
Metabolomics Analysis Revealed Significant Metabolic Changes in Brain Cancer Cells Treated with Paclitaxel and/or Etoposide
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
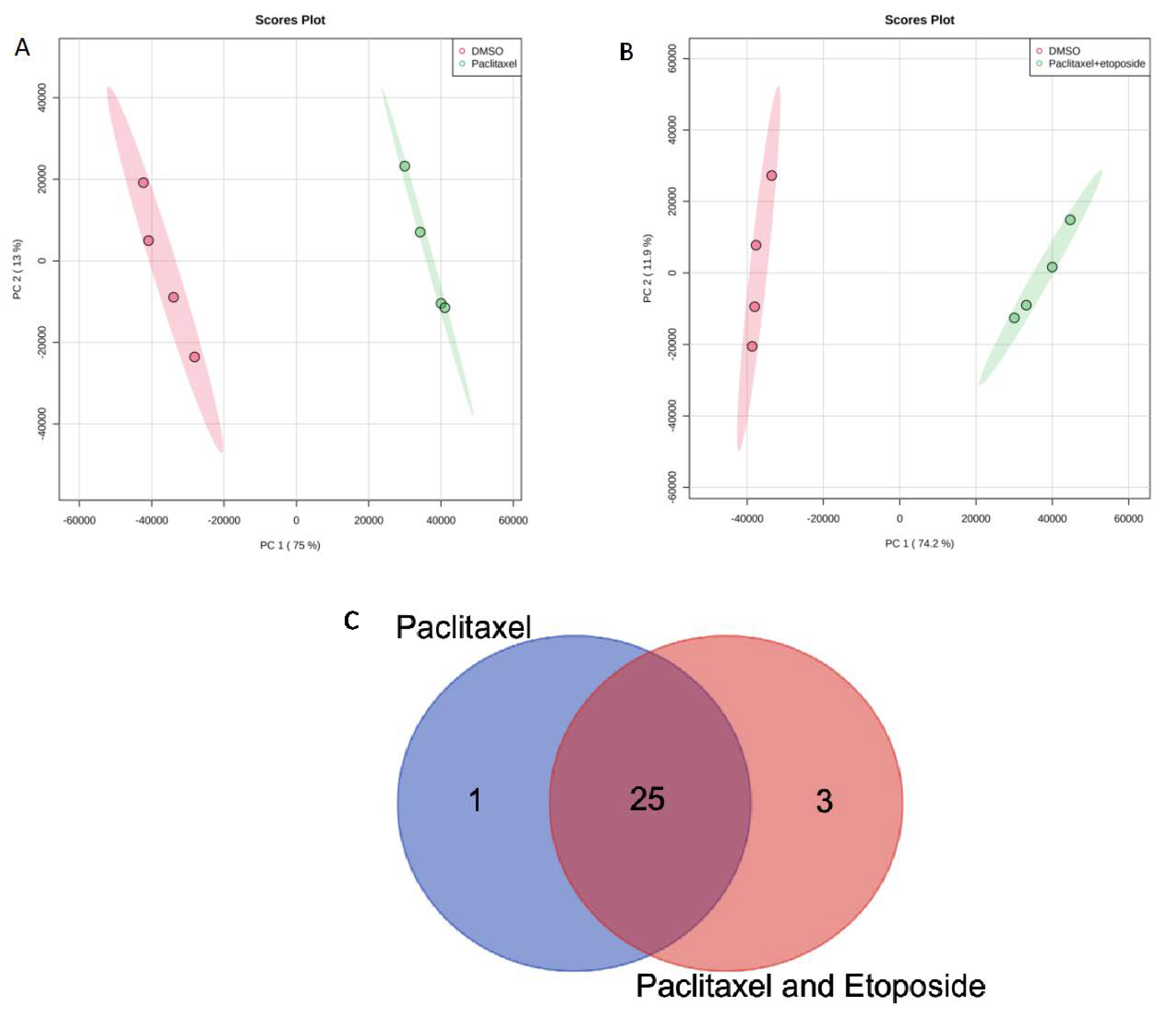
2.1. Metabolites in U87 Cell Line
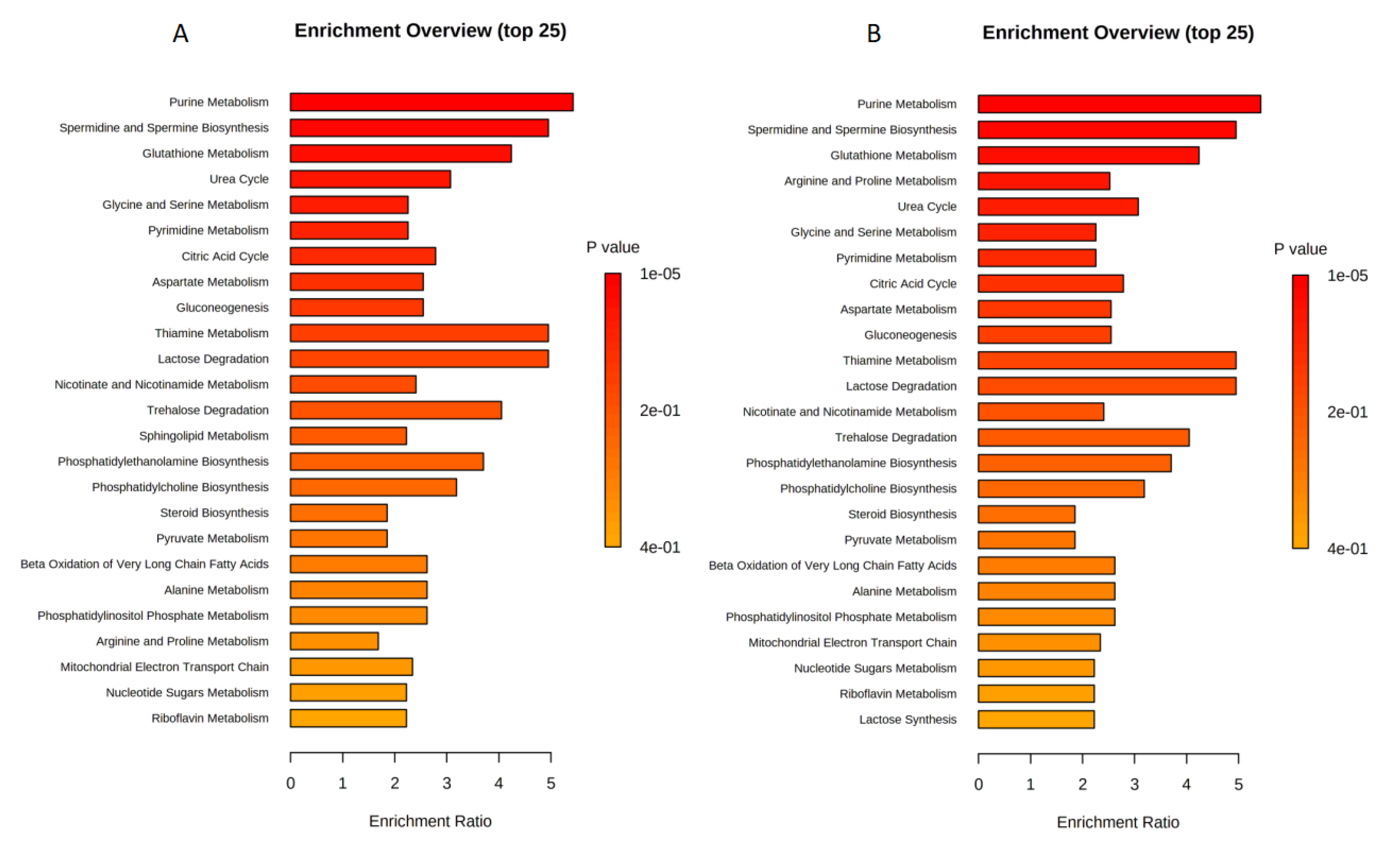
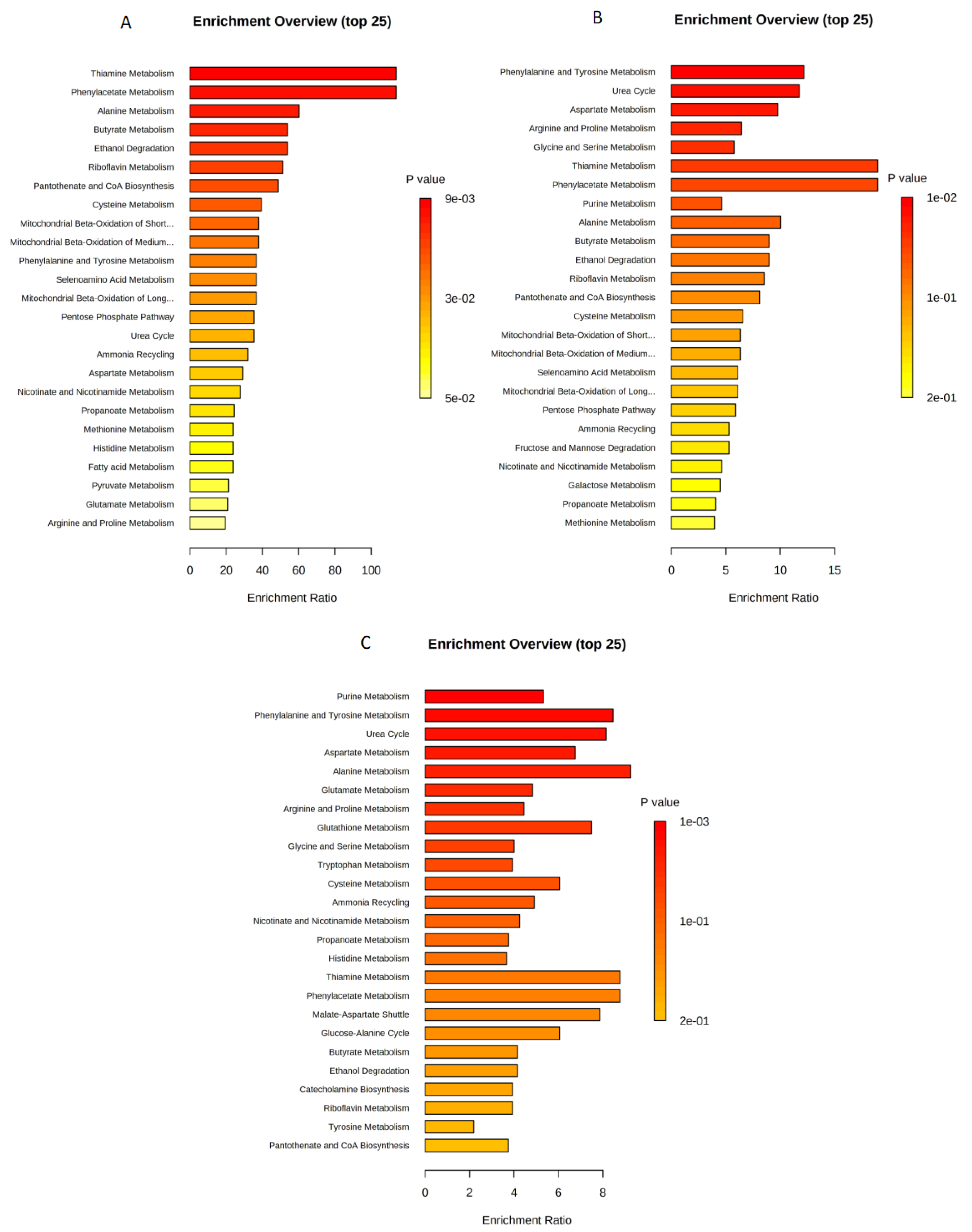
2.2. Enrichment Analysis for U87 Cell Line
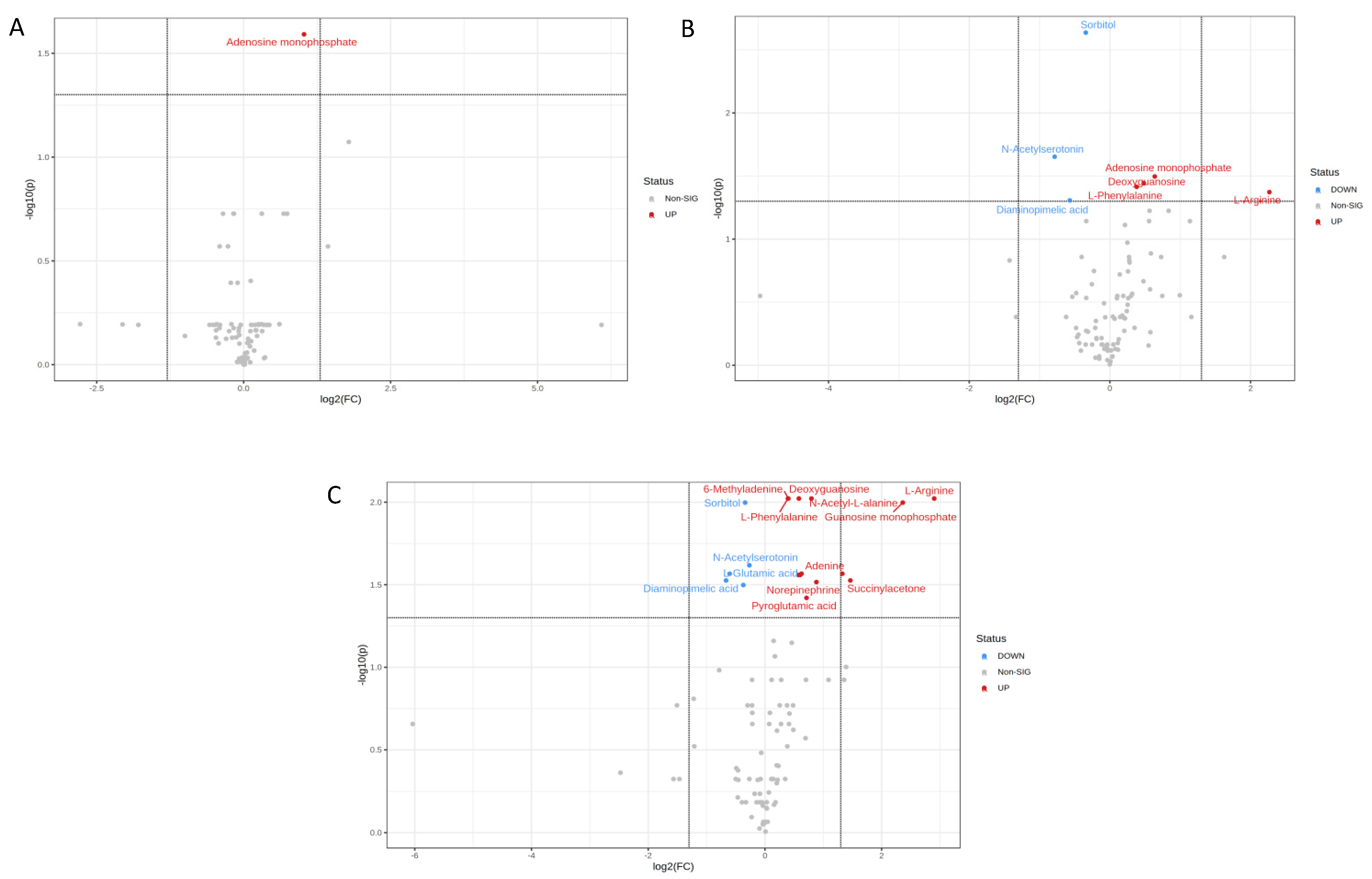
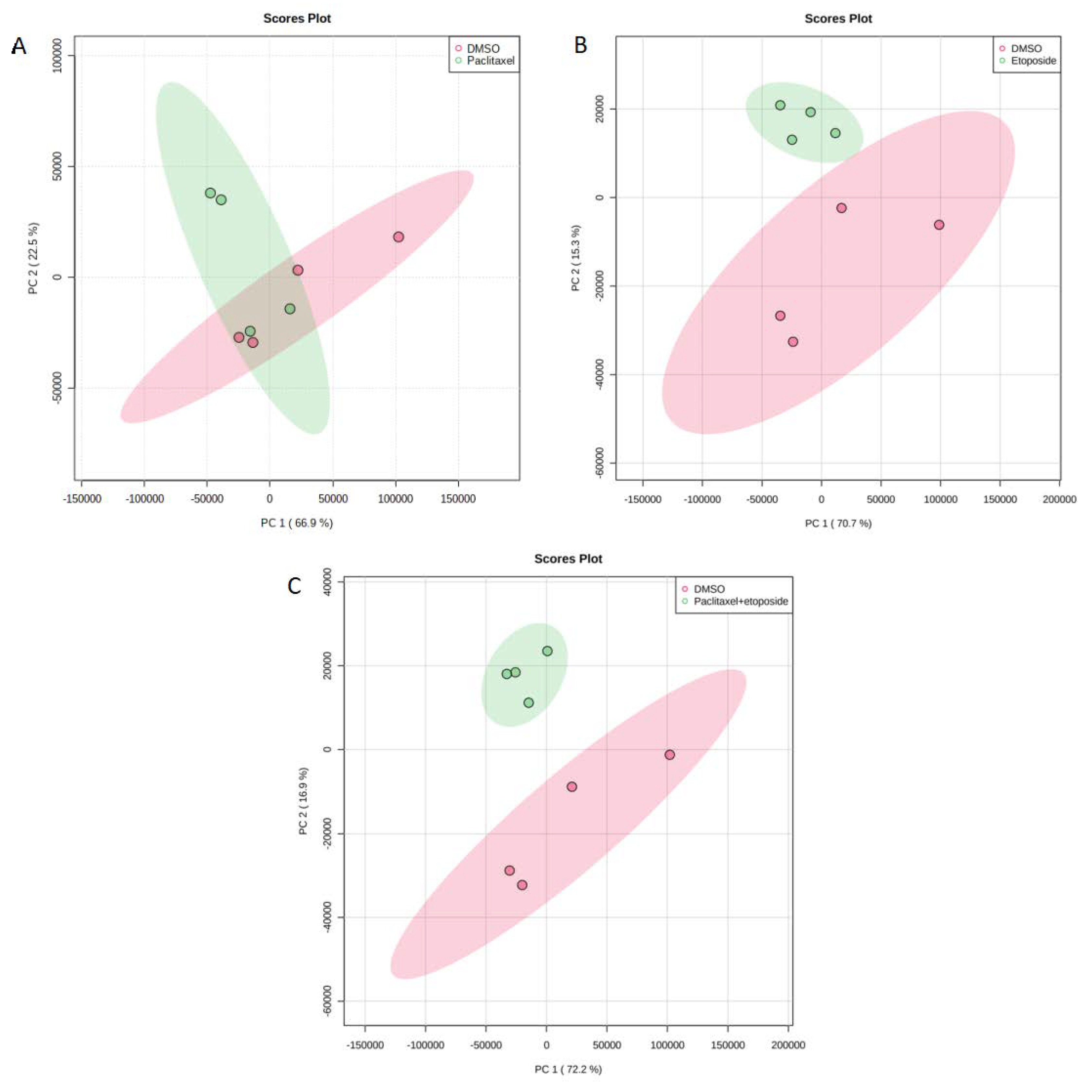
2.3. Metabolites in U373 Cell Line
2.4. Enrichment Analysis for U373 Cell Line
3. Discussion
4. Materials and Methods
4.1. Reagents and Materials
4.2. Cell Culture
4.3. Treatment of Cells with Anticancer Drugs
4.4. Cells Samples Preparation and Metabolites Extraction
4.5. Tandem Liquid Chromatography Mass Spectrometry (QTOF MS) Conditions
4.6. Data Processing and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Worldwide Cancer Data|World Cancer Research Fund International. Available online: https://www.wcrf.org/cancer-trends/worldwide-cancer-data/ (accessed on 7 July 2022).
- Mesfin, F.B.; Al-Dhahir, M.A. Gliomas; StatPearls: Tampa, FL, USA, 2022. [Google Scholar]
- Alexander, S.; Slazinski, T.; Zrelak, P.; Lovely, M.P.; Stewart-Amidei, C.; Arzbaecher, J.; Bell, S.; Maher, M.E.; Maida, M.; Mogensen, K.; et al. Care of the Adult Patient with a Brain Tumor. 2014. Available online: https://aann.org/uploads/Publications/CPGs/AANN14_ABT_Module_FINAL.pdf (accessed on 5 September 2022).
- Ostrom, Q.T.; Gittleman, H.; Fulop, J.; Liu, M.; Blanda, R.; Kromer, C.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2008–2012. Neuro Oncol. 2015, 17, iv1–iv62. [Google Scholar] [CrossRef] [PubMed]
- Glioblastoma: A Q&A about Brain Cancer|Cancer Care|Healthy News Blog|Lee Health. Available online: https://www.leehealth.org/health-and-wellness/healthy-news-blog/cancer-care/glioblastoma-a-q-a-about-brain-cancer (accessed on 8 September 2022).
- Davis, M.E. Glioblastoma: Overview of Disease and Treatment. Clin. J. Oncol. Nurs. 2016, 20, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, T.A.; Karajannis, M.A.; Harter, D.H. Glioblastoma Multiforme: State of the Art and Future Therapeutics. Surg. Neurol. Int. 2014, 5, 64. [Google Scholar] [CrossRef]
- TimsTOF|Bruker. Available online: https://www.bruker.com/en/products-and-solutions/mass-spectrometry/timstof.html (accessed on 7 July 2022).
- Chakraborty, S.; Hosen, M.I.; Ahmed, M.; Shekhar, H.U. Onco-Multi-OMICS Approach: A New Frontier in Cancer Research. Biomed. Res. Int. 2018, 2018, 9836256. [Google Scholar] [CrossRef] [Green Version]
- St-Coeur, P.D.; Poitras, J.J.; Cuperlovic-Culf, M.; Touaibia, M.; Morin, P.J. Investigating a Signature of Temozolomide Resistance in GBM Cell Lines Using Metabolomics. J. Neuro-Oncol. 2015, 125, 91–102. [Google Scholar] [CrossRef]
- Paclitaxel Is a Suitable Chemotherapy for Patients with Metastatic Brain Tumors. Nat. Clin. Pract. Neurol. 2007, 3, 69. [CrossRef]
- Carrillo, J.A.; Hsu, F.P.K.; Delashaw, J.; Bota, D. Efficacy and Safety of Bevacizumab and Etoposide Combination in Patients with Recurrent Malignant Gliomas Who Have Failed Bevacizumab. Rev. Health Care 2014, 5, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond Biomarkers and towards Mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451. [Google Scholar] [CrossRef] [Green Version]
- Human Metabolome Database: Showing Metabocard for Nutriacholic Acid (HMDB0000467). Available online: https://hmdb.ca/metabolites/HMDB0000467 (accessed on 5 September 2022).
- Lu, D.; Yang, F.; Lin, Z.; Zhuo, J.; Liu, P.; Cen, B.; Lian, Z.; Xie, H.; Zheng, S.; Xu, X. A Prognostic Fingerprint in Liver Transplantation for Hepatocellular Carcinoma Based on Plasma Metabolomics Profiling. Eur. J. Surg. Oncol. 2019, 45, 2347–2352. [Google Scholar] [CrossRef]
- Tamanoi, F.; Azizian, M.; Ashrafi, M.; Bathaie, S. Mevalonate Pathway and Human Cancers. Curr. Mol. Pharmacol. 2017, 10, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.S.; Noh, K.; Haemmerle, M.; Li, D.; Park, H.; Hu, Q.; Hisamatsu, T.; Mitamura, T.; Mak, S.L.C.; Kunapuli, S.; et al. Role of ADP Receptors on Platelets in the Growth of Ovarian Cancer. Blood 2017, 130, 1235. [Google Scholar] [CrossRef] [PubMed]
- Belluardo, N.; Mudò, G.; di Liberto, V.; Frinchi, M.; Condorelli, D.F.; Traversa, U.; Ciruela, F.; Ciccarelli, R.; di Iorio, P.; Giuliani, P. Investigating the Role of Guanosine on Human Neuroblastoma Cell Differentiation and the Underlying Molecular Mechanisms. Front. Pharmacol. 2021, 12, 729. [Google Scholar] [CrossRef] [PubMed]
- Human Metabolome Database: Showing Metabocard for N-Acetyl-L-Alanine (HMDB0000766). Available online: https://hmdb.ca/metabolites/HMDB0000766 (accessed on 7 September 2022).
- Zand, B.; Previs, R.A.; Zacharias, N.M.; Rupaimoole, R.; Mitamura, T.; Nagaraja, A.S.; Guindani, M.; Dalton, H.J.; Yang, L.; Baddour, J.; et al. Role of Increased N-Acetylaspartate Levels in Cancer. J. Natl. Cancer Inst. 2016, 108, djv426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.L.; Hsu, S.C.; Ann, D.K.; Yen, Y.; Kung, H.J. Arginine Signaling and Cancer Metabolism. Cancers 2021, 13, 3541. [Google Scholar] [CrossRef]
- Yin, J.; Ren, W.; Huang, X.; Deng, J.; Li, T.; Yin, Y. Potential Mechanisms Connecting Purine Metabolism and Cancer Therapy. Front. Immunol. 2018, 9, 1697. [Google Scholar] [CrossRef] [Green Version]
- Su, W.J.; Lu, P.Z.; Wu, Y.; Kalpana, K.; Yang, C.K.; Lu, G.D. Identification of Key Genes in Purine Metabolism as Prognostic Biomarker for Hepatocellular Carcinoma. Front. Oncol. 2021, 10, 3057. [Google Scholar] [CrossRef]
- Siddiqui, A.; Ceppi, P. A Non-Proliferative Role of Pyrimidine Metabolism in Cancer. Mol. Metab. 2020, 35, 100962. [Google Scholar] [CrossRef]
- Lee, J.S.; Adler, L.; Karathia, H.; Hannenhalli, S.; Ruppin, E.; Erez, A. Urea Cycle Dysregulation Generates Clinically Relevant Genomic and Biochemical Signatures. Biomol. Ther. 2018, 26, 19. [Google Scholar] [CrossRef] [Green Version]
- Keshet, R.; Szlosarek, P.; Carracedo, A.; Erez, A. Rewiring Urea Cycle Metabolism in Cancer to Support Anabolism. Nat. Rev. Cancer 2018, 18, 634–645. [Google Scholar] [CrossRef]
- Traverso, N.; Ricciarelli, R.; Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C. Role of Glutathione in Cancer Progression and Chemoresistance. Oxid. Med. Cell Longev. 2013, 2013, 972913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant Properties Dedicated to Nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, L.; Sandhu, J.K.; Harper, M.E.; Cuperlovic-culf, M. Role of Glutathione in Cancer: From Mechanisms to Therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Storz, P. Reactive Oxygen Species in Cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Backos, D.S.; Franklin, C.C.; Reigan, P. The Role of Glutathione in Brain Tumor Drug Resistance. Biochem. Pharmacol. 2012, 83, 1005–1012. [Google Scholar] [CrossRef]
- Anderson, N.M.; Mucka, P.; Kern, J.G.; Feng, H. The Emerging Role and Targetability of the TCA Cycle in Cancer Metabolism. Protein Cell 2018, 9, 216. [Google Scholar] [CrossRef]
- Akram, M. Citric Acid Cycle and Role of Its Intermediates in Metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef]
- Liu, S.; Miriyala, S.; Keaton, M.A.; Jordan, C.T.; Wiedl, C.; Clair, D.K.S.; Moscow, J.A. Metabolic Effects of Acute Thiamine Depletion Are Reversed by Rapamycin in Breast and Leukemia Cells. PLoS ONE 2014, 9, e85702. [Google Scholar] [CrossRef]
- Lu’o’ng, K.V.Q.; Nguyên, L.T.H. Thiamine and Parkinson’s Disease. J. Neurol. Sci. 2012, 316, 1–8. [Google Scholar] [CrossRef]
- Comín-Anduix, B.; Boren, J.; Martinez, S.; Moro, C.; Centelles, J.J.; Trebukhina, R.; Petushok, N.; Lee, W.N.P.; Boros, L.G.; Cascante, M. The Effect of Thiamine Supplementation on Tumour Proliferation. Eur. J. Biochem. 2001, 268, 4177–4182. [Google Scholar] [CrossRef]
- Human Metabolome Database: Showing Metabocard for Phenyl Acetate (HMDB0040733). Available online: https://hmdb.ca/metabolites/HMDB0040733 (accessed on 10 September 2022).
- Phenylacetate Inhibits Growth and Modulates Cell Cycle Gene Expression in Renal Cancer Cell Lines—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/12820434/#:~:text=Background%3A%20Phenylacetate%20(PA)%2C,inhibition%20in%20prostate%20cancer%20cells (accessed on 10 September 2022).
- Scharlau, D.; Borowicki, A.; Habermann, N.; Hofmann, T.; Klenow, S.; Miene, C.; Munjal, U.; Stein, K.; Glei, M. Mechanisms of Primary Cancer Prevention by Butyrate and Other Products Formed during Gut Flora-Mediated Fermentation of Dietary Fibre. Mutat. Res. 2009, 682, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.W.; Yin, F.Y.; Zhang, Z.F.; Gong, X.; Yang, Y. Butyrate Suppresses Glucose Metabolism of Colorectal Cancer Cells via GPR109a-AKT Signaling Pathway and Enhances Chemotherapy. Front. Mol. Biosci. 2021, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Usiello, A.; di Fiore, M.M.; de Rosa, A.; Falvo, S.; Errico, F.; Santillo, A.; Nuzzo, T.; Baccari, G.C. New Evidence on the Role of D-Aspartate Metabolism in Regulating Brain and Endocrine System Physiology: From Preclinical Observations to Clinical Applications. Int. J. Mol. Sci. 2020, 21, 8718. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bermudez, J.; Baudrier, L.; La, K.; Zhu, X.G.; Fidelin, J.; Sviderskiy, V.O.; Papagiannakopoulos, T.; Molina, H.; Snuderl, M.; Lewis, C.A.; et al. Aspartate Is a Limiting Metabolite for Cancer Cell Proliferation under Hypoxia and in Tumors. Nat. Cell Biol. 2018, 20, 775. [Google Scholar] [CrossRef]
- Patri, M. Synaptic Transmission and Amino Acid Neurotransmitters. In Neurochemical Basis of Brain Function and Dysfunction; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino Acids in Cancer. Exp. Mol. Med. 2020, 52, 15. [Google Scholar] [CrossRef]
- Vettore, L.; Westbrook, R.L.; Tennant, D.A. New Aspects of Amino Acid Metabolism in Cancer. Br. J. Cancer 2020, 122, 150. [Google Scholar] [CrossRef]
- MetaboScape|Bruker. Available online: https://www.bruker.com/en/products-and-solutions/mass-spectrometry/ms-software/metaboscape.html (accessed on 19 May 2022).
- Human Metabolome Database. Available online: https://hmdb.ca/ (accessed on 20 April 2022).
- MetaboAnalyst. Available online: https://www.metaboanalyst.ca/ (accessed on 20 April 2022).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Statistically Significant Metabolites in U87 Cell Line Treated with Paclitaxel 4.2 nM/mL | ||||
|---|---|---|---|---|
| Metabolite | t-stat | p-value | FDR | Fold change |
| ADP | 39.479 | 0.000025 | 0.00045982 | 0.0073262 |
| Guanosine | 19.744 | 0.00017013 | 0.0017391 | 0.01855 |
| Guanine | 27.592 | 0.000075 | 0.0011507 | 0.018726 |
| Hypoxanthine | 29.998 | 0.00000482 | 0.00017402 | 0.028151 |
| L-Methionine | 11.449 | 0.0010822 | 0.0066372 | 0.031249 |
| p-Hydroxyphenylacetic acid | 12.337 | 0.00090239 | 0.00593 | 0.031378 |
| Glycyl-L-leucine | 11.288 | 0.00087297 | 0.00593 | 0.046307 |
| Adenine | 15.341 | 0.0000249 | 0.00045982 | 0.066085 |
| Glycerophosphocholine | 28.954 | 0.000000113 | 0.0000104 | 0.067262 |
| Pyroglutamic acid | 6.3228 | 0.0066699 | 0.027892 | 0.075638 |
| Inosine | 13.845 | 0.0005007 | 0.00458 | 0.081824 |
| Niacinamide | 19.588 | 0.00000567 | 0.00017402 | 0.084552 |
| L-Phenylalanine | 14.813 | 0.00063069 | 0.0048353 | 0.08486 |
| Spermine | 7.2734 | 0.0022883 | 0.01108 | 0.11408 |
| Uric acid | 22.543 | 0.00014664 | 0.0017391 | 0.11878 |
| Thymidine | 6.5862 | 0.0071226 | 0.02849 | 0.13229 |
| L-Arginine | 13.13 | 0.00015563 | 0.0017391 | 0.1326 |
| Cytidine | 7.7175 | 0.0045226 | 0.019813 | 0.14374 |
| L-Tryptophan | 9.8826 | 0.0021262 | 0.010867 | 0.1597 |
| Guanosine 5’-diphosphate | 11.515 | 0.0014062 | 0.0080855 | 0.16321 |
| Cyclic AMP | 8.3747 | 0.00054761 | 0.00458 | 0.23873 |
| Androstenedione | 4.5213 | 0.010954 | 0.038761 | 0.66828 |
| Sphinganine | 4.9041 | 0.0088823 | 0.034049 | 0.68976 |
| L-Norleucine | 6.6297 | 0.0030279 | 0.013928 | 0.71434 |
| Mevalonic acid | −3.8293 | 0.010428 | 0.038375 | 1.1212 |
| Nutriacholic acid | −10.591 | 0.0017986 | 0.0097337 | 6.25 |
| B. Statistically significant metabolites in U87 cell line treated with paclitaxel 4.2 nM /mL + etoposide 10 μg/mL | ||||
| Metabolite | t-stat | p-value | FDR | Fold change |
| ADP | 39.641 | 0.0000273 | 0.00041814 | 0.0062162 |
| Guanosine | 20.192 | 0.00022842 | 0.0026268 | 0.010662 |
| Guanine | 27.875 | 0.0000928 | 0.0012194 | 0.016895 |
| Thymidine | 7.3937 | 0.0048716 | 0.021342 | 0.022396 |
| Hypoxanthine | 28.231 | 0.00000129 | 0.0000592 | 0.029428 |
| Glycerophosphocholine | 35.644 | 0.000000565 | 0.000052 | 0.061413 |
| Inosine | 13.625 | 0.00028557 | 0.0029192 | 0.069193 |
| Glycyl-L-leucine | 11.044 | 0.0009866 | 0.0069563 | 0.069508 |
| Uric acid | 21.477 | 0.0000139 | 0.00036381 | 0.075097 |
| Niacinamide | 20.815 | 0.000019 | 0.00036381 | 0.091824 |
| L-Phenylalanine | 14.45 | 0.00060549 | 0.0055705 | 0.10242 |
| Pyroglutamic acid | 6.1554 | 0.0081168 | 0.032467 | 0.10904 |
| Cyclic AMP | 10.615 | 0.0010586 | 0.0069563 | 0.13018 |
| Adenine | 13.059 | 0.0000198 | 0.00036381 | 0.13747 |
| Cytidine | 7.7175 | 0.0045226 | 0.020804 | 0.14374 |
| L-Methionine | 10.229 | 0.0018547 | 0.0089805 | 0.14382 |
| p-Hydroxyphenylacetic acid | 10.849 | 0.0016757 | 0.0089805 | 0.15864 |
| Guanosine 5’-diphosphate | 11.515 | 0.0014062 | 0.0086245 | 0.16321 |
| L-Tryptophan | 9.6991 | 0.0017879 | 0.0089805 | 0.16393 |
| Spermine | 5.7422 | 0.0016656 | 0.0089805 | 0.1902 |
| L-Arginine | 7.0817 | 0.00070717 | 0.0057029 | 0.23436 |
| L-Norleucine | 3.6225 | 0.0113 | 0.043318 | 0.80446 |
| 3,4-Dihydroxybenzeneacetic acid | 3.4628 | 0.013422 | 0.047493 | 0.93086 |
| Phosphocreatine | −3.5105 | 0.012692 | 0.046708 | 1.0684 |
| Mevalonic acid | −6.3456 | 0.00074386 | 0.0057029 | 1.1743 |
| Nutriacholic acid | −6.5371 | 0.0072758 | 0.030426 | 6.7728 |
| Groups | Metabolites |
|---|---|
| Common metabolites between paclitaxel and paclitaxel + etoposide treatments (25) | Niacinamide Nutriacholic acid Glycyl-L-leucine L-Arginine L-Norleucine Guanosine 5’-diphosphate Guanine Cytidine Thymidine Sphinganine Spermine p-Hydroxyphenylacetic acid L-Tryptophan Cyclic AMP Mevalonic acid Glycerophosphocholine ADP Hypoxanthine L-Methionine Pyroglutamic acid Adenine Guanosine L-Phenylalanine Uric acid Inosine |
| Unique metabolite from paclitaxel treatment (1) | Androstenedione |
| Unique metabolites from paclitaxel and etoposide treatment (3) | 3,4-Dihydroxybenzeneacetic acid Saccharopine Phosphocreatine |
| A. Statistically Significant Metabolites in U373 Cells Treated with Etoposide 10 μg/mL | ||||
|---|---|---|---|---|
| t-stat | p-value | FDR | Fold Change | |
| N-Acetylserotonin | 9.5789 | 0.00047303 | 0.022232 | 0.58014 |
| Diaminopimelic acid | 4.6918 | 0.003674 | 0.049336 | 0.67382 |
| Sorbitol | 11.637 | 0.0000246 | 0.002309 | 0.78847 |
| L-Phenylalanine | −5.9766 | 0.0020496 | 0.038532 | 1.3019 |
| Deoxyguanosine | −5.4895 | 0.0015345 | 0.036061 | 1.397 |
| Adenosine monophosphate | −6.3756 | 0.0010177 | 0.031888 | 1.5557 |
| L-Arginine | −5.0157 | 0.0027084 | 0.042432 | 4.8143 |
| B. Statistically significant metabolites in U373 cells treated with paclitaxel nM /mL + etoposide 10 μg/mL | ||||
| t-stat | p-value | FDR | Fold Change | |
| Diaminopimelic acid | 4.4706 | 0.0045103 | 0.029796 | 0.62996 |
| L-Glutamic acid | 5.0364 | 0.0031745 | 0.027128 | 0.65732 |
| 3a,6b,7b-Trihydroxy-5b-cholanoic acid | 5.0002 | 0.0057423 | 0.031751 | 0.77372 |
| Sorbitol | 7.9213 | 0.00064331 | 0.01006 | 0.79015 |
| N-Acetylserotonin | 8.7249 | 0.0020526 | 0.024118 | 0.83152 |
| L-Phenylalanine | −10.2 | 0.00012294 | 0.0095052 | 1.3149 |
| 6-Methyladenine | −7.3836 | 0.0005056 | 0.0095052 | 1.32 |
| L-Tryptophan | −4.6148 | 0.0038147 | 0.027583 | 1.4871 |
| Deoxyguanosine | −6.8588 | 0.00047465 | 0.0095052 | 1.4964 |
| Glycerophosphocholine | -4.9201 | 0.0036767 | 0.027583 | 1.5182 |
| Adenine | -4.8976 | 0.0028844 | 0.027113 | 1.5443 |
| Pyroglutamic acid | −4.333 | 0.0072763 | 0.037998 | 1.6388 |
| N-Acetyl-L-alanine | −8.0485 | 0.00027672 | 0.0095052 | 1.7372 |
| Norepinephrine | −5.415 | 0.0051875 | 0.030476 | 1.8433 |
| Adenosine monophosphate | −7.0313 | 0.0028079 | 0.027113 | 2.5075 |
| Succinylacetone | −7.2032 | 0.0047548 | 0.029796 | 2.7582 |
| Guanosine monophosphate | −6.8338 | 0.00074918 | 0.01006 | 5.1413 |
| L-Arginine | −7.9259 | 0.000333 | 0.0095052 | 7.4674 |
| Groups | Metabolites |
|---|---|
| Common metabolites between paclitaxel and paclitaxel + etoposide treatments (1) | Adenosine monophosphate |
| Unique metabolites from paclitaxel and etoposide treatment (6) | N-Acetylserotonin L-Arginine Diaminopimelic acid Sorbitol Deoxyguanosine L-Phenylalanine |
| Unique metabolites from paclitaxel and etoposide treatment (11) | 6-Methyladenine Succinylacetone Guanosine monophosphate L-Glutamic acid L-Tryptophan N-Acetyl-L-alanine Glycerophosphocholine Pyroglutamic acid Adenine Norepinephrine 3a,6b,7b-Trihydroxy-5b-cholanoic acid |
| Time Point (min) | Mobile Phase Concentration (A = Water with 0.1% Formic Acid; B = Acetonitrile with 0.1% Formic Acid) | Flow Rate |
|---|---|---|
| 0–2 | 99% A and 1% B | 0.25 mL/min |
| 2–17 | 99 to 1% A and 1 to 99% B | 0.25 mL/min |
| 17–20 | 99% B and 1% A | 0.25 mL/min |
| 20–20.1 | 99% B changed to 99% A | 0.35 mL/min |
| 20.1–28.5 | 99% A changed to 1% B | 0.35 mL/min |
| 28.5–30 | 99% A changed to 1% B | 0.25 mL/min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semreen, A.M.; Alsoud, L.O.; El-Huneidi, W.; Ahmed, M.; Bustanji, Y.; Abu-Gharbieh, E.; El-Awady, R.; Ramadan, W.S.; Alqudah, M.A.Y.; Shara, M.; et al. Metabolomics Analysis Revealed Significant Metabolic Changes in Brain Cancer Cells Treated with Paclitaxel and/or Etoposide. Int. J. Mol. Sci. 2022, 23, 13940. https://doi.org/10.3390/ijms232213940
Semreen AM, Alsoud LO, El-Huneidi W, Ahmed M, Bustanji Y, Abu-Gharbieh E, El-Awady R, Ramadan WS, Alqudah MAY, Shara M, et al. Metabolomics Analysis Revealed Significant Metabolic Changes in Brain Cancer Cells Treated with Paclitaxel and/or Etoposide. International Journal of Molecular Sciences. 2022; 23(22):13940. https://doi.org/10.3390/ijms232213940
Chicago/Turabian StyleSemreen, Ahlam M., Leen Oyoun Alsoud, Waseem El-Huneidi, Munazza Ahmed, Yasser Bustanji, Eman Abu-Gharbieh, Raafat El-Awady, Wafaa S. Ramadan, Mohammad A.Y. Alqudah, Mohd Shara, and et al. 2022. "Metabolomics Analysis Revealed Significant Metabolic Changes in Brain Cancer Cells Treated with Paclitaxel and/or Etoposide" International Journal of Molecular Sciences 23, no. 22: 13940. https://doi.org/10.3390/ijms232213940