Abstract
Cytochrome P450 2U1 (CYP2U1) identified from the human genome remains poorly known since few data are presently available on its physiological function(s) and substrate(s) specificity. CYP2U1 mutations are associated with complicated forms of hereditary spastic paraplegia, alterations of mitochondrial architecture and bioenergetics. In order to better know the biological roles of CYP2U1, we used a bioinformatics approach. The analysis of the data invited us to focus on leukotriene B4 (LTB4), an important inflammatory mediator. Here, we show that CYP2U1 efficiently catalyzes the hydroxylation of LTB4 predominantly on its ω-position. We also report docking experiments of LTB4 in a 3D model of truncated CYP2U1 that are in agreement with this hydroxylation regioselectivity. The involvement of CYP2U1 in the metabolism of LTB4 could have strong physiological consequences in cerebral pathologies including ischemic stroke because CYP2U1 is predominantly expressed in the brain.
1. Introduction
Cytochromes P450 (CYPs) are members of a superfamily of heme-thiolate proteins that are widely distributed in living organisms and are involved in the monooxygenation of endogenous compounds, such as steroids, fatty acids, eicosanoids and vitamins, and xenobiotics such as drugs and environmental compounds [1]. Among the 57 human CYPs, CYP2U1 is intriguing because of its singularity in terms of gene organization, protein sequence, and tissue expression (mainly expressed in brain and thymus) [2,3,4,5,6]. CYP2U1 is the only member known so far of this subfamily; it seems to be very old and highly conserved across species [2,3,4,7]. However, presently, the biological functions of CYP2U1 in humans are poorly known. It was found to hydroxylate arachidonic acid (AA) on its ω- and ω-1 positions [8] and to oxidize N-arachidonoylserotonin on position 2 of the indole ring [9] with Km values at the high µM level but with low kcat values (around 0.02 min−1). It was also found to hydroxylate xenobiotics such as debrisoquine and terfenadone analogues but with Km values at the mM level [10].
CYP2U1 mutations have been associated with complicated forms of hereditary spastic paraplegia that are characterized by variable conditions of neurologic and extra-neurologic disorders. Alterations of mitochondrial architecture and bioenergetics were associated with these pathophysiologies, suggesting a key role of CYP2U1 in lipids and energetic metabolism [11,12,13]. In order to better know the biological roles of CYP2U1 and to find possible new endogenous substrates, we used a bioinformatics approach carried out through molecular modelling studies using a previously described 3D-model of CYP2U1 [10]. The analysis of the data invited us to focus on leukotriene B4, an important mediator of inflammation derived from AA by the actions of 5-lipoxygenase and leukotriene A4 hydrolase [14,15]. LTB4 is a potent chemotactic agent able to recruit and activate neutrophiles and thus exerts a wide range of biological effects including brain inflammatory disorders [16,17,18,19,20]. Moreover, CYP2U1 is mainly expressed in the brain [2,3]. This led us to study the possible implication of CYP2U1 in the metabolism of LTB4. In the present manuscript, we show that CYP2U1 efficiently catalyzes the hydroxylation of LTB4 predominantly on its ω- position (Scheme 1) and report docking experiments of LTB4 in a 3D model structure of CYP2U1 [10] that are in agreement with this hydroxylation regioselectivity.
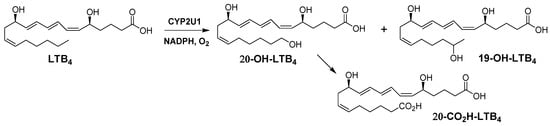
Scheme 1.
Structures of LTB4 and of metabolites obtained from incubations of LTB4 in the presence of NADPH and yeast microsomes expressing human CYP2U1.
2. Results
2.1. Screening of Databases
Our research strategy for finding new substrates for CYP2U1 was based on a bioinformatics approach carried out through molecular modelling studies using a virtual screening of HMDB, Cayman and Lipid Maps-Lipidomics databases on our 3D homology model of truncated CYP2U1 [10]. Further details on the choice of molecules are indicated in Materials and Methods (Section 4.4), and complete data of this study will be published elsewhere. Among the best promising compounds, we selected LTB4 as a putative substrate for CYP2U1 because of its important biological properties.
2.2. LTB4 Metabolism by Yeast Microsomes Expressing CYP2U1
Figure 1A shows the HPLC analysis of a typical incubation of microsomes from yeast expressing CYP2U1 and human CPR in the presence of 20 µM LTB4 and 1 mM NADPH for 15 min at 28 °C. Metabolites were separated by HPLC and identified by their retention times, UV-vis spectra and mass-spectra by comparison with authentic compounds, as indicated in Materials and Methods. Co-injection with authentic 20-OH-LTB4, 19-OH-LTB4 and 20-CO2H-LTB4 (retention times 17.6, 17.2 and 17.4 min, respectively), and comparison of their UV-spectra (all with maximal absorbance at 270 nm) and mass-spectra (m/z 351 and 365, respectively) of their carboxylate anions fully identified these metabolites (Scheme 1, Figure 1). No further metabolite with either the typical UV-spectra with maximum absorbance at 270 nm, or mass-spectra with signals at m/z 337 (reduction of a double bond) or 349 (formation of an aldehyde) could be identified. Recombinant CYP2U1 predominantly formed 20-OH-LTB4 (~90%) with minor amounts of 19-OH-LTB4 and 20-CO2H-LTB4 (~6 and ~4%, respectively) (Figure 1A). Microsomes from yeast transformed with the empty vector pYeDP60 were inactive (data not shown). Omission of NADPH (Figure 1B) or the addition of 1 mM Bz-ImH, an inhibitor of CYPs [21], in the reaction mixture did not lead to any formation of those oxidation products (data not shown).
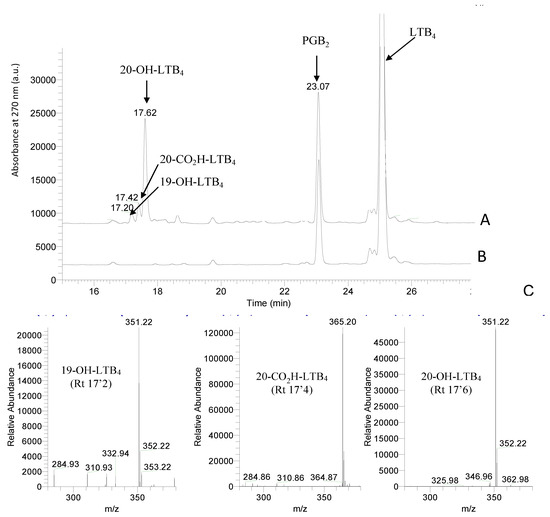
Figure 1.
(A) Typical reverse-phase HPLC analysis of LTB4 incubated for 15 min at 28 °C in the presence of microsomes from yeast transformed to express CYP2U1(incubation and analysis conditions in Materials and Methods). (B) Same incubation as in A but in the absence of NADPH. Figure shows the absorbance at 270 nm. (C) Mass spectra of the observed metabolites of LTB4 absorbing at 270 nm.
2.3. Kinetic Analysis of LTB4 Metabolism by CYP2U1
In the presence of 20 µM LTB4, the enzymatic activity, expressed as the sum of the three metabolites (19-OH-LTB4, 20-OH-LTB4 and 20-CO2H-LTB4) formed in 15 min at 28 °C, was dependent upon the microsomal proteins concentration and linearly varied in the 0 to 2 mg/mL range (data not shown). Kinetic experiments led to Km and Vm values of 12 ± 4 µM and 148 ± 25 pmol (total products)/min/mg protein, respectively (Figure 2). The affinity of LTB4 for CYP2U1 thus seems lower than that of AA (2.7 μM) [8] but higher than that observed for arachidonoylserotonin (82 µM) [9].
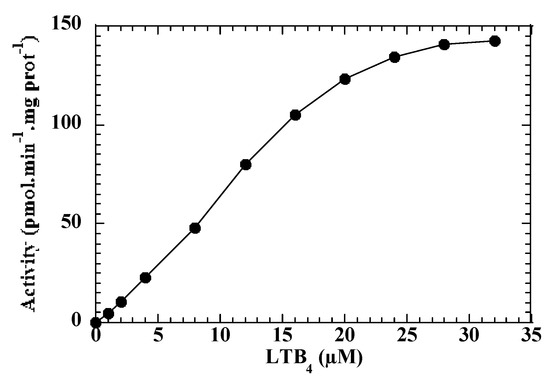
Figure 2.
Dependence of ω-hydroxylase activity of yeast microsomes expressing CYP2U1 upon LTB4 concentration. Microsomes were incubated for 15 min at 28 °C with various concentrations of LTB4 in the presence of 1 mM NADPH, treated and analyzed by HPLC as indicated in Materials and Methods. Results from a representative experiment.
2.4. Competition with Fatty Acid Derivatives
Then, we studied if several biologically active fatty acid derivatives could inhibit the CYP2U1-dependent oxidation of LTB4. Among them, the LTB4 analogues 12-HETE and 1-oleoyl-LPA appeared as potent inhibitors of the reaction with ~60% inhibition at 50 µM and an almost complete inhibition at 200 μM. Interestingly, 5-HETE, LTE4 and LTD4 also significantly inhibited the reaction at 200 µM. By contrast, Palm-Ser appeared much less efficient (Table 1).

Table 1.
Effects of some fatty acids on the ω-hydroxylation of LTB4 catalyzed by yeast microsomes expressing CYP2U1 and CPR.
2.5. Molecular Docking of LTB4 at the Active Site
In order to better understand the origin of the regioselectivity of the CYP2U1-catalyzed hydroxylation of LTB4, we performed molecular docking experiments with CDOCKER [22] using our truncated 3D model of CYP2U1 [10,23]. All poses where LTB4 was located outside the active site and those placing its carboxylate or OH functions that are potential iron ligands near the iron of the heme were discarded. Among the 1760 poses obtained with CDOCKER energies between −80 and −48 kcal/mol, a cluster of 60 poses showed the C20-atom of the alkyl chain positioned close to the iron of the heme (Figure 3). The molecule was inserted at the level of the active site so as to place the C20-atom at a distance of 3.6 Å from the heme iron, with the C20-Fe segment almost perpendicular to the heme. LTB4 was in an extended conformation, folded around helix I and close to helices G and I. In all these poses, the charged carboxyl group of LTB4 interacted with Lys 292 and Thr 295 at the entrance of channel 2ac (following the nomenclature of Cojocaru et al. [24,25], distances between the Lys Nε-atom and the OH-Thr group and an O-atom of LTB4-carboxylate of about 2.9, 2.7 and 2.5 Å, respectively) (Figure 3). The alkyl chain of LTB4 established hydrophobic interactions with hydrophobic residues of Phe 167 (π-staking with the diene-moiety), Tyr 344, Ala 352, Leu 530 and Ile 351, all along channel 2ac. Interestingly, the O-atom carried by the C5-atom interacted with one of the H-atoms of Gly 347 (distance 2.55 Å). By contrast, no clear interaction could be observed between the OH-group at C12 of LTB4 and a residue of the protein. The positioning of LTB4 in our model thus displays similarities with that observed for AA [10,23] except for new interactions linked to the insertion of the C5-OH group. This could explain the low Km values measured for the two compounds.
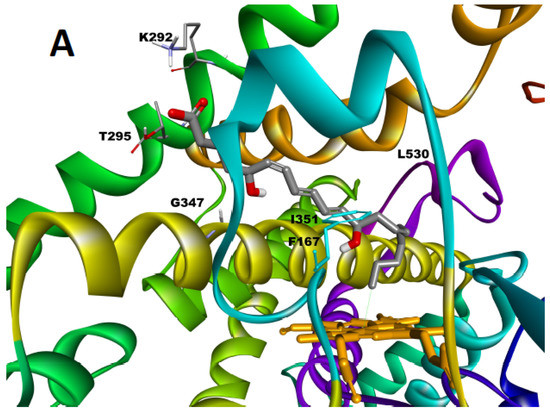
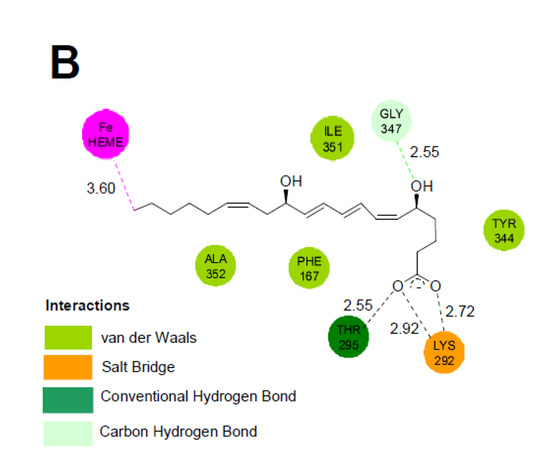
Figure 3.
Docking of LTB4 in the CYP2U1 3D model. (A): Pose illustrating the most energetically favored situation with the C20-atom of LTB4 close to the iron (distance Fe-C: 3.60 Å). The protein is shown in cartoon representation with the BC loop colored in blue, the helix I in yellow, helix G in green and helix F in orange. The heme (orange) and LTB4 (grey) are shown in stick representation with O-atoms in red. (B): 2D Representation of the predominant interactions between LTB4 and the protein.
3. Discussion
The inactivation of LTB4 is an important metabolic process for the regulation of its physiological and deleterious effects. The predominant pathway for LTB4 metabolism involves ω-oxidation to 20-OH-LTB4 predominantly catalyzed by cytochromes P450 of the 4F family that are mainly expressed in liver and neutrophiles [26,27,28,29]. Then, 20-OH-LTB4 can be re-oxidized to 20-CO2H-LTB4 and further eliminated by β-oxidation [30]. 20-OH-LTB4 and 20-CO2H-LTB4 are considered as almost inactive compounds [26]. In this study, we found that CYP2U1 is also able to hydroxylate LTB4 with major formation of 20-OH-LTB4 and minor formation of 19-OH- and 20-COOH-LTB4 (Scheme 1). LTB4 has a relatively good affinity for CYP2U1 as shown by the Km value found for its CYP2U1-dependent oxidation (12 µM), close to that described for AA (2.7 µM) [8] and higher than that described for arachidonoylserotonin (82 µM) [9]. Molecular docking results for LTB4 binding on a previously described 3D model of truncated CYP2U1 were in agreement with the regioselectivity found for its CYP2U1-dependent hydroxylation, with a cluster of poses showing the LTB4 C20 atom at 3.6 Å from the heme iron.
The involvement of CYP2U1 in the metabolism of LTB4 could have strong physiological consequences in cerebral pathological events including ischemic stroke because CYP2U1 is predominantly expressed in the brain whereas CYP4F2/4F3 are mainly expressed in liver and neutrophiles. The physiological consequences of the ω-oxidation of LTB4 by CYP2U1 in brain remains to be determined. In any case, this reaction should result in an inactivation of LTB4. Moreover, taking into account that CYP2U1 is mostly expressed in brain and that LTB4 is an important mediator in brain inflammatory disorders [18,19,20], this activity could correspond to a protective role of CYP2U1. Finally, the hydroxylation of LTB4 is easy to follow by a simple HPLC/UV-method. It could be used to screen molecules having a high affinity for CYP2U1. In this study, we found that some fatty acid derivatives such as 12-HETE, LTD4, LTE4 and 1-oleoyl-LPA can act as potent inhibitors of CYP2U1-dependent hydroxylation of LTB4. Further studies are in progress to characterize this extending range of substrates.
4. Materials and Methods
4.1. Materials
LTB4, 20-OH-LTB4, 20-CO2H-LTB4, LTD4, LTE4, 5-HETE, 12-HETE, palmitoylserotonin (Palm-Ser) and 1-oleoyl-LPA, came from Bertin-Pharma (Montigny-le-Bretonneux, France). 19-OH-LTB4 was prepared in our department by an adaptation of the synthetic scheme used for preparing LTB4 [31,32]. NADPH, N-benzyl-imidazole (Bz-ImH), prostaglandin B2 (PGB2) and all other chemicals were purchased from Sigma-Aldrich (Saint-Quentin Fallavier, France) and were of the highest commercially available purity. HPLC-grade acetonitrile was obtained from SDS (Peypin, France).
4.2. Expression of CYP2U1 in Yeast
Preparation of pYeDP60-cyp2U1 plasmid to express full-length CYP2U1 and human CYP reductase (CPR) in yeast Saccharomyces cerevisiae, strain W(hR), preparation of yeast microsomes, electrophoresis and western blot analysis for CYP2U1 were performed following previously published procedures [10]. The protein concentrations were measured by the Bradford method using bovine serum albumin as a standard [33] and the CYP concentrations were measured by UV-visible difference spectroscopy of the FeII-CO complex using an ε value of 91,000 M−1.cm−1 [34]. The typical preparations obtained displayed a CYP2U1 content of 55 ± 12 pmol.mg prot−1.
4.3. LTB4 Metabolism
Microsomes from yeast expressing CYP2U1 and human CPR were incubated with 1 mM NADPH and 20 μM LTB4 in a total volume of 100 μL of 50 mM phosphate buffer (pH 7.4) for 2–60 min at 28 °C. The reactions were stopped with the addition of 50 μL cold acetonitrile/acetic acid mixture (9/1, v/v) containing 6.0 µM PGB2 (internal standard), centrifuged at 13,000 rpm for 20 min and aliquots (20 μL) of supernatants were analyzed by HPLC-MS. Control experiments were carried out under identical conditions but either without protein, without NADPH, in the presence of 1 mM Bz-ImH, or with microsomes from yeast W(hR) transformed with an empty pYeDP60 plasmid [10].
The supernatants were analyzed on a Surveyor HPLC system including a PDA detector and coupled to a LCQ Advantage ion trap mass spectrometer (Thermo, Les Ulis, France). HPLC separations were achieved on a Gemini C18 column (100 × 2 mm, 3 μm; Phenomenex, Le Pecq, France). Elution (flow rate 250 μL/min) was performed using a mixture of solvent A (water + 0.1% formic acid) and B (acetonitrile + 0.1% formic acid) with the following gradient conditions: 10% B for 1 min, linear gradient to 80% B in 18 min, linear gradient to 100% B in 1 min, holding at 100% B for 3 min, returning to 10% B in 0.1 min, and re-equilibration at 10% B for 5 min. The column effluent was directed to the ion source from 3 to 23 min after injection to reduce contamination. Mass spectra were obtained by electrospray ionisation in the negative detection mode using the following parameters: capillary temperature, 250 °C; capillary voltage, 6 V; spray voltage, 4.5 kV; primary gas flow, 20 a.u.; auxillary gas flow, 5 a.u. Mass spectra were recorded with a resolution of 1 a.m.u., with a frequency of 20 Hz. The range of masses scanned for the total-ion chromatogram was m/z 120–600. For all products, the indicated molecular ions corresponded to M-H. After data acquisition, HPLC-MS chromatograms and UV-Vis spectra (230–550 nm) were analyzed with Xcalibur 2.2.0 software (Thermo, Les Ulis, France).
4.4. Computational
All computations were performed as previously described [10,23] using the software package Biovia Discovery Studio 2016 (Dassault Systems, Cambridge, UK). The parameters applied for the heme (defined as FeIII protoporphyrin IX) were obtained from Oda et al. [35]. A 3D model of CYP2U1 deprived of its predicted membrane-spanning domain (residues 57 to 544) was used as previously described [10,23]. Each of the three databases: Human Metabolome Database (HMDB, https://hmdb.ca/), ~2400 molecules, metabolites found in human); Cayman Chemical (https://www.caymanchem.com/), ~5000 molecules, predominantly lipids and eicosanoids); and Lipid Maps–Lipidomics (https://www.lipidmaps.org/) ~4500 molecules, fatty acids, glycerolipids) was downloaded. The compounds were first totally ionized (6.5 < pH < 8.5) to make them electrically neutral. Then, the tautomers and conformers for each molecule were generated and the ligands were screened on the CYP2U1 3D model. Poses were generated and their analysis allowed the selection of ligands according to their CDOCKER interaction energies (~−80 to −50 kcal/mol) [22] leading to compounds with a better affinity for the active site. Then, the positioning of the molecules close to the active site and the distance between the heme iron and oxidizable atoms (distance < 5 Å) were checked to select the most interesting compounds. From this study, 32 compounds were selected from the HMDB; 80 from Cayman and 130 from Lipidmaps. Manual analysis of the data resulted in the selection of ~10 compounds of high pharmacological interest. Among them, LTB4 appeared as an important target due to its key roles in inflammatory disorders. Molecular docking experiments of LTB4 at the active site were performed using default parameters from CDOCKER using CHARMm force-fields with Discovery Studio 2016. Molecular Graphics and analyses were performed with Discovery Studio 2016 also used for structure rendering and generation of figures.
Author Contributions
K.N. performed the preparation of yeast microsomes, the incubations, HPLC-MS analyses and computational experiments; N.P. and L.L.C. performed computational experiments; P.M.D. supervised the experiments and wrote the draft manuscript; D.M. and J.-L.B. supervised the experiments and wrote the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a doctoral fellowship from University Paris Descartes to K.N.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in this article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors acknowledge Patrice Gerardo (UMR 8601) for mass spectra analyses, Booma Ramassamy (UMR 8601) for technical assistance, and Julien Dairou (UMR 8601) for helpful discussions.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study, in the collection, analyses or interpretation of the data, in the writing of the manuscript or the decision to publish the results.
Abbreviations
| AA | arachidonic acid |
| Bz-ImH | N-benzyl-imidazole |
| CYP | cytochrome P450 |
| CYP2U1 | cytochrome P450 2U1 |
| CPR | cytochrome P450 reductase |
| HMDB | Human Metabolome Database |
| LTB4 | leukotriene B4 (5S,12R-dihydroxy-6Z,8E,10E,14Z-eicosatetraenoic acid) |
| LTD4 | leukotriene D4; LTE4, leukotriene E4 |
| 5-HETE | 5(S)-hydroxy-6E,8Z,11Z,14Z-eicosatetraenoic acid |
| 12-HETE | 12(S)-hydroxy-5Z,8Z,10E,14Z-eicosatetraenoic acid |
| PGB2 | prostaglandin B2 |
| Palm-Ser | palmitoylserotonin |
| 1-oleoyl-LPA | 1-O-9Z-octadecenoyl-sn-glyceryl-3-phosphoric acid |
References
- Guengerich, F.P. Human Cytochrome P450 Enzymes. In Cytochrome P450: Structure, Mechanism and Biochemistry, 3rd ed.; Ortiz de Montellano, P.R., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2005; pp. 377–530. [Google Scholar]
- Karlgren, M.; Backlund, M.; Johansson, L.; Oscarson, M.; Ingelman-Sundberg, M. Characterization and Tissue Distribution of a Novel Human Cytochrome P450-CYP2U1. Biochem. Biophys. Res. Commun. 2004, 315, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Karlgren, M.; Miura, M.; Ingelman-Sundberg, M. Novel Extra Hepatic Cytochrome P450s. Toxicol. Appl. Pharmacol. 2005, 207, 57–61. [Google Scholar] [CrossRef]
- Dhers, L.; Ducassou, L.; Boucher, J.L.; Mansuy, D. Cytochrome P450 2U1, a Very Peculiar Member of the Human P450s Family. Cell. Mol. Life Sci. 2017, 74, 1859–1869. [Google Scholar] [CrossRef] [PubMed]
- Devos, A.; Lino-Cardenas, C.L.; Glowacki, F.; Engels, A.; Lo-Guidice, J.M.; Chevalier, D.; Allorge, D.; Broly, F.; Cauffiez, C. Genetic Polymorphism of CYP2U1, a Cytochrome P450 Involved in Fatty Acids Hydroxylation. Prostaglandins Leukot. Essent. Fatty Acids 2010, 83, 105–110. [Google Scholar] [CrossRef]
- Toselli, F.; Booth Depaz, I.M.; Worrall, S.; Etheridge, N.; Dodd, P.R.; Wilce, P.A.; Gillam, E.M. Expression of CYP2E1 and CYP2U1 Proteins in Amygdala and Prefrontal Cortex: Influence of Alcoholism and Smoking. Alcohol Clin. Exp. Res. 2015, 39, 790–797. [Google Scholar] [CrossRef]
- Nelson, D.R. Comparison of P450s from Human and Fugu: 420 Million Years of Vertebrate P450 Evolution. Arch. Biochem. Biophys. 2003, 409, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.S.; Helvig, C.; Taimi, M.; Ramshaw, H.A.; Collop, A.H.; Amad, M.; White, J.A.; Petkovich, M.; Jones, G.; Korczak, B. CYP2U1, a Novel Human Thymus- and Brain-Specific Cytochrome P450, Catalyzes Omega- and (Omega-1)-Hydroxylation of Fatty Acids. J. Biol. Chem. 2004, 279, 6305–6314. [Google Scholar] [CrossRef]
- Siller, M.; Goyal, S.; Yoshimoto, F.K.; Xiao, Y.; Wei, S.; Guengerich, F.P. Oxidation of Endogenous N-Arachidonoylserotonin by Human Cytochrome P450 2U1. J. Biol. Chem. 2014, 289, 10476–10487. [Google Scholar] [CrossRef] [PubMed]
- Ducassou, L.; Jonasson, G.; Dhers, L.; Pietrancosta, N.; Ramassamy, B.; Xu-Li, Y.; Loriot, M.A.; Beaune, P.; Bertho, G.; Lombard, M.; et al. Expression in Yeast, New Substrates, and Construction of a First 3D Model of Human Orphan Cytochrome P450 2U1: Interpretation of Substrate Hydroxylation Regioselectivity from Docking Studies. Biochim. Biophys. Acta 2015, 1850, 1426–1437. [Google Scholar] [CrossRef] [PubMed]
- Tesson, C.; Nawara, M.; Salih, M.A.; Rossignol, R.; Zaki, M.S.; Al Balwi, M.; Schule, R.; Mignot, C.; Obre, E.; Bouhouche, A.; et al. Alteration of Fatty-Acid-Metabolizing Enzymes Affects Mitochondrial Form and Function in Hereditary Spastic Paraplegia. Am. J. Human Genet. 2013, 91, 1051–1064. [Google Scholar] [CrossRef] [PubMed]
- Durand, C.M.; Dhers, L.; Tesson, C.; Tessa, A.; Fouillen, L.; Jacqueré, S.; Raymond, L.; Coupry, I.; Benard, G.; Darios, F.; et al. CYP2U1 Activity is Altered by Missense Mutations in Hereditary Spastic Paraplegia 56. Hum. Mutat. 2018, 39, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Pujol, C.; Legrand, A.; Parodi, L.; Thomas, P.; Mochel, F.; Saracino, D.; Coarelli, G.; Croon, M.; Popovic, M.; Valet, M.; et al. Implication of Folate Deficiency in CYP2U1 Loss of Function. J. Exp. Med. 2021, 218, e20210846. [Google Scholar] [CrossRef] [PubMed]
- Borgeat, P.; Samuelsson, B. Metabolism of Arachidonic Acid in Polymorphonuclear Leukocytes. Structural Analysis of Novel Hydroxylated Compounds. J. Biol. Chem. 1979, 254, 7865–7869. [Google Scholar] [CrossRef]
- Lindgren, J.A.; Hansson, G.; Samuelsson, B. Formation of Novel Hydroxylated Eicosatetraenoic Acids in Preparations of Human Polymorphonuclear Leukocytes. FEBS Lett. 1981, 128, 329–335. [Google Scholar] [CrossRef]
- Ford-Hutchinson, A.W.; Bray, M.A.; Doig, M.V.; Shipley, M.E.; Smith, M.J. Leukotriene B4, a Potent Chemokinetic and Aggregating Substance Released from Polymorphonuclear Leukocytes. Nature 1980, 286, 264–265. [Google Scholar] [CrossRef]
- Borgeat, P.; Naccache, P.H. Biosynthesis and Biological Activity of Leukotriene B4. Clin. Biochem. 1990, 23, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, J.; Kaisotra, A.; Turman, C.T.; Grill, R.J.; Dash, P.K.; Strobel, H.W. CYP4Fs Expression in Rat Brain Correlates with Changes in LTB4 Level after Traumatic Brain Injury. J. Neurotrauma 2008, 25, 1187–1194. [Google Scholar] [CrossRef]
- Hijioka, M.; Anan, J.; Ishibashi, H.; Kurauchi, Y.; Hisatsune, A.; Seki, T.; Koga, T.; Yokomizo, T.; Shimizu, T.; Katsuki, H. Inhibition of Leukotriene B4 Action Mitigates Intracerebral Hemorrhage-Associated Pathological Events in Mice. J. Pharm. Exp. Ther. 2017, 360, 399–408. [Google Scholar] [CrossRef]
- Jing Chan, S.; Ng, M.P.E.; Zhao, H.; Ng, G.J.L.; De Foo, C.; Wong, P.T.H.; Seet, R.C.S. Early and Sustained Increases in Leukotriene B4 Levels are Associated with Poor Clinical Outcome in Ischemia Stroke Patients. Neurotherapeutics 2020, 17, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Correia, M.A.; Ortiz de Montellano, P.R. Inhibition of Cytochrome P450 enzymes. In Cytochrome P450: Structure, Mechanism, and Biochemistry, 3rd ed.; Ortiz de Montellano, P.R., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2005; pp. 247–322. [Google Scholar]
- Wu, G.; Robertson, D.H.; Brooks, C.L.; Vieth, M. Detailed Analysis of Grid-Based Molecular Docking: A Case Study of CDOCKER-A CHARMm-Based MD Docking Algorithm. J. Compt. Chem. 2003, 24, 1549–1562. [Google Scholar] [CrossRef] [PubMed]
- Dhers, L.; Pietrancosta, N.; Ducassou, L.; Ramassamy, B.; Dairou, J.; Jaouen, M.; André, F.; Mansuy, D.; Boucher, J.L. Spectral and 3D Model Studies of the Interaction of Orphan Human Cytochrome P450 2U1 with Substrates and Ligands. Biochim. Biophys. Acta 2017, 1861, 3144–3153. [Google Scholar] [CrossRef] [PubMed]
- Cojocaru, V.; Winn, P.J.; Wade, R.C. The Ins and Outs of Cytochrome P450s. Biochim. Biophys. Acta 2007, 1770, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Cojocaru, V.; Balali-Mood, K.; Sansom, M.S.; Wade, R.C. Structure and Dynamics of the Membrane-Bound Cytochrome P450 2C9. PLoS Comput. Biol. 2011, 7, e1002152. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.; Lindgren, J.A.; Dahlen, S.E.; Hedqvist, P.; Samuelsson, B. Identification and Biological Activity of Novel Omega-Oxidized Metabolites of Leukotriene B4 from Human Leukocytes. FEBS Lett. 1981, 130, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Shak, S.; Goldstein, I.M. Omega-Oxidation is the Major Pathway for the Catabolism of Leukotriene B4 in Human Polymorphonuclear Leukocytes. J. Biol. Chem. 1984, 259, 10181–10187. [Google Scholar] [CrossRef]
- Shak, S.; Goldstein, I.M. Leukotriene B4 Omega-Hydroxylase in Human Polymorphonuclear Leukocytes. Partial Purification and Identification as a Cytochrome P-450. J. Clin. Investig. 1985, 76, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, Y.; Kusunose, E.; Kusunose, M. Characterization of Human Liver Leukotriene B4 Omega-Hydroxylase P450 (CYP4F2). J. Biochem. 2000, 127, 1047–1052. [Google Scholar] [CrossRef]
- Harper, T.W.; Garrity, M.J.; Murphy, R.C. Metabolism of Leukotriene B4 in Isolated Hepatocytes: Identification of a Novel 18-Carboxy-19,20-dinor Leukotriene B4 Metabolite. J. Biol. Chem. 1986, 261, 5414–5418. [Google Scholar] [CrossRef]
- Le Merrer, Y.; Bonnet, A.; Depezay, J.C. Synthesis of 19-Hydroxy LTB4, an Assumed Metabolite of Leukotriene B4. Tetrahedron Lett. 1988, 29, 2647–2650. [Google Scholar] [CrossRef]
- LeMerrer, Y.; Gravier-Pelletier, C.; Micas-Languin, D.; Mestre, F.; Dureault, A.; Depezay, J.C. Total Synthesis of Leukotriene B4 [(+)-LTB4] and Homo-LTB4 from D-Mannitol. J. Org. Chem. 1989, 54, 2409–2416. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Omura, T.; Sato, R. The Carbon Monoxide-Binding Pigment of Liver Microsomes. I Evidence for Its Hemoprotein Nature. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar] [CrossRef]
- Oda, A.; Yamaotsu, N.; Hirono, S. New AMBER Force Field Parameters of Heme Iron for Cytochrome P450s Determined by Quantum Chemical Calculations of Simplified Models. J. Comput. Chem. 2005, 26, 818–826. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).