- Rao, Q.; Cheng, L.; Xia, Q.-Y.; Liu, B.; Li, L.; Shi, Q.-L.; Shi, S.-S.; Yu, B.; Zhang, R.-S.; Ma, H.-H.; et al. Cathepsin K expression in a wide spectrum of perivascular epithelioid cell neoplasms (PEComas): A clinicopathological study emphasizing extrarenal PEComas. Histopatology 2013, 62, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Kojima, F.; Okabe, H. cathepsin K expression in basal cell carcinoma. J. Eur. Acad. Dermatol. Venereol. 2013, 27, e128–e130. [Google Scholar] [CrossRef]
- Bitu, C.C.; Kauppila, J.H.; Bufalino, A.; Nurmenniemi, S.; Teppo, S.; Keinänen, M.; Vilen, S.-T.; Lehenkari, P.; Nyberg, P.; Coletta, R.D.; et al. Cathepsin K is present in invasive oral tongue squamous cell carcinoma in vivo and in vitro. PLoS ONE 2013, 8, e70925. [Google Scholar] [CrossRef]
- Khan, I.Z.; Del Guzzo, C.A.; Shao, A.; Cho, J.; Du, R.; Cohen, A.O.; Owens, D.M. The CD200-CD200R axis promotes squamous cell carcinoma. Cancer Res. 2021, 81, 5021–5032. [Google Scholar] [CrossRef]
- Littlewood-Evans, A.; Bilbe, G.; Bowler, W.; Farley, D.; Wlodarski, B.; Kokubo, T.; Gallagher, J.A. The osteoclast-associated protease cathepsin K is expressed in human breast carcinoma. Cancer Res. 1997, 57, 5386–5390. [Google Scholar]
- Brubaker, K.D.; Vessella, K.D.; True, L.D.; Thomas, R.; Corey, E. Cathepsin K mRNA and protein expression in prostate cancer progression. J. Bone Miner. Res. 2003, 18, 222–230. [Google Scholar] [CrossRef]
- Liu, L.; Fan, Y.; Chen, Z.; Zhang, Y.; Yu, J. CaSR Induces Osteoclast Differentiation and Promotes Bone Metastasis in Lung Adenocarcinoma. Front. Oncol. 2020, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Haeckel, C.; Krueger, S.; Kuester, D.; Ostertag, H.; Samii, M.; Buehling, F.; Broemme, D.; Czerniak, B.; Roessner, A. Expression of cathepsin K in chordoma. Hum. Pathol. 2000, 31, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Jaber, O.; Kirby, P. Alveolar Soft Part Sarcoma. Arch. Pathol. Lab. Med. 2015, 11, 1459–1462. [Google Scholar] [CrossRef] [PubMed]
- Ciążyńska, M.; Bednarski, I.; Wódz, K.; Kolano, P.; Narbutt, J.; Sobjanek, M.; Woźniacka, A.; Lesiak, A. Proteins involved in cutaneous basal cell carcinoma development. Oncol. Lett. 2018, 3, 4064–4072. [Google Scholar] [CrossRef]
- Reithmeier, A.; Norgård, M.; Ek-Rylander, B.; Näreoja, T.; Andersson, G. Cathepsin K regulates localization and secretion of Tartrate-Resistant Acid Phosphatase (TRAP) in TRAP-overexpressing MDA-MB-231 breast cancer cells. BMC Mol. Cell Biol. 2020, 21, 15. [Google Scholar] [CrossRef]
- Andrade, S.; Gouvea, I.; Silva, M.; Castro, E.; de Paula, C.; Okamoto, D.; Oliveira, L.; Peres, G.; Ottaiano, T.; Facina, T.; et al. Cathepsin K induces platelet dysfunction and affects cell signaling in breast cancer—Molecularly distinct behavior of cathepsin K in breast cancer. BMC Cancer 2016, 16, 173. [Google Scholar] [CrossRef]
- Wu, N.; Wang, Y.; Wang, K.; Zhong, B.; Liao, Y.; Liang, J.; Jiang, N. Cathepsin K regulates the tumor growth and metastasis by IL-17/CTSK/EMT axis and mediates M2 macrophage polarization in castration-resistant prostate cancer. Cell Death Dis. 2022, 13, 813. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, S.; Wu, T.; Lv, Z.; Ba, J.; Gu, W.; Mu, Y. Expression and clinical significance of Cathepsin K and MMPs in invasive non-functioning pituitary adenomas. Front. Oncol. 2022, 12, 901647. [Google Scholar] [CrossRef]
- Takeshita, S.; Kaji, K.; Kudo, A. Identification and characterization of the new osteoclast progenitor with macrophage phenotypes being able to differentiate into mature osteoclasts. J. Bone Miner Res. 2000, 15, 1477–1488. [Google Scholar] [CrossRef]
- Podgorski, I.; Linebaugh, B.; Koblinski, J.; Rudy, D.; Herroon, M.; Olive, M.; Sloane, B. Bone marrow-derived cathepsin K cleaves SPARC in bone metastasis. Am. J. Pathol. 2009, 175, 1255–1269. [Google Scholar] [CrossRef]
- Herroon, M.; Rajagurubandara, E.; Rudy, D.; Chalasani, A.; Hardaway, A.; Podgorski, I. Macrophage cathepsin K promotes prostate tumor progression in bone. Oncogene 2013, 32, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Haagerup, A.; Hertz, J.M.; Christensen, M.F.; Binderup, H.; Kruse, T.A. Cathepsin K gene mutations and 1q21 haplotypes in at patients with pycnodysostosis in an outbred population. Eur. J. Hum. Genet. 2000, 8, 431–436. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, D.H.; Wang, Z.; He, X.J.; Diplas, B.H.; Yang, R.; Killela, P.J.; Liang, J.; Meng, Q.; Ye, Z.Y.; Wang, W.; et al. A Mutation in CATK Gene in an Autosomal Recessive Pycnodysostosis Family of Chinese Origin. Calcif. Tissue Int. 2015, 96, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Kozawa, E.; Cheng, X.W.; Urakawa, H.; Arai, E.; Yamada, Y.; Kitamura, S.; Sato, K.; Kuzuya, M.; Ishiguro, N.; Nishida, Y. Increased expression and activation of cathepsin K in human osteoarthritic cartilage and synovial tissues. J. Orthop. Res. 2016, 34, 127–134. [Google Scholar] [CrossRef]
- Hao, L.; Zhu, G.; Lu, Y.; Wang, M.; Jules, J.; Zhou, X.; Chen, W. Deficiency of cathepsin K prevents inflammation and bone erosion in rheumatoid arthritis and periodontitis and reveals its shared osteoimmune role. FEBS Lett. 2015, 589, 1331–1339. [Google Scholar] [CrossRef]
- Yamada, H.M. Effects of the cathepsin K Inhibitor ONO-5334 and concomitant use of ONO-5334 with methotrexate on collagen-induced arthritis in cynomolgus monkeys. Int. J. Rheumatol. 2019, 2019, 5710340. [Google Scholar] [CrossRef]
- Yamashita, T.H. Effect of a cathepsin K inhibitor on arthritis and bone mineral density in ovariectomized rats with collagen-induced arthritis. Bone Rep. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Zhang, D.L. The effect of cathepsin K deficiency on airway development and TGF-beta1 degradation. Respir. Res. 2011, 12, 72. [Google Scholar] [CrossRef]
- Calio, A.M. Cathepsin K expression in clear cell “sugar” tumor (PEComa) of the lung. Virchows Arch. 2018, 473, 55–59. [Google Scholar] [CrossRef]
- Chilosi, M.P. Cathepsin-K expression in pulmonary lymphangioleiomyomatosis. Mod. Pathol. 2009, 22, 161–166. [Google Scholar] [CrossRef]
- Platt, M.O.; Ankeny, R.F.; Shi, G.P.; Weiss, D.; Vega, J.D.; Taylor, W.R.; Jo, H. Expression of cathepsin K is regulated by shear stress in cultured endothelial cells and is increased in endothelium in human atherosclerosis. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1479–H1486. [Google Scholar] [CrossRef] [PubMed]
- Funicello, M.; Novelli, M.; Ragni, M.; Vottari, T.; Cocuzza, T.; Soriano-Lopez, J.; Chiellini, C.; Boschi, F.; Marzola, P.; Masiello, P.; et al. Cathepsin K null mice show reduced adiposity during the rapid accumulation of fat stores. PLoS ONE 2007, 2, e683. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.J.; Sun, J.; Liu, J.; Zhang, M.A.; Shi, F.; Darakhshan, M.; Guerre-Millo, G.; Clement, B.G.; Dolgnov, G.; Shi, G.P. Deficiency and Inhibition of cathepsin K Reduce Body Weight Gain and Increase Glucose Metabolism in Mice. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 2202–2208. [Google Scholar] [CrossRef]
- Lendeckel, U.; Kähne, T.; Have, S.; Bukowska, A.; Wolke, C.; Bogerts, B.; Keilhoff, G.; Bernstein, H. Cathepsin K generates enkephalin from beta-endorphin: A new mechanism with possible relevance for schizophrenia. Neurochem. Int. 2009, 54, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, S.; Ogino, I.; Miyajima, M.; Nonaka, S.; Ito, M.; Yasumoto, Y.; Arai, H. Role of cathepsin K in the development of chronic subdural hematoma. Clin. Neurosci. 2017, 45, 343–347. [Google Scholar] [CrossRef]
- Thompson, S.K.; Halbert, S.M.; Bossard, M.J.; Tomaszek, T.A.; Levy, M.A.; Zhao, B.; Smith, W.W.; Abdel-Meguid, S.S.; Janson, C.A.; D’Alessio, K.J.; et al. Design of potent and selective human cathepsin K inhibitors that span the active site. Proc. Natl. Acad. Sci. USA 1997, 94, 14249–14254. [Google Scholar] [CrossRef]
- Law, S.; Panwar, P.; Li, J.; Aguda, A.H.; Jamroz, A.; Guido, R.V.C.; Brömme, D. A composite docking approach for the identification and characterization of ectosteric inhibitors of cathepsin K. PLoS ONE 2017, 12, e0186869. [Google Scholar] [CrossRef]
- Brömme, D.F. Cathepsin K inhibitors for osteoporosis and potential off-target effects. Expert Opin. Investig. Drugs 2009, 18, 585–600. [Google Scholar] [CrossRef]
- Lindström, E.; Rizoska, B.; Henderson, I.; Terelius, Y.; Jerling, M.; Edenius, C.; Grabowska, U. Nonclinical and clinical pharmacological characterization of the potent and selective cathepsin K inhibitor MIV-711. J. Transl. Med. 2018, 16, 125. [Google Scholar] [CrossRef]
- Stroup, G.B.; Kumar, S.; Jerome, C.P. Treatment with a potent cathepsin K inhibitor preserves cortical and trabecular bone mass in ovariectomized monkeys. Calcif. Tissue Int. 2009, 85, 344–355. [Google Scholar] [CrossRef]
- Jerome, C.; Missbach, M.; Gamse, R. Balicatib, a cathepsin K inhibitor, stimulates periosteal bone formation in monkeys. Osteoporos. Int. 2011, 22, 3001–3011. [Google Scholar] [CrossRef] [PubMed]
- Masarachia, P.J.; Pennypacker, B.L.; Pickarski, M.; Scott, K.R.; Wesolowski, G.A.; Smith, S.Y.; Samadfam, R.; Goetzmann, J.E.; Scott, B.B.; Kimmel, D.B.; et al. Odanacatib reduces bone turnover and increases bone mass in the lumbar spine of skeletally mature ovariectomized rhesus monkeys. J. Bone Miner. Res. 2012, 27, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Vashum, Y.; Premsingh, R.; Kottaiswamy, A.; Soma, M.; Padmanaban, A.; Kalaiselvan, P.; Samuel, S. Inhibitory effect of cathepsin K inhibitor (ODN-MK-0822) on invasion, migration and adhesion of human breast cancer cells in vitro. Mol. Biol. Rep. 2021, 48, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.U.; Woo, S.M.; Kim, S.; Park, J.W.; Lee, H.S.; Bae, Y.S.; Kim, S.H.; Im, S.S.; Seo, J.H.; Min, K.; et al. Inhibition of cathepsin K sensitizes oxaliplatin-induced apoptotic cell death by Bax upregulation through OTUB1-mediated p53 stabilization in vitro and in vivo. Oncogene 2022, 41, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Zerbini, C.A.; McClung, M.R. Odanacatib in postmenopausal women with low bone mineral density: A review of current clinical evidence. Ther. Adv. Musculoskelet. Dis. 2013, 5, 199–209. [Google Scholar] [CrossRef]
- Nakamura, T.; Shiraki, M.; Fukunaga, M.; Tomomitsu, T.; Santora, A.C.; Tsai, R.; Fujimoto, G.; Nakagomi, M.; Tsubouchi, H.; Rosenberg, E.; et al. Effect of the cathepsin K inhibitor odanacatib administered once weekly on bone mineral density in Japanese patients with osteoporosis—A double-blind, randomized, dose-finding study. Osteoporos. Int. 2014, 25, 367–376. [Google Scholar] [CrossRef]
- Langdahl, B.; Binkley, N.; Bone, H.; Gilchrist, N.; Resch, H.; Rodriguez Portales, J.; Denker, A.; Lombardi, A.; Le Bailly De Tilleghem, C.; Dasilva, C.; et al. Odanacatib in the treatment of postmenopausal women with low bone mineral density: Five years of continued therapy in a phase 2 study. J. Bone Miner. Res. 2012, 27, 2251–2258. [Google Scholar] [CrossRef]
- Bone, H.G.; Dempster, D.W.; Eisman, J.A.; Greenspan, S.L.; McClung, M.R.; Nakamura, T.; Papapoulos, S.; Shih, W.J.; Rybak-Feiglin, A.; Santora, A.C.; et al. Odanacatib for the treatment of postmenopausal osteoporosis: Development history and design and participant characteristics of LOFT, the Long-Term Odanacatib Fracture Trial. Osteoporos. Int. 2015, 26, 699–712. [Google Scholar] [CrossRef]
- Jensen, A.B.; Wynne, C.; Ramirez, G.; He, W.; Song, Y.; Berd, Y.; Wang, H.; Mehta, A.; Lombardi, A. The cathepsin K inhibitor odanacatib suppresses bone resorption in women with breast cancer and established bone metastases: Results of a 4-week, double-blind, randomized, controlled trial. Clin. Breast Cancer. 2010, 10, 452–458. [Google Scholar] [CrossRef]
- Gu, X.; Peng, Y.; Zhao, Y.; Liang, X.; Tang, Y.; Liu, J. A novel derivative of artemisinin inhibits cell proliferation and metastasis via down-regulation of cathepsin K in breast cancer. Eur. J. Pharmacol. 2019, 858, 172382. [Google Scholar] [CrossRef]
- Benýšek, J.; Buša, M.; Rubešová, P.; Fanfrlík, J.; Lepšík, M.; Brynda, J.; Matoušková, Z.; Bartz, U.; Horn, M.; Gütschow, M.; et al. Highly potent inhibitors of cathepsin K with a differently positioned cyanohydrazide warhead: Structural analysis of binding mode to mature and zymogen-like enzymes. J. Enzyme Inhib. Med. Chem. 2022, 37, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chen, W.; Hao, L.; Zhu, G.; Feng, S.; Ci, H.; Zhou, X.; Stashenko, P.; Li, Y.P. Inhibiting periapical lesions through AAV-RNAi silencing of cathepsin K. J. Dent. Res. 2013, 92, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gao, B.; Hao, L.; Zhu, G.; Jules, J.; MacDougall, M.J.; Wang, J.; Han, X.; Zhou, X.; Li, Y.P. The silencing of cathepsin K used in gene therapy for periodontal disease reveals the role of cathepsin K in chronic infection and inflammation. J. Periodontal. Res. 2016, 51, 647–660. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yin, W.; Yang, Q.; Ren, J.; Tan, L.; Wang, J.; Liu, J.; Lu, Q.; Ding, H.; Zhan, W.; et al. Inhibition of Cathepsin K Alleviates Autophagy-Related Inflammation in Periodontitis-Aggravating Arthritis. Infect. Immun. 2020, 88, e00498-20. [Google Scholar] [CrossRef]
- Rünger, T.M.; Quintanilla-Dieck, M.J.; Bhawan, J. Role of cathepsin K in the turnover of the dermal extracellular matrix during scar formation. J. Investig. Dermatol. 2007, 127, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Kozloff, K.M.; Quinti, L.; Patntirapong, S.; Hauschka, P.V.; Tung, C.H.; Weissleder, R.; Mahmood, U. Non-invasive optical detection of cathepsin K-mediated fluorescence reveals osteoclast activity in vitro and in vivo. Bone 2009, 44, 190–198. [Google Scholar] [CrossRef]
- Ranade, H.; Paliwal, P.; Chaudhary, A.A.; Piplani, S.; Rudayni, H.A.; Al-Zharani, M.; Niraj, R.R.; Datta, M. Predicting Diagnostic Potential of Cathepsin in Epithelial Ovarian Cancer: A Design Validated by Computational, Biophysical and Electrochemical Data. Biomolecules 2021, 12, 53. [Google Scholar] [CrossRef]
- Zhang, M.; Hu, W.; Cai, C.; Wu, Y.; Li, J.; Dong, S. Advanced application of stimuli-responsive drug delivery system for inflammatory arthritis treatment. Mater. Today Bio 2022, 14, 100223. [Google Scholar] [CrossRef]
- Bolignano, D.; Greco, M.; Arcidiacono, M.; Tripolino, O.; Vita, C.; Provenzano, M.; Donato, C.; Chiarella, S.; Fuiano, G.; Sarro, G.; et al. Cathepsin-K is a potential cardiovascular risk biomarker in prevalent hemodialysis patients. Int. Urol. Nephrol. 2021, 53, 171–175. [Google Scholar] [CrossRef]
- Izumi, Y.; Hayashi, M.; Morimoto, R.; Cheng, X.W.; Wu, H.; Ishii, H.; Yasuda, Y.; Yoshikawa, D.; Izawa, H.; Matsuo, S.; et al. Impact of circulating cathepsin K on the coronary calcification and the clinical outcome in chronic kidney disease patients. Heart Vessel. 2016, 31, 6–14. [Google Scholar] [CrossRef]
- Siddiqui, J.A.; Seshacharyulu, P.; Muniyan, S.; Pothuraju, R.; Khan, P.; Vengoji, R.; Chaudhary, S.; Maurya, S.M.; Lele, S.M.; Jain, M.; et al. GDF15 promotes prostate cancer bone metastasis and colonization through osteoblastic CCL2 and RANKL activation. Bone Res. 2022, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Oso, B.J.; Adeoye, A.O.; Olaoye, I.F. Pharmacoinformatics and hypothetical studies on allicin, curcumin, and gingerol as potential candidates against COVID-19-associated proteases. J. Biomol. Struct. Dyn. 2022, 40, 389–400. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tissue | Disease | Involvement of CatK |
|---|---|---|
| Bone | Pycnodysostosis (PD) | A homozygous or compound heterozygous mutation (mutations that occur on different copies of genes and may completely “knock-out” gene function) in the CatK gene 236G > A, 121–1G > A and 926T > C, causes lower degradation of type I collagen due to lack of CatK activity [94,95]. |
| Postmenopausal osteoporosis (PO) | In a combination of hyper-active osteoclasts and less functional osteoblasts (typically for estrogen-deficient women), CatK inhibitors can target the resorption process [54]. | |
| Osteoarthritis (OA) | The up-regulated expression of the CatK collagenase activity could affect the cartilage matrix degradation in the late stadium of osteoarthritis [96]. | |
| Rheumatoid arthritis (RA) | CatK inhibition can play a significant role in cartilage degradation, retarding the bone loss process and joint destruction [97,98,99]. | |
| Gaucher Disease (GD) | A multisystemic disorder, associated with progressive accumulation of Gaucher cells—large macrophages that store glucocerebroside. In GD patients elevated CatK levels are observed, which can lead to osteoporosis and lytic bone lesions [56]. | |
| Skin | Psoriasis (PS) | Psoriatic lesions (an increased copy number of variations in β-defensin gene locus) express an elevated level of CatK compared to healthy skin [65]. |
| Scar formation (SF) | In surgical scars compared to normal skin, is observed a proteolytic activity of CatK [57]. | |
| Lung | Lung fibrosis (LF) | CatK protects against matrix deposition in bleomycin-induced LF [100]. |
| Pulmonary lymphangioleiomyomatosis (PLAM) | General expression of CatK activity was observed during PLAM [101,102]. | |
| Atherosclerosis (AS) | Stress-induced—CatK expression in endothelial cells can lead to vascular remodeling and atherosclerosis [103]. | |
| Adipose tissue | Obesity/overweight | CatK expression can be detected in pre-adipocytes and be further up-regulated in the process of differentiation. The lack of CatK retards the adipogenesis [104,105]. |
| Central nervous system | Schizophrenia | High levels of CatK in individuals suffering from schizophrenia, as a result of long-term treatment of the patients with neuroleptics [68,106]. |
| Cerebral aneurysm (CA) | Cystein cathepsins cause degradation of ECM in aneurysmal wall in the late state of CA. The administration of cysteine proteases inhibitors (CPIs) leads to prevention of CA progression [67]. | |
| Chronic subdural hematoma (CSH) | The expression of CatK in CSH patients, may lead to CSH development [107]. | |
| Cancer | Adenocarcinoma Chondrosarcoma Renal and extrarenal perivascular epithelioid tumors (PEComas) Alveolar soft part sarcoma Basal cell carcinoma Oral tongue squamous carcinoma cells Breast and prostate cancer Other epithelial-derived cell cancers | High levels of cathepsin K have been observed in metastatic tumors, which correlates with its primary function to degrade collagen, thus aiding tumor invasion and metastasis [82]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijanović, O.; Jakovleva, A.; Branković, A.; Zdravkova, K.; Pualic, M.; Belozerskaya, T.A.; Nikitkina, A.I.; Parodi, A.; Zamyatnin, A.A., Jr. Cathepsin K in Pathological Conditions and New Therapeutic and Diagnostic Perspectives. Int. J. Mol. Sci. 2022, 23, 13762. https://doi.org/10.3390/ijms232213762
Mijanović O, Jakovleva A, Branković A, Zdravkova K, Pualic M, Belozerskaya TA, Nikitkina AI, Parodi A, Zamyatnin AA Jr. Cathepsin K in Pathological Conditions and New Therapeutic and Diagnostic Perspectives. International Journal of Molecular Sciences. 2022; 23(22):13762. https://doi.org/10.3390/ijms232213762
Chicago/Turabian StyleMijanović, Olja, Aleksandra Jakovleva, Ana Branković, Kristina Zdravkova, Milena Pualic, Tatiana A. Belozerskaya, Angelina I. Nikitkina, Alessandro Parodi, and Andrey A. Zamyatnin, Jr. 2022. "Cathepsin K in Pathological Conditions and New Therapeutic and Diagnostic Perspectives" International Journal of Molecular Sciences 23, no. 22: 13762. https://doi.org/10.3390/ijms232213762
APA StyleMijanović, O., Jakovleva, A., Branković, A., Zdravkova, K., Pualic, M., Belozerskaya, T. A., Nikitkina, A. I., Parodi, A., & Zamyatnin, A. A., Jr. (2022). Cathepsin K in Pathological Conditions and New Therapeutic and Diagnostic Perspectives. International Journal of Molecular Sciences, 23(22), 13762. https://doi.org/10.3390/ijms232213762