Abstract
Pineapple (Ananas comosus L.) is an important fruit crop in tropical regions, and it requires efficient sugar allocation during fruit development. Sugars Will Eventually be Exported Transporters (SWEETs) are a group of novel sugar transporters which play critical roles in seed and fruit development. However, the function of AcSWEETs remains unknown in the sugar accumulation. Herein, 17 AcSWEETs were isolated and unevenly located in 11 chromosomes. Analysis of a phylogenetic tree indicated that 17 genes were classified into four clades, and the majority of AcSWEETs in each clade shared similar conserved motifs and gene structures. Tissue-specific gene expression showed that expression profiles of AcSWEETs displayed differences in different tissues and five AcSWEETs were strongly expressed during fruit development. AcSWEET11 was highly expressed in the stage of mature fruits in ‘Tainong16’ and ‘Comte de paris’, which indicates that AcSWEET11 was important to fruit development. Subcellular localization analysis showed that AcSWEET11 was located in the cell membrane. Notably, overexpression of AcSWEET11 could improve sugar accumulation in pineapple callus and transgenic tomato, which suggests that AcSWEET11 might positively contribute to sugar accumulation in pineapple fruit development. These results may provide insights to enhance sugar accumulation in fruit, thus improving pineapple quality in the future.
1. Introduction
Carbohydrates are not only vital to supplying necessary energy during plant growth and development, but they also determine fruit quality and flavour [1]. Carbohydrate in plants are produced in the chloroplast by photosynthesis, and it is then translocated to the sink organs by the long-distance transport system of phloem [2]. The symplasmic pathway, the apoplasmic pathway and the alternation between apoplasmic and symplasmic unloading are potential ways for phloem uploading. However, the apoplasmic phloem uploading needs the assistance of specific sugar transporters. Three types of sugar transporters have been identified to date, including monosaccharide transporters [3], sucrose transporters [4], and Sugars Will Eventually be Exported Transporter (SWEET) proteins [5].
The SWEET family members are the novel proteins of carbohydrate transporters in plants, and they are featured by the MtN3/Saliva (PF03083) motif (known as PQ-loop repeat) with seven alpha-helical transmembrane domains connected by a PQ-loop repeat [6]. SWEETs are comprised of a 3–1–3 TMH structure based on the tandem repeats of two 3-TMH domains by a single transmembrane domain [7]. To date, genome-wide analysis of SWEET genes has identified their presence in plants, including 17 in Arabidopsis thaliana [6] (Chen et al., 2010), 21 in rice (Oryza sativa) [8], 52 in soybean (Glycine max) [9], 33 in apple (Malus domestica) [10], 25 in banana (Musa acuminata L.) [11], and 17 in grape (Vitis vinifera) [12]. SWEETs could be divided into four groups (clades I–IV) [6,8] which have many different functions. Clades I and II are mainly transported hexoses, whilst Clade III comprises transported sucrose [12,13]. Clade IV consists of the vacuolar transporters, which regulate the flux of fructose amongst the tonoplast [14].
SWEETs play an important role in accumulation and transportation of sugar. SWEET could change the sugar composition to improve the sugar accumulation in fruit. In tomato, SlSWEET1a is highly expressed in young leaves and regulates glucose accumulation. Glucose contents are reduced after the overexpression of SlSWEET1a, and up-regulating the ratio of fructose/glucose in the developing fruit modifies the sugar accumulation pattern [15]. Similarly, the overexpression of VvSWEET10 could increase the contents of glucose and fructose, which improves the total contents in grape [16]. In pear, PuSWEET15 increased the content of sucrose after overexpression in fruit, whilst the content of sucrose was decreased after silencing [17]. SWEET uploads carbohydrates from the source to the sink via the transport system of phloem, which regulates sugar accumulation for the fruit. Ko et al. [18] found that SlSWEET15 mediates efflux of sucrose from phloem cells to the fruit apoplasm and then imports it into the storage parenchyma cells during tomato fruit development. Similar results were found in soybean, where GmSWEET15 exports the sucrose from the endosperm to the embryo during seed development [19]. Moreover, AcSWEET2a/2b and AcSWEET16b in carambola (Averrhoa carambola L.). [20] and MdSWEET9b and MdSWEET15a in apple [21] could participate in sugar accumulation during fruit development.
Pineapple is an important fruit in tropical regions. Sugar composition and content determine the quality of pineapple fruit. Hao et al. [22] isolated AcSWEETs and then analyzed the expression of fruit development by transcriptome; they speculated that AcSWEETs might play an important role in fruit development. However, the function of AcSWEETs remains largely unknown in sugar accumulation during fruit development. In this study, 17 AcSWEETs were isolated, and the expression patterns of these genes in different tissues were analyzed. AcSWEET11 was screened out after we analyzed the expression of these genes during fruit development. The function of AcSWEET11 in sugar content was then demonstrated by the transformation of pineapple callus and tomatoes. Our study will not only contribute to understanding the mechanism of sugar accumulation in fruit crops but also will be helpful in improving the quality of pineapple in the future.
2. Results
2.1. Identification of AcSWEET Family Genes in Pineapple
A total of 17 and 21 SWEET proteins in Arabidopsis and rice, respectively, were obtained from the Unirpot database and were blasted in the pineapple genome (https://genomevolution.org/CoGe/NotebookView.pl?nid=937 (accessed on 3 August 2022)). The conserved domain was validated by NCBI-CDD to verify the AcSWEETs. The results showed that all the AcSWEETs contained two MtN3/saliva or PQ-loop superfamily (Figure 1A). A total of 17 AcSWEETs were identified and unevenly distributed on 11 chromosomes (Table 1, Figure 1B). These genes were named on the basis of Arabidopsis homologs. The open reading frame (ORF) lengths of AcSWEETs ranged from 579 bp (AcSWEET2) to 906 bp (AcSWEET12), and their protein length varied from 193 to 302 amino acids. The molecular weights were between 22.29 kDa (AcSWEET2) and 33.36 kDa (AcSWEET12). The theoretical isoelectric point (pI) ranged from 6.53 (AcSWEET12) to 9.75 (AcSWEET5), and most of the pI were more than 8.5 except for AcSWEET2, AcSWEET12 and AcSWEET14. The transmembrane regions of AcSWEET were predicted by TMHMM software. The results showed that 13 members of the encoded protein contained seven transmembranes, and two members harbored six transmembranes, whilst two members only had three transmembranes (Figure S1). Further analysis of putative subcellular localization showed that most of the genes were located in the cell membrane, except AcSWEET5 and AcSWEET13, which were located in the nucleus and chloroplast, respectively. Moreover, AcSWEET15 was located not only in the cell membrane but also in the golgi apparatus (Table 1). These results indicated that AcSWEETs may have a membrane protein function.
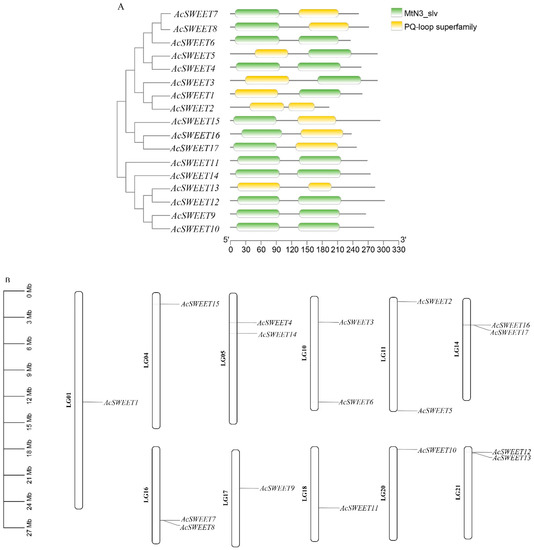
Figure 1.
Conserved domain analysis of AcSWEETs and chromosomal location. (A) Different domains are represented in different colors along with the conservative domain of prediction as shown in the legend. (B) Location of AcC5-MTase genes on the chromosomes.

Table 1.
Overview of AcSWEET genes identified in pineapple.
2.2. Phylogenetic Tree Analysis of SWEET in Pineapple, Rice and Arabidopsis
A phylogenetic tree of 17, 17 and 21 SWEETs was constructed using MEGA6 software to study the evolutionary relationships of SWEET proteins amongst pineapple, Arabidopsis and rice, respectively. The phylogenetic tree showed that these genes were classified into four clades (I–IV) (Figure 2 and Figure 3A). Clades II and III in Arabidopsis and rice contained a large number of SWEET family members, with five and six, respectively. In the meantime, clades I and IV had three members. Most SWEETs were grouped into the same branches between rice and pineapple, which implied that the evolutionary relationships of rice and pineapple were close.
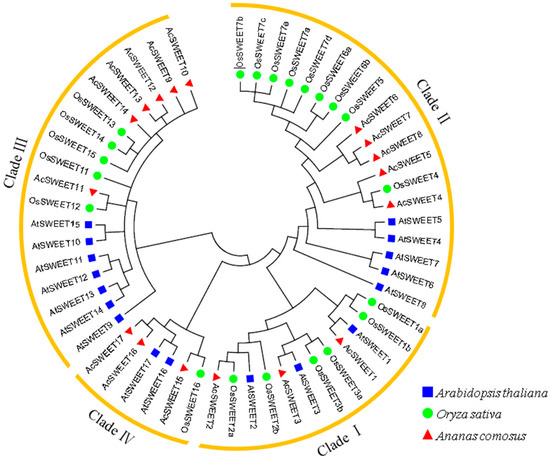
Figure 2.
Phylogenetic tree of SWEETs from Arabidopsis, rice and pineapple. We used the neighbor-joining method to draw phylogenetic the tree with MEGA6.0 with 1000 bootstraps replicates. Four subgroups were divided as Clades I–IV.
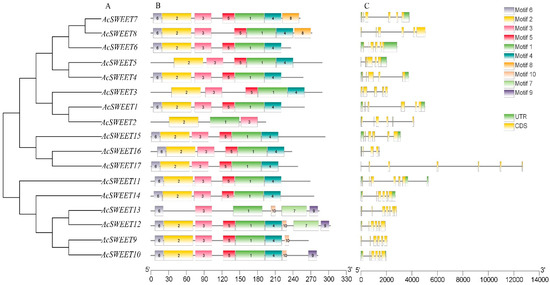
Figure 3.
Evolutionary, gene structural and motif analyses of AcSWEETs in pineapple. (A) phylogenetic tree of candidate amino acid sequences was constructed using the neighbor-joining method with 1,000 bootstraps in MEGA 6.0. (B) The conserved motifs of AcSWEETs proteins were analyzed using the MEME tool. (C) The gene structure of AcSWEETs was identified by TBtools.
2.3. Analysis of Gene Structural and Conserved Motifs in AcSWEETs
The analysis of intron–exon structures showed that the numbers of intron–exon were highly conserved in AcSWEETs. All AcSWEETs contained several exons, which varied from five to seven. A total of 12 members of AcSWEETs harbored six exons. AcSWEET2, AcSWEET3 and AcSWEET16 contained five exons. AcSWEET3 and AcSWEET17 displayed seven exons. Interestingly, AcSWEET17 had a special structure, with six axons composed of two long introns and four short introns (Figure 3C). A total of 10 motifs were identified in AcSWEETs using MEME. Three to nine motifs were detected in AcSWEETs. AcSWEET2 had the shortest and least amino acid sequence with three motifs. By contrast, AcSWEET13 had the longest sequence, with nine motifs. Motif1 and motif3 were observed in all AcSWEETs. Motif2, motif4 and motif5 were present in all AcSWEETs except AcSWEET2 and AcSWEET13. Motif10 was the specific motif in AcSWEET9, AcSWEET10, AcSWEET12 and AcSWEET13, which were classified in the same clade (Figure 3B).
2.4. Expression Profiles of AcSWEETs in Different Pineapple Tissues
The expression levels of AcSWEETs were analyzed in root, stem, leaf, flower, carpopodium, mature fruit and carpodermis by qRT-PCR to understand the potential functions of these genes (Figure 4). These results indicated that AcSWEET1, AcSWEET10 and AcSWEET13 were highly expressed in root. AcSWEET4 and AcSWEET16 were highly expressed not only in root but also in carpopodium. The expression levels of AcSWEET2 and AcSWEET15 were higher in flower than in other tissues. AcSWEET3, AcSWEET7, AcSWEET11, AcSWEET12 and AcSWEET14 were strongly expressed in fruit. AcSWEET5 was predominantly expressed in the carpodermis. Interestingly, all AcSWEETs had lower expression in leaf and stem. These results indicated that AcSWEETs may be involved in the unloading of pineapple.
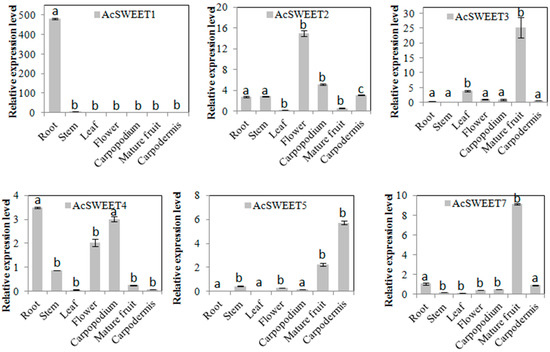
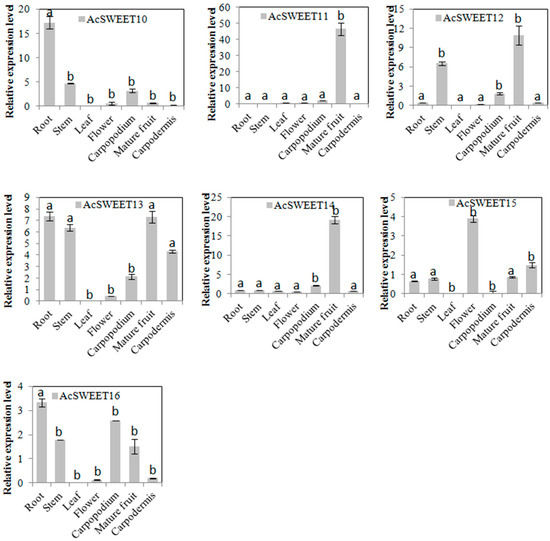
Figure 4.
Tissue-specific expression profiles of AcSWEETs in pineapple. Values represented as mean of three biological replicates, and standard deviation bars represented as means ± SD (n = 3). Different letters indicate significant differences at p < 0.05.
2.5. Expression Patterns of AcSWEETs during Pineapple Fruit Development
AcSWEETs significantly affected plant development by regulating carbohydrate transport. AcSWEETs were expressed in fruits during development at 0, 7, 21, 35, 49, 63 and 77 days after anthesis. AcSWEET9 and AcSWEET13 exhibited high expression during fruit development, whilst AcSWEET1, AcSWEET2, AcSWEET4 and AcSWEET15 exhibited low expression. AcSWEET3, AcSWEET5, AcSWEET7, AcSWEET10, AcSWEET12 and AcSWEET16 exhibited high expression at 0 and 21 days, but they showed low expression during pineapple fruit development. In the meantime, AcSWEET11 and AcSWEET14 had the lowest expression at 0 days, and then their expression increased during fruit development (Figure 5). Further analysis of two pineapple varieties revealed that AcSWEET11 showed high expression in the stage of mature fruits in ‘Tainong16’ and ‘Comte de paris’ (Figure S2). These results indicated that AcSWEET11 might play a critical role in pineapple fruit development.
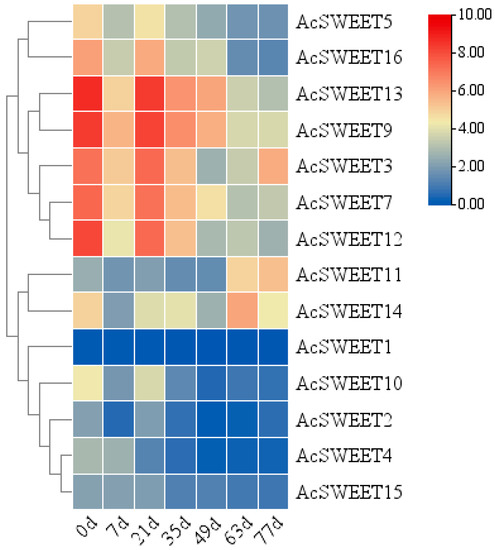
Figure 5.
Expression analysis of AcSWEETs during ‘Comte de paris’ fruit development conducted by qRT-PCR. d, days after anthesis.
2.6. Subcellular Localization of AcSWEET11 Protein
The coding sequence of AcSWEET11 was fused with EGFP driven by 35S promoter to explore the subcellular localization of the AcSWEET11, and the gene was transiently expressed in the cells of Nicotiana benthamiana. The transient expression of AcSWEET11 showed that the GFP fluorescence was specifically observed in the cell membrane (Figure 6). This result indicated that the function of AcSWEET11 was predominantly at the membrane of plant cells.
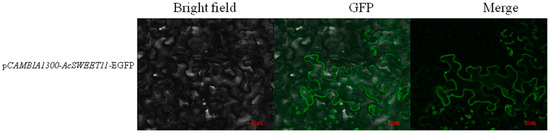
Figure 6.
Subcellular localization analysis of AcSWEET11 in Nicotiana benthaminana leaves after 2 days of agrobacterium injection. Scale bars = 20 μm.
2.7. Overexpression of AcSWEET11 Enhances the Accumulation of Soluble Sugar
The pCAMBIA1300-AcSWEET11-EGFP vector was transformed into an embryogenic callus of pineapple by agrobacterium -mediated vacuum infiltration to further confirm the function of AcSWEET11. The callus was used to detect the soluble sugar after co-cultivation for 3 days. The result showed that transient callus (18.01 mg/g FW) had 1.4 times the content of soluble sugar as a normal callus (12.81 mg/g FW). The expression of transient calluses were significantly higher than normal calluses (Figure S3). Therefore, AcSWEET11 plays an essential role in sugar accumulation of pineapple.
The pCAMBIA1300-AcSWEET11-EGFP constructor was transferred into micro-tom tomatoes to gain further insight into the function of AcSWEET11. Independent transgenic lines (15-1) were obtained from hygromycin B resistance selection with genomic PCR detection. The result showed that soluble sugar contents had significant differences between transgenic lines (15-1) and WT in green and mature fruits. The content of green fruit was 9.07 mg/g FW in WT, whilst it was 16.51 mg/g FW in 15-1, which was 1.8 times as many as that of WT. In mature fruits, the content of 15-1 was 20.33 mg/g FW, which was significantly higher than that of WT (11.63 mg/g FW). Intriguingly, green fruit of 15-1 accumulated significantly more soluble sugar than WT mature fruits (Figure 7). These results suggest that AcSWEET11 enhanced soluble sugar accumulation in fruit.
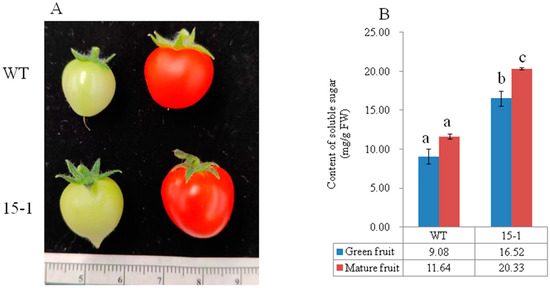
Figure 7.
Overexpression of AcSWEET11 improved soluble sugar in micro-tom tomato. (A) Development of tomato fruit in WT and transgenic tomato lines (15-1). (B) Soluble sugar content of WT and transgenic tomato lines (15-1). Blue columns presented in green fruit, red columns presented in mature fruit. Values denoted three biological replicates, and vertical lines represented as mean of SD. Different letters denoted significant difference with one-way ANOVA test by Tukey’s test (p < 0.05).
3. Discussion
Carbohydrates play an important role in plant growth and development [16,18], and their allocation and accumulation determine the fruit quality during development [1]. SWEET genes are a novel class of sugar transporters that move sugars between tissues by the phloem [23]. Hitherto, genome-wide analysis showed that SWEETs were separated in many plants, such as rice [8], apples [10], banana [11], tomato [15], cucumbers [20] and longan [24]. In our study, 17 SWEETs were identified from the pineapple genome, which is different from a previous report that isolated 18 in MD-2. This difference is likely due to the different criteria used in identifying the candidate genes. Hao et al. [22] believed that if only the genes have MtN3/saliva and PQ-loop domains, they will belong to the SWEET family. In the present study, the candidate members contained either of two MtN3/saliva or the PQ-loop structural domain, and the lengths of amino acids were from 100 aa to 400 aa from the MD-2 genome. Aco006158.1 contained MtN3/saliva and PQ-loop domains in addition to the PQQ-DH-like superfamily, and the length of amino acids was 885 aa, which was larger than that of SWEET members. Therefore, Aco006158.1 did not belong to the SWEET family. Furthermore, Hao et al. [22] found 21 members in F153, which shows that the difference in the numbers of genes is due to the genome of different varieties.
SWEET proteins play key roles in plant growth. Chen et al. [25] and Guo et al. [26] have found that AtSWEET2 and AtSWEET17 were highly expressed in root, but they perform a different function. Further study showed that AtSWEET2 limits carbon sequestration in root, whilst AtSWEET17 was a fructose uniporter in the root of Arabidopsis. Similarly, the interaction between StSWEET11 and StSP6A triggers the efflux of sucrose to regulate tuber formation of potato [27]. In our study, AcSWEET1, AcSWEET4, AcSWEET10, AcSWEET13 and AcSWEET16 were highly expressed in root (Figure 4), suggesting that these genes may have effected on sugar transported in pineapple root. SWEET not only affects root formation but also plays an important role in floral organ development and flowering time. In Jasminum sambac, SWEET5 is upregulated during flower opening and sexual reproduction [28]. In turn, the overexpression of AtSWEET10 can significantly accelerate flowering in Arabidopsis [29]. OsSWEET11 affects pollen development [30], whilst AtSWEET15 is expressed during pollen maturation and germination [31]. In our study, AcSWEET2 and AcSWEE15 were more highly expressed in flower compared to other tissues, which indicates their role in flower formation or florescence (Figure 4). Moreover, AcSWEET3, AcSWEET7, AcSWEET11 and AcSWEET14 were strongly expressed in fruit (Figure 4). Similar results were found in pears [17], loquats [32] and grapes [16], which imply their key role in pineapple fruit. Further analysis of expression in pineapple fruit development demonstrated that AcSWEET3 and AcSWEET7 were highly expressed at 0 days, and they then decreased with fruit development. In the meantime, the expression levels of AcSWEET11 and AcSWEET14 increased. The expression in two varieties exhibited that AcSWEET11 played a central role during pineapple fruit development (Figure S2). Interestingly, SWEET is involved in phloem loading from the source leaves [6]. However, in our study, AcSWEETs had low expression in leaf and stem. These results suggest that AcSWEETs transported sugars, and might work together with other sugar transporters in pineapple, such as sucrose carriers. Moreover, AcSWEET6, AcSWEET8, AcSWEET9 and AcSWEET17 have not yet been detected in specific tissues including root, leaf, stem, flower, carpopodium, carpodermis, and mature fruit; these results indicate that AcSWEET6, AcSWEET8, AcSWEET9 and AcSWEET17 might be expressed in other tissues of pineapple.
The ability of transmembrane transport would determine sugar accumulation in fruit. SWEET is one of the most important sugar transporters, and it can transport sucrose, glucose and fructose [5,33,34]. Several reports have shown that SWEET improved sugar contents in different plants, such as apple [10], banana [11] and grape [16]. Recent studies have found that SWEET mediated sugar efflux across the cell membrane following the concentration gradient without supplementary energy [35]. SlSWEET15 regulates sucrose efflux from phloem cells to the fruit apoplasm, and then it imports it into the storage parenchyma cells during fruit development in tomato [18]. In grape, the levels of glucose, fructose and total sugar increased significantly after the overexpression of VvSWEET10 in grapevine calli and tomatoes, which implied that VvSWEET10 could improve sugar accumulation [16]. Further study demonstrated that sucrose/H+ exchangers may efflux cytosolic sugars in fruit to alleviate the demand for supplementary energy of sugar transport at the plasma membrane [36]. These results suggest that SWEETs regulated sugar efflux from cell membranes to control sugar accumulation in fruit development. Herein, AcSWEET11 was located in the cell membrane, whilst sugar contents were enhanced after the overexpression of AcSWEET11 in pineapple callus and tomatoes. These results indicate that AcSWEET11 may regulate sugar transportation and subsequently lead to sugar accumulation in fruit.
4. Materials and Methods
4.1. Plant Materials
‘Comte de paris’ and ‘Tainong16’ were cultured in the South Subtropical Crop Research Institute of the Chinese Academy of Tropical Agricultural Sciences (21_1002” N; 110_16034” E). Different tissues were collected including root, leaf, stem, flower, carpopodium, carpodermis, young fruit, immature fruit and mature fruit, and they were stored at −80 °C. The induction of embryogenic callus were conducted following Lin et al. [37].
4.2. Genome-Wide Identification of SWEET Family in Pineapple
The protein sequences of SWEET in Arabidopsis and rice were downloaded from the Unirpot database (https://www.uniprot.org/uniprotkb?facets=model_organism%3A3702%2Creviewed%3Atrue&query=sweet (accessed on 3 August 2022)). Related information of SWEETs is shown in Supplementary Table S1. The candidate protein sequences were identified from pineapple genome based on a homolog of Arabidopsis SWEET sequences by TBtools [38]. Thereafter, the conserved domain database (NCBI-CDD, https://www.ncbi.nlm.nih.gov/cdd (accessed on 11 August 2022)) was used to confirm whether the candidate proteins have one or more MtN3/saliva. Subsequently, these sequences were scanned by TBtools to visualise the common conservative motif. Ultimately, 17 AcSWEETs were identified (Table 1). These AcSWEETs in pineapple were named according to their phylogenetic relationships to the members of Arabidopsis. The molecular weight (Mw) and theoretical (pI) of AcSWEET proteins were predicted by the Expasy tool (https://www.expasy.org/ (accessed on 20 August 2022)). Subcellular localization was predicted by Plant-mPLoc (http://www.csbio.sjtu.edu.cn/cgi-bin/PlantmPLoc.cgi (accessed on 21 August 2022)). Chromosomal localization was drawn by TBtools [38] based on the starting and stop positions of all SWEETs in pineapple. The transmembrane helices of AcSWEET protein sequences were predicted by TMHMM (http://www.cbs.dtu.dk/services/TMHMM/ (accessed on 24 August 2022)).
4.3. Analysis of Conserved Motif and Gene Structure
The conserved motifs of 17 AcSWEETs were analysed using the MEME software version 5.3.2 (https://meme-suite.org/meme/meme_5.3.2/tools/meme (accessed on 25 August 2022)). Gene structures were analyzed according to the exon/intron of AcSWEETs. The schematics of conserved motifs and gene structures were visualized using TBtools software.
4.4. Sequences Alignment and Phylogenetic Tree Analysis
The amino sequences of SWEET genes in pineapple, Arabidopsis and rice were obtained for multiple alignments using the MUSCLE alignment module in MEGA6 with default parameters. The data matrix of alignment was used to generate a phylogenetic tree with the value of 1000 bp replicates by the neighbor-joining method [39].
4.5. Analysis of Gene Expression
Total RNA extraction and qRT-PCR were conducted following Lin [40]. qRT-PCR was conducted on a LightCycler® 480 II (Roche, Basel, Switzerland) using SYBR Green qPCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA). Specific primers were designed using Primer-BLAST programme (https://www.ncbi.nlm.nih.gov/tools/primer- blast/ (accessed on 13 February 2019)) of NCBI. The primer sequences were listed in Supplementary Table S2. Relative gene expression was calculated using the 2−ΔΔCT method with the pineapple β-actin gene as the internal reference gene. All analyses were performed in three biological replicates.
4.6. Subcellular Localization of AcSWEET11
The CDS of AcSWEET11 was conducted into pCAMBIA1300-35S-EGFP without a stop condone by BamH I and Sal I digestion. Subsequently, the plasmid pCAMBIA1300-AcSWEET11-EGFP was transferred into A. tumefaciens strain EHA105 and was injected into tobacco leaves. After 2 days, the GFP fluorescence of these tobacco leaves was observed by confocal laser scanning microscopy. The primers of pCAMBIA1300-35S-EGFP vector are provided in Table S3.
4.7. AcSWEET11 Overexpression in Pineapple Embryogenic Callus and Tomato
The plasmid pCAMBIA1300-AcSWEET11-EGFP underwent transient transformation into embryogenic callus of pineapple by agrobacterium vacuum infiltration following Luan [41]. After 3 days of co-culture, the sugar contents of these callus were detected.
Plasmid pCAMBIA1300-AcSWEET11-EGFP was transformed into tomatoes by agrobacterium strain EHA105 to probe the function of AcSWEET11 in fruit. The plants were generated following Sun et al. [42]. The transgenic tomato plants were identified by detection with the primers of hygromycin (Table S3). Transgenic tomato plant T1 lines named as 15-1 were obtained for further analysis.
4.8. Analysis of Sugar Content
The total soluble sugar of tomato fruits and pineapple callus were analyzed using the anthrone-sulphuric acid colorimetry method [43]. Data are presented as the mean of ± SDs. Significant analysis was carried out using SPSS software (version 16.0) and one-way ANOVA test with a Tukey’s test (p < 0.05).
5. Conclusions
A total of 17 AcSWEETs were identified in the pineapple genome, and they were clustered into four clades. Tissue-specific expression showed that AcSWEET genes may play different roles in the development of pineapple tissues and that AcSWEET11 might be involved in fruit development. Furthermore, the overexpression of AcSWEET11 has improved sugar accumulation in pineapple callus and transgenic tomato. This work will contribute to understanding the functional characteristics of AcSWEETs and improving the quality of pineapple.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232213875/s1
Author Contributions
Methodology, S.L. and Y.Y. (Yumei Yang); software, Y.P.; resources Q.W.; data curation, Y.Y. (Yanli Yao); writing—original draft preparation and funding acquisition, W.L.; writing—review and editing, W.S.; project administration, X.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by the National Key R&D Program of China (No. 2019YFD1000500), National Natural Science Foundation of China (No. 32001989), and the Natural Science Foundation of Hainan Province (No. 321QN302).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We wish to thank Xi’ou Xiao of South Subtropical Crops Research Institute of CATAS for providing research strategy.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Wen, S.Y.; Neuhaus, H.E.; Cheng, J.T.; Bie, Z.L. Contributions of sugar transporters to crop yield and fruit quality. J. Exp. Bot. 2022, 73, 2275–2289. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhang, Z.; Li, B.; Qin, G.; Tian, S. Molecular basis for optimizing sugar metabolism and transport during fruit development. aBIOTECH 2021, 2, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Slewinski, T.L. Diverse functional roles of monosaccharide transporters and their homologs in vascular plants: A physiological perspective. Mol. Plant 2011, 4, 641–662. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Qu, X.Q.; Hou, B.H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Hou, B.H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.Q.; Guo, W.; Kim, J.G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef]
- Baker, R.F.; Leach, K.A.; Braun, D.M. SWEET as sugar, new sucrose effluxers in plants. Mol. Plant 2012, 5, 766–768. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, S. Rice MtN3/saliva/SWEET family genes and their homologs in cellular organisms. Mol. Plant 2013, 6, 665–674. [Google Scholar] [CrossRef]
- Patil, G.; Valliyodan, B.; Deshmukh, R.; Prince, S.; Nicander, B.; Zhao, M.Z.; Sonah, H.; Song, L.; Lin, L.; Chaudhary, J.; et al. Nguyen Soybean (Glycine max) SWEET gene family: Insights through comparative genomics, transcriptome profiling and whole genome re-sequence analysis. BMC Genom. 2015, 16, 520. [Google Scholar] [CrossRef]
- Wei, X.; Liu, F.; Chen, C.; Ma, F.; Li, M. The Malus domestica sugar transporter gene family: Identifications based on genome and expression profiling related to the accumulation of fruit sugars. Front. Plant Sci. 2014, 5, 569. [Google Scholar] [CrossRef]
- Miao, H.X.; Sun, P.G.; Liu, Q.; Miao, Y.L.; Liu, J.H.; Zhang, K.X.; Hu, W.; Zhang, J.B.; Wang, J.Y.; Wang, Z.; et al. Genome-wide analyses of SWEET family proteins reveal involvement in fruit development and abiotic/biotic stress responses in banana. Sci. Rep. 2017, 7, 3536. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Piron, M.C.; Meyer, S.; Merdinoglu, D.; Bertsch, C.; Mestre, P. The SWEET family of sugar transporters in grapevine: Vv SWEET4 is involved in the interaction with Botrytis cinerea. J. Exp. Bot. 2014, 65, 6589–6601. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, L.; Huang, W.; Yuan, M.; Zhou, F.; Li, X.; Lin, Y. Overexpression of Os SWEET5 in rice causes growth retardation and precocious senescence. PLoS ONE 2014, 9, e94210. [Google Scholar]
- Chardon, F.; Bedu, M.; Calenge, F.; Klemens, P.A.W.; Spinner, L.; Clement, G.; Chietera, G.; Léran, S.; Ferrand, M.; Lacombe, B.; et al. Leaf fructose content is controlled by the vacuolar transporter SWEET17 in Arabidopsis. Curr. Biol. 2013, 23, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Shammai, A.; Petreikov, M.; Yeselson, Y.; Faigenboim, A.; Moy-Komemi, M.; Cohen, S.; Cohen, D.; Besaulov, E.; Efrati, A.; Houminer, N.; et al. Natural genetic variation for expression of a SWEET transporter among wild species of Solanum lycopersicum (tomato) determines the hexose composition of ripening tomato fruit. Plant J. 2018, 96, 343–357. [Google Scholar] [CrossRef]
- Zhang, Z.; Zou, L.M.; Ren, C.; Ren, F.; Wang, Y.; Fan, P.; Li, S.H.; Liang, Z.C. VvSWEET10 mediates sugar accumulation in grapes. Genes 2019, 10, 255. [Google Scholar] [CrossRef]
- Li, X.Y.; Guo, W.; Li, J.C.; Yue, P.T.; Bu, H.D.; Jiang, J.; Liu, W.T.; Xu, Y.X.; Yuan, H.; Li, T.; et al. Histone acetylation at the promoter for the transcription factor PuWRKY31 affects sucrose accumulation in pear fruit. Plant Physiol. 2020, 182, 2035–2046. [Google Scholar] [CrossRef]
- Ko, H.; Ho, L.; Neuhaus, H.E.; Guo, W. Transporter SlSWEET15 unloads sucrose from phloem and seed coat for fruit and seed development in tomato. Plant Physiol. 2021, 187, 2230–2245. [Google Scholar] [CrossRef]
- Wang, S.; Yokosho, K.; Guo, R.; Whelan, J.; Ruan, Y.-L.; Ma, J.F.; Shou, H. The soybean sugar transporter GmSWEET15 mediates sucrose export from endosperm to early embryo. Plant Physiol. 2019, 180, 2133–2141. [Google Scholar] [CrossRef]
- Lin, Q.H.; Zhong, Q.Z.; Zhang, Z.H. Identification and functional analysis of SWEET gene family in Averrhoa carambola L. fruits during ripening. Peer J. 2021, 9, e11404. [Google Scholar] [CrossRef]
- Zhen, Q.L.; Fang, T.; Peng, Q.; Liao, L.; Li Zhao, L.; Owiti, A.; Han, Y.P. Developing gene-tagged molecular markers for evaluation of genetic association of apple SWEET genes with fruit sugar accumulation. Hortic. Res. 2018, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Li, H.; Xia, X.; Liu, X.; Yang, L. Functional and evolution characterization of SWEET sugar transporters in Ananas Comosus. Biochem. Biophys. Res. Commun. 2018, 496, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Frommer, W.B. Transport of sugars. Annu. Rev. Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Ya Rao, Y.; Wang, M.Z.; Li, Y.; Liu, Y.J.; Xiong, P.P.; Zeng, L.H. Characterization of the SWEET gene family in longan (Dimocarpus longan) and the role of DlSWEET1 in cold tolerance. Int. J. Mol. Sci. 2022, 23, 8914. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Huh, J.; Yu, Y.; Ho, L.; Chen, L.; Tholl, D.; Fromme, W.B.; Guo, W. The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection. Plant J. 2015, 83, 1046–1058. [Google Scholar] [CrossRef]
- Guo, W.; Nagy, R.; Chen, H.; Frunder, S.; Yu, Y.; Santelia, D.; Frommer, W.; Martinoia, E. SWEET17, a facilitative transporter, mediates fructose transport across the tonoplast of Arabidopsis roots and leaves. Plant Physiol. 2014, 164, 777–789. [Google Scholar] [CrossRef]
- Abelenda, J.; Bergonzi, S.; Oortwijn, M.; Sonnewald, S.; Du, M.; Visser, R.; Sonnewald, U.; Bachem, C. Source-sink regulation is mediated by interaction of an FT Homolog with a SWEET protein in potato. Curr. Biol. 2019, 9, 1178–1186. [Google Scholar] [CrossRef]
- Wang, P.P.; Wei, P.N.; Niu, F.F.; Liu, X.F.; Zhang, H.L.; Lyu, M.L.; Yuan, Y.; Wu, B.H. Cloning and functional assessments of floral-expressed SWEET transporter genes from Jasminum sambac. Int. J. Mol. Sci. 2019, 20, 4001. [Google Scholar] [CrossRef]
- Andrés, F.; Kinoshita, A.; Kalluri, N.; Fernández, V.; Falavigna, V.; Cruz, T.; Jang, S.; Chiba, Y.; Mitsunori Seo, M.; Mettler-Altmann, T.; et al. The sugar transporter SWEET10 acts downstream of FLOWERING LOCUS T during floral transition of Arabidopsis thaliana. BMC Plant Biol. 2020, 20, 53. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, D.; Miao, Q.; Yang, J.; Xuan, Y.; Hu, Y. Essential role of sugar transporter OsSWEET11 during the early stage of rice grain filling. Plant Cell Physiol. 2017, 58, 863–873. [Google Scholar] [CrossRef]
- Engel, M.L.; Holmes-Davis, R.; McCormick, S. Green sperm. Identification of male gamete promoters in Arabidopsis. Plant Physiol. 2005, 138, 2124–2133. [Google Scholar] [CrossRef]
- Li, J.; Chen, D.; Jiang, G.; Song, H.; Tu, M.; Sun, S. Molecular cloning and expression analysis of EjSWEET15, encoding for a sugar transporter from loquat. Sci. Hortic. 2020, 272, 109552. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, C.; Han, X.; Wang, Z.Y.; Ma, L.; Yuan, D.P.; Wu, J.N.; Zhu, X.F.; Liu, J.M.; Li, D.P.; et al. Inhibition of OsSWEET11 function in mesophyll cells improves resistance of rice to sheath blight disease. Mol. Plant Pathol. 2018, 19, 2149–2161. [Google Scholar] [CrossRef] [PubMed]
- Bezrutczyk, M.; Hartwig, T.; Horschman, M.; Char, S.N.; Yang, J.; Yang, B.; Frommer, W.B.; Sosso, D. Impaired phloem loading in zmsweet13a,b,c sucrose transporter triple knock-out mutants in Zea mays. New Phytol. 2018, 218, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhu, Y.P.; Liu, M.; Zhou, Y.; Lu, G.Y.; Lan, L.; Wang, X.P.; Xuejun CZhang, X.J. Molecular mechanism of substrate recognition and transport by the AtSWEET13 sugar transporter. Proc. Natl. Acad. Sci. USA 2017, 114, 10089–10094. [Google Scholar] [CrossRef]
- Savoi, S.; Torregrosal, L.; Romieu, C. Transcripts switched off at the stop of phloem unloading highlight the energy efficiency of sugar import in the ripening V. vinifera fruit. Hortic. Res. 2021, 8, 193. [Google Scholar] [CrossRef]
- Lin, W.Q.; Xiao, X.O.; Zhang, H.N.; Li, Y.H.; Liu, S.H.; Sun, W.S.; Zhang, X.Z.; Wu, Q.S. Whole-genome bisulfite sequencing reveals a role for DNA methylation in variants from callus culture of pineapple (Ananas comosus L.). Genes 2019, 10, 877. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.; Frank, M.; He, Y.; Xia, R. TBtools—An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6, Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Lin, W.Q.; Xiao, X.O.; Sun, W.S.; Liu, S.H.; Wu, Q.S.; Yao, Y.L.; Zhang, H.N.; Zhang, X.Z. Genome-wide identification and expression analysis of cytosine DNA methyltransferase genes related to somaclonal variation in Pineapple (Ananas comosus L.). Agronomy 2022, 12, 1039. [Google Scholar] [CrossRef]
- Luan, A.P.; He, Y.H.; Xie, T.; Chen, C.J.; Mao, Q.; Wang, X.S.; Li, C.H.; Yaqi Ding, Y.Q.; Lin, W.Q.; Liu, C.Y.; et al. Identification of an embryonic cell-specific region within the pineapple SERK1 promoter. Genes 2019, 10, 883. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Uchii, S.; Watanabe, S.; Ezura, H. A highly efficient transformation protocol for Micro-Tom, a model cultivar for tomato functional genomics. Plant Cell Physiol. 2006, 47, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Nie, P.X.; Wang, X.Y.; Hu, L.P.; Zhang, H.Y.; Zhang, J.X.; Zhang, Z.X.; Zhang, L.Y. The predominance of the apoplasmic phloem unloading pathway is interrupted by a symplasmic pathway during Chinese jujube fruit development. Plant Cell Physiol. 2010, 51, 1007–1018. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).