

Deletion of POMT2 in Zebrafish Causes Degeneration of Photoreceptors
Abstract
:1. Introduction
2. Results
2.1. Pomt2 Mutation in Zebrafish Caused Diminished Expression of Matriglycan and EYS Binding
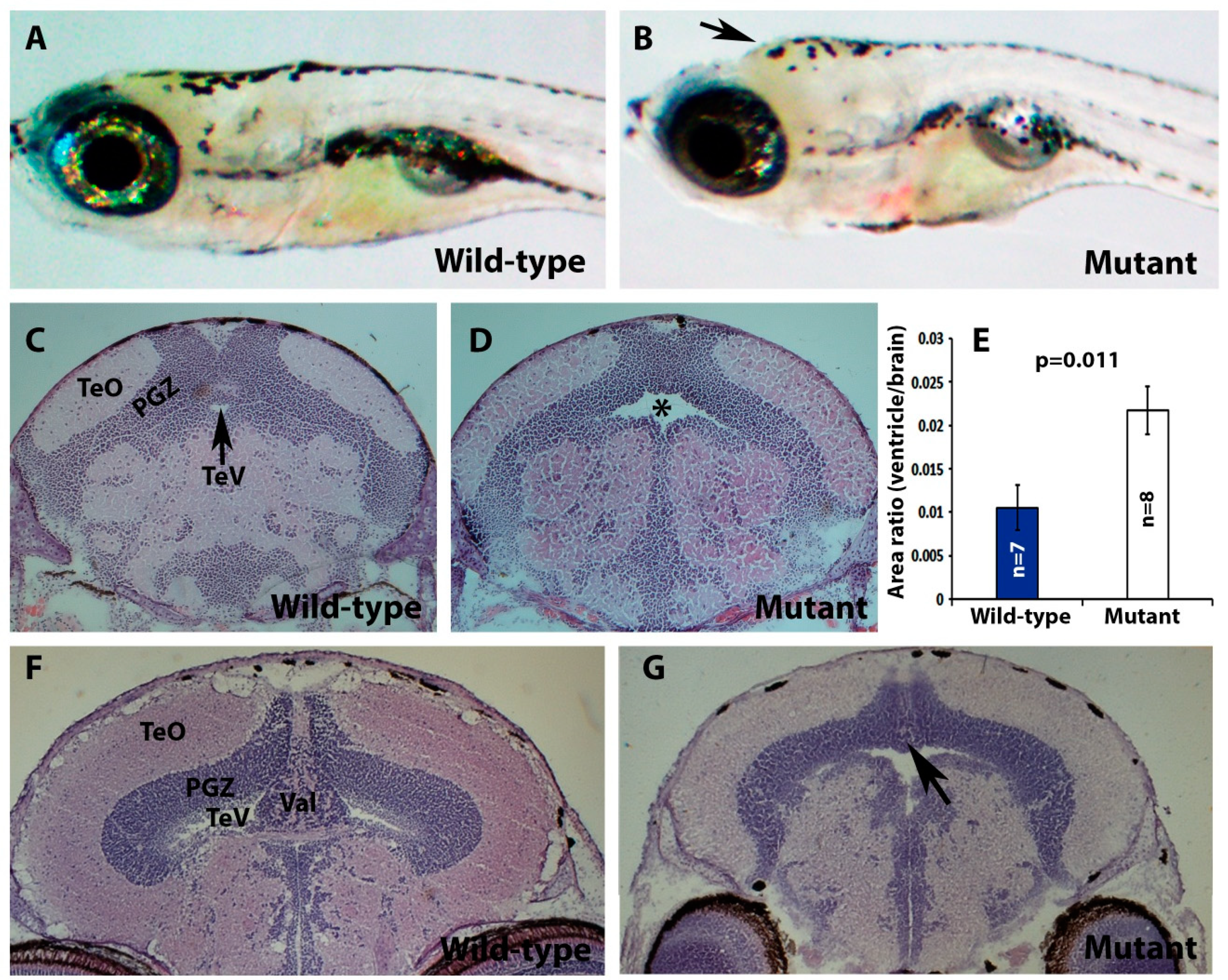
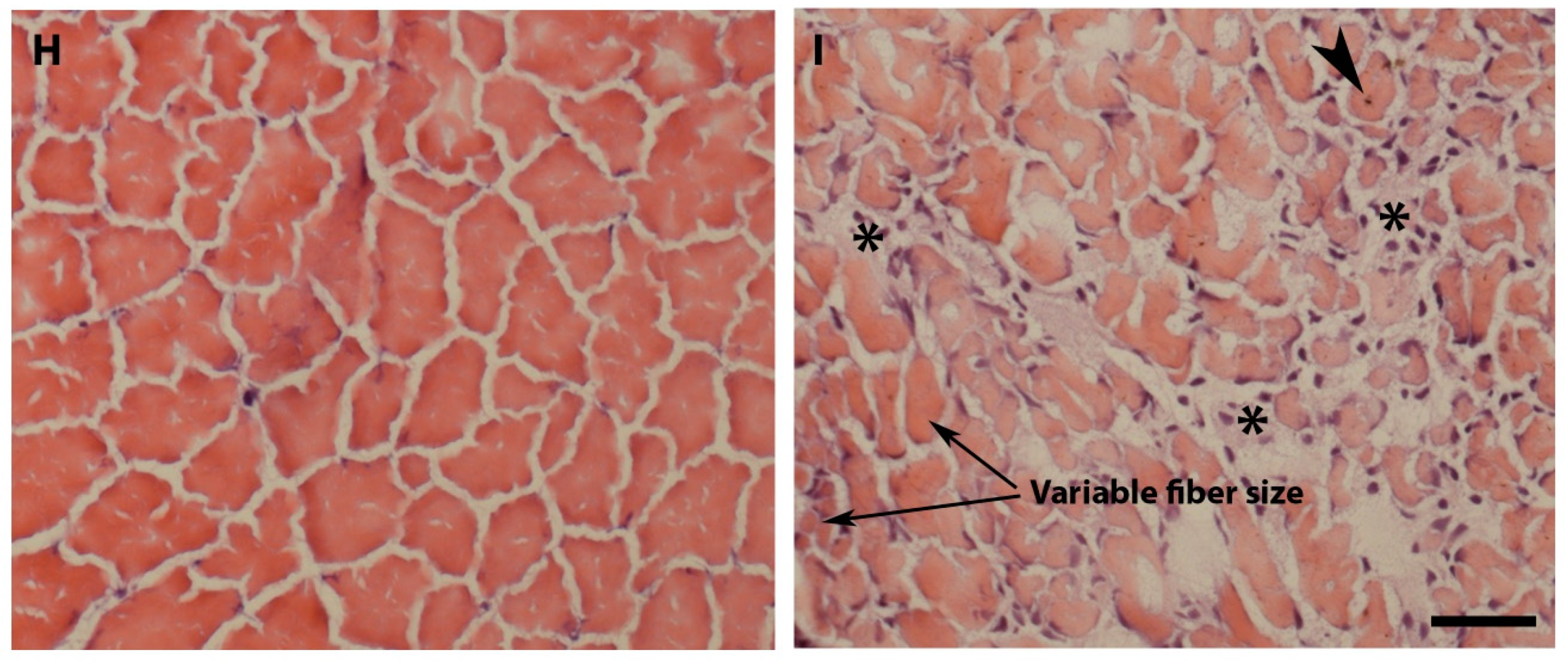
2.2. Pomt2 Mutant Zebrafish Exhibited Hydrocephalus, Hypoplasia of the Cerebellum, and Muscular Dystrophy
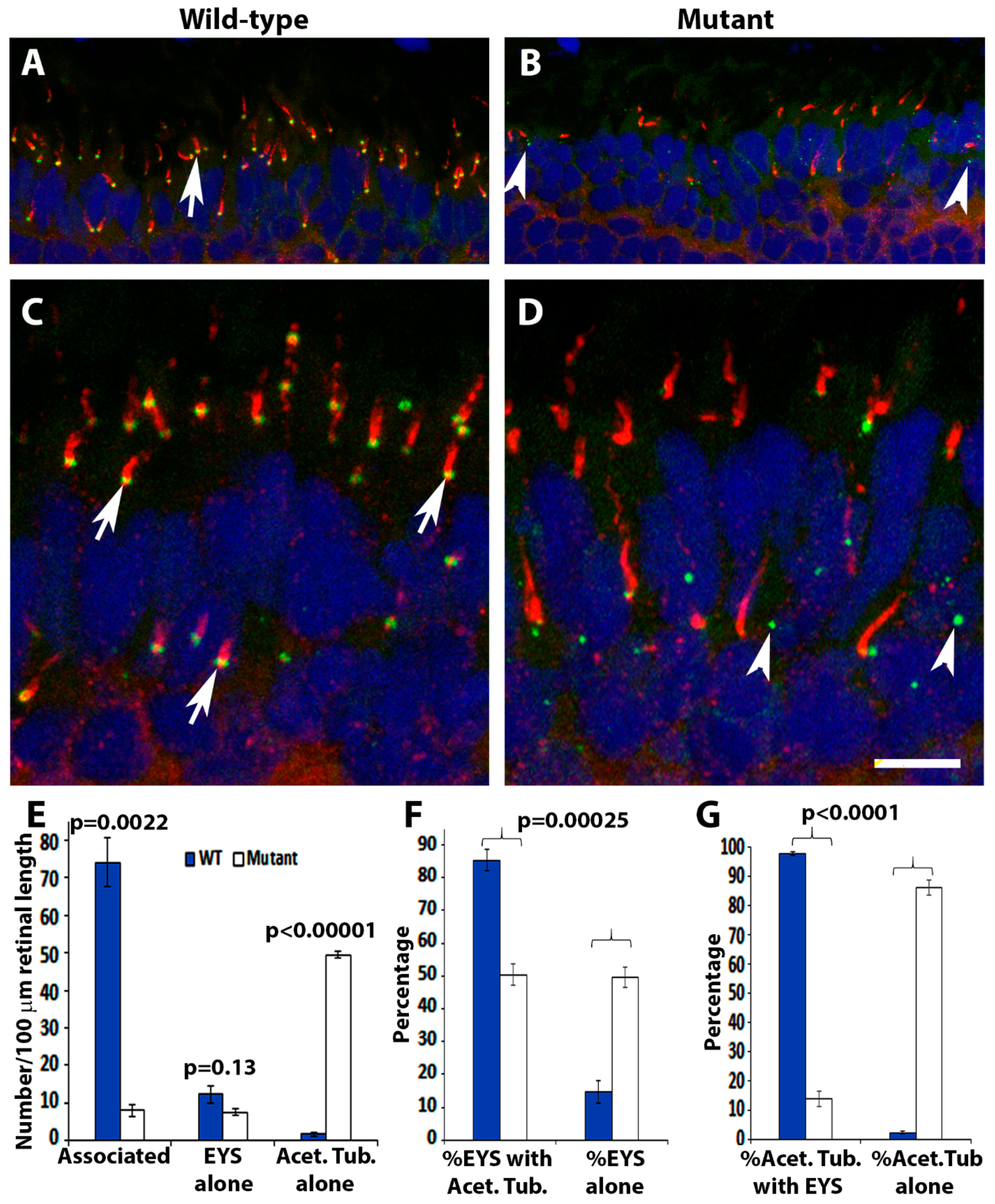
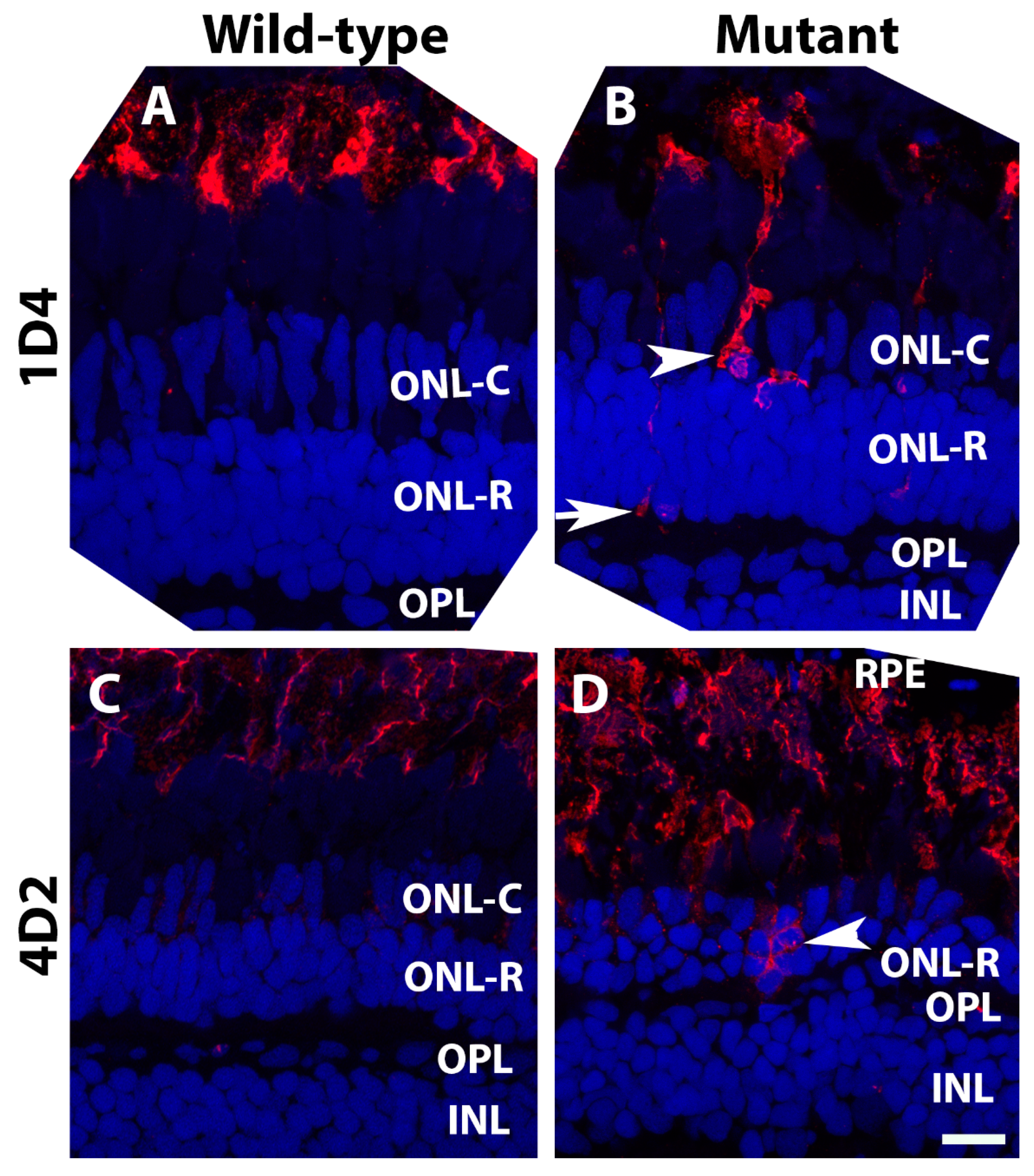
2.3. EYS Protein Was Reduced and Mislocalized in the Pomt2 Mutant Retina
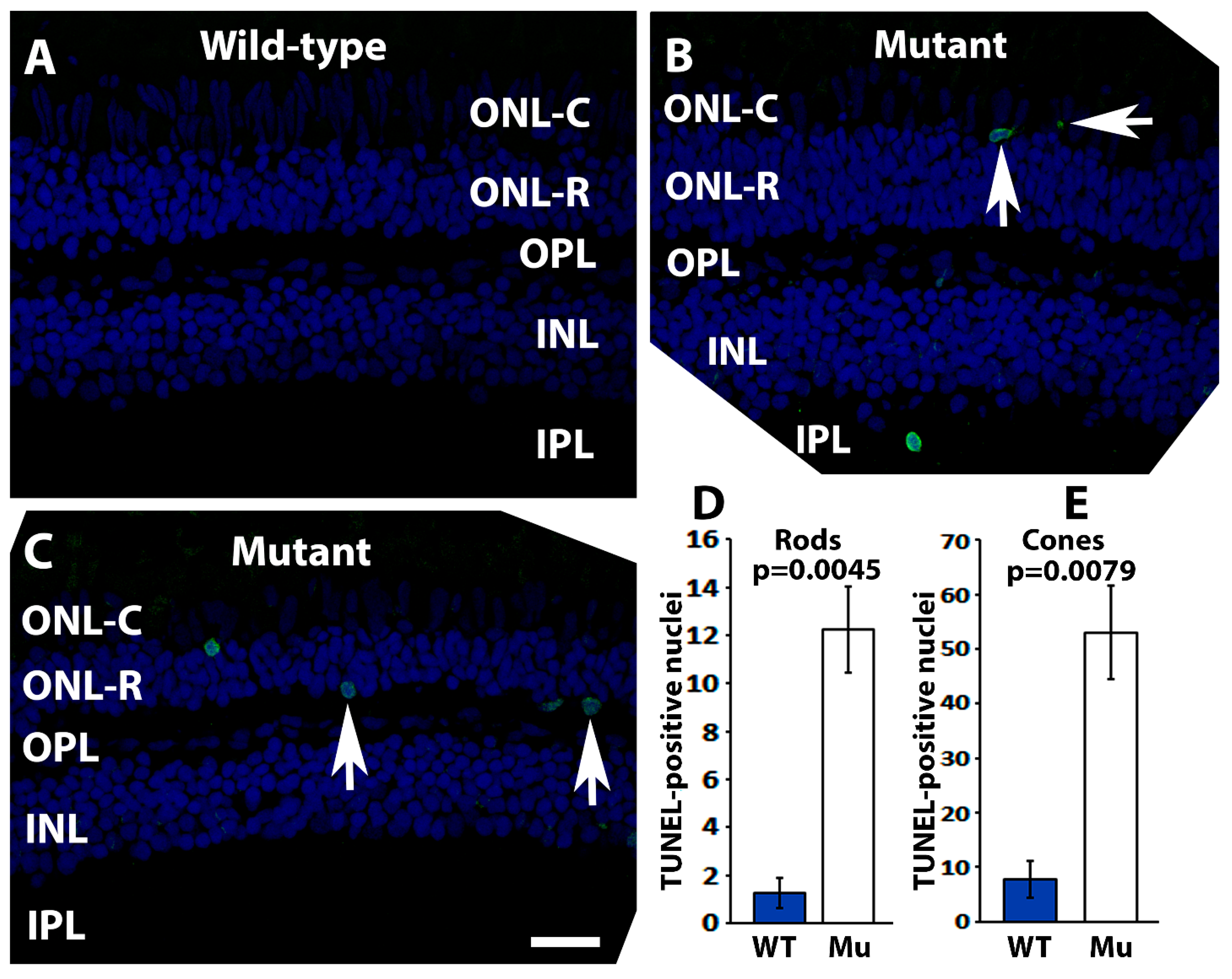
2.4. Pomt2 Mutant Zebrafish Exhibited Photoreceptor Degeneration
3. Discussion
4. Materials and Methods
4.1. Zebrafish Maintenance
4.2. Generation of the Pomt2sny5+13 Mutant Line
4.3. RT-PCR
4.4. Western Blotting Overlay Experiment
4.5. Immunofluorescence Staining
4.6. Semi-Quantitative Analyses Immunofluorescence Staining Signals
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Abd El-Aziz, M.M.; Barragan, I.; O’Driscoll, C.A.; Goodstadt, L.; Prigmore, E.; Borrego, S.; Mena, M.; Pieras, J.I.; El-Ashry, M.F.; Safieh, L.A.; et al. EYS, encoding an ortholog of Drosophila spacemaker, is mutated in autosomal recessive retinitis pigmentosa. Nat. Genet. 2008, 40, 1285–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, R.W.; Littink, K.W.; Klevering, B.J.; van den Born, L.I.; Koenekoop, R.K.; Zonneveld, M.N.; Blokland, E.A.; Strom, T.M.; Hoyng, C.B.; den Hollander, A.I.; et al. Identification of a 2 Mb human ortholog of Drosophila eyes shut/spacemaker that is mutated in patients with retinitis pigmentosa. Am. J. Hum. Genet. 2008, 83, 594–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandah-Rozenfeld, D.; Littink, K.W.; Ben-Yosef, T.; Strom, T.M.; Chowers, I.; Collin, R.W.; den Hollander, A.I.; van den Born, L.I.; Zonneveld, M.N.; Merin, S.; et al. Novel null mutations in the EYS gene are a frequent cause of autosomal recessive retinitis pigmentosa in the Israeli population. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4387–4394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littink, K.W.; van den Born, L.I.; Koenekoop, R.K.; Collin, R.W.; Zonneveld, M.N.; Blokland, E.A.; Khan, H.; Theelen, T.; Hoyng, C.B.; Cremers, F.P.; et al. Mutations in the EYS gene account for approximately 5% of autosomal recessive retinitis pigmentosa and cause a fairly homogeneous phenotype. Ophthalmology 2010, 117, 2026–2033. [Google Scholar] [CrossRef]
- Hosono, K.; Ishigami, C.; Takahashi, M.; Park, D.H.; Hirami, Y.; Nakanishi, H.; Ueno, S.; Yokoi, T.; Hikoya, A.; Fujita, T.; et al. Two novel mutations in the EYS gene are possible major causes of autosomal recessive retinitis pigmentosa in the Japanese population. PLoS ONE 2012, 7, e31036. [Google Scholar] [CrossRef]
- Audo, I.; Sahel, J.A.; Mohand-Said, S.; Lancelot, M.E.; Antonio, A.; Moskova-Doumanova, V.; Nandrot, E.F.; Doumanov, J.; Barragan, I.; Antinolo, G.; et al. EYS is a major gene for rod-cone dystrophies in France. Hum. Mutat. 2010, 31, E1406–E1435. [Google Scholar] [CrossRef] [Green Version]
- Barragan, I.; Borrego, S.; Pieras, J.I.; Gonzalez-del Pozo, M.; Santoyo, J.; Ayuso, C.; Baiget, M.; Millan, J.M.; Mena, M.; Abd El-Aziz, M.M.; et al. Mutation spectrum of EYS in Spanish patients with autosomal recessive retinitis pigmentosa. Hum. Mutat. 2010, 31, E1772–E1800. [Google Scholar] [CrossRef] [Green Version]
- Iwanami, M.; Oshikawa, M.; Nishida, T.; Nakadomari, S.; Kato, S. High prevalence of mutations in the EYS gene in Japanese patients with autosomal recessive retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Bonilha, V.L.; Rayborn, M.E.; Bell, B.A.; Marino, M.J.; Pauer, G.J.; Beight, C.D.; Chiang, J.; Traboulsi, E.I.; Hollyfield, J.G.; Hagstrom, S.A. Histopathological comparison of eyes from patients with autosomal recessive retinitis pigmentosa caused by novel EYS mutations. Graefes Arch. Clin. Exp. Ophthalmol. 2015, 253, 295–305. [Google Scholar] [CrossRef]
- Katagiri, S.; Akahori, M.; Hayashi, T.; Yoshitake, K.; Gekka, T.; Ikeo, K.; Tsuneoka, H.; Iwata, T. Autosomal recessive cone-rod dystrophy associated with compound heterozygous mutations in the EYS gene. Doc. Ophthalmol. 2014, 128, 211–217. [Google Scholar] [CrossRef]
- Husain, N.; Pellikka, M.; Hong, H.; Klimentova, T.; Choe, K.M.; Clandinin, T.R.; Tepass, U. The agrin/perlecan-related protein eyes shut is essential for epithelial lumen formation in the Drosophila retina. Dev. Cell 2006, 11, 483–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelhof, A.C.; Hardy, R.W.; Becker, A.; Zuker, C.S. Transforming the architecture of compound eyes. Nature 2006, 443, 696–699. [Google Scholar] [CrossRef]
- Yu, M.; Liu, Y.; Li, J.; Natale, B.N.; Cao, S.; Wang, D.; Amack, J.D.; Hu, H. Eyes shut homolog is required for maintaining the ciliary pocket and survival of photoreceptors in zebrafish. Biol. Open 2016, 5, 1662–1673. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Hu, X.; Liu, F.; Soares, D.C.; Liu, X.; Yu, S.; Gao, M.; Han, S.; Qin, Y.; Li, C.; et al. Ablation of EYS in zebrafish causes mislocalisation of outer segment proteins, F-actin disruption and cone-rod dystrophy. Sci. Rep. 2017, 7, 46098. [Google Scholar] [CrossRef] [PubMed]
- Messchaert, M.; Dona, M.; Broekman, S.; Peters, T.A.; Corral-Serrano, J.C.; Slijkerman, R.W.N.; van Wijk, E.; Collin, R.W.J. Eyes shut homolog is important for the maintenance of photoreceptor morphology and visual function in zebrafish. PLoS ONE 2018, 13, e0200789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Yu, M.; Shang, X.; Nguyen, M.H.H.; Balakrishnan, S.; Sager, R.; Hu, H. Eyes shut homolog (EYS) interacts with matriglycan of O-mannosyl glycans whose deficiency results in EYS mislocalization and degeneration of photoreceptors. Sci. Rep. 2020, 10, 7795. [Google Scholar] [CrossRef] [PubMed]
- Inamori, K.; Yoshida-Moriguchi, T.; Hara, Y.; Anderson, M.E.; Yu, L.; Campbell, K.P. Dystroglycan function requires xylosyl- and glucuronyltransferase activities of LARGE. Science 2012, 335, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Yoshida-Moriguchi, T.; Campbell, K.P. Matriglycan: A novel polysaccharide that links dystroglycan to the basement membrane. Glycobiology 2015, 25, 702–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ervasti, J.M.; Campbell, K.P. A role for the dystrophin-glycoprotein complex as a transmembrane linker between laminin and actin. J. Cell Biol. 1993, 122, 809–823. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Shimizu, T.; Tanaka, T.; Campbell, K.P.; Matsumura, K. Dystroglycan is a binding protein of laminin and merosin in peripheral nerve. FEBS Lett. 1994, 352, 49–53. [Google Scholar] [CrossRef]
- Gee, S.H.; Blacher, R.W.; Douville, P.J.; Provost, P.R.; Yurchenco, P.D.; Carbonetto, S. Laminin-binding protein 120 from brain is closely related to the dystrophin-associated glycoprotein, dystroglycan, and binds with high affinity to the major heparin binding domain of laminin. J. Biol. Chem. 1993, 268, 14972–14980. [Google Scholar] [CrossRef] [PubMed]
- Gee, S.H.; Montanaro, F.; Lindenbaum, M.H.; Carbonetto, S. Dystroglycan-alpha, a dystrophin-associated glycoprotein, is a functional agrin receptor. Cell 1994, 77, 675–686. [Google Scholar] [CrossRef]
- Peng, H.B.; Ali, A.A.; Daggett, D.F.; Rauvala, H.; Hassell, J.R.; Smalheiser, N.R. The relationship between perlecan and dystroglycan and its implication in the formation of the neuromuscular junction. Cell Adhes. Commun. 1998, 5, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Saito, F.; Tang, J.; Satz, J.; Campbell, K.; Sudhof, T.C. A stoichiometric complex of neurexins and dystroglycan in brain. J. Cell Biol. 2001, 154, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Wright, K.M.; Lyon, K.A.; Leung, H.; Leahy, D.J.; Ma, L.; Ginty, D.D. Dystroglycan organizes axon guidance cue localization and axonal pathfinding. Neuron 2012, 76, 931–944. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Omori, Y.; Katoh, K.; Kondo, M.; Kanagawa, M.; Miyata, K.; Funabiki, K.; Koyasu, T.; Kajimura, N.; Miyoshi, T.; et al. Pikachurin, a dystroglycan ligand, is essential for photoreceptor ribbon synapse formation. Nat. Neurosci. 2008, 11, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Kanagawa, M.; Omori, Y.; Sato, S.; Kobayashi, K.; Miyagoe-Suzuki, Y.; Takeda, S.; Endo, T.; Furukawa, T.; Toda, T. Post-translational maturation of dystroglycan is necessary for pikachurin binding and ribbon synaptic localization. J. Biol. Chem. 2010, 285, 31208–31216. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Li, J.; Zhang, Z.; Yu, M. Pikachurin interaction with dystroglycan is diminished by defective O-mannosyl glycosylation in congenital muscular dystrophy models and rescued by LARGE overexpression. Neurosci. Lett. 2011, 489, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Endo, T. Glycobiology of alpha-dystroglycan and muscular dystrophy. J. Biochem. 2015, 157, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Yamada, T.; Sun, Z.; Eblimit, A.; Lopez, I.; Wang, F.; Manya, H.; Xu, S.; Zhao, L.; Li, Y.; et al. Mutations in POMGNT1 cause non-syndromic retinitis pigmentosa. Hum. Mol. Genet. 2016, 25, 1479–1488. [Google Scholar] [CrossRef]
- Wang, N.H.; Chen, S.J.; Yang, C.F.; Chen, H.W.; Chuang, H.P.; Lu, Y.H.; Chen, C.H.; Wu, J.Y.; Niu, D.M.; Chen, Y.T. Homozygosity Mapping and Whole-Genome Sequencing Links a Missense Mutation in POMGNT1 to Autosomal Recessive Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3601–3609. [Google Scholar] [CrossRef] [Green Version]
- Manya, H.; Endo, T. Glycosylation with ribitol-phosphate in mammals: New insights into the O-mannosyl glycan. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2462–2472. [Google Scholar] [CrossRef] [PubMed]
- van Reeuwijk, J.; Janssen, M.; van den Elzen, C.; Beltran-Valero de Bernabe, D.; Sabatelli, P.; Merlini, L.; Boon, M.; Scheffer, H.; Brockington, M.; Muntoni, F.; et al. POMT2 mutations cause alpha-dystroglycan hypoglycosylation and Walker-Warburg syndrome. J. Med. Genet. 2005, 42, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Brocher, J.; Linder, B.; Hirmer, A.; Sundaramurthi, H.; Fischer, U.; Winkler, C. The 1D4 antibody labels outer segments of long double cone but not rod photoreceptors in zebrafish. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4943–4951. [Google Scholar] [CrossRef] [Green Version]
- Longman, C.; Brockington, M.; Torelli, S.; Jimenez-Mallebrera, C.; Kennedy, C.; Khalil, N.; Feng, L.; Saran, R.K.; Voit, T.; Merlini, L.; et al. Mutations in the human LARGE gene cause MDC1D, a novel form of congenital muscular dystrophy with severe mental retardation and abnormal glycosylation of alpha-dystroglycan. Hum. Mol. Genet. 2003, 12, 2853–2861. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, K.; Sawaki, H.; Sakai, T.; Hiruma, T.; Nakanishi, N.; Sato, T.; Ohkura, T.; Narimatsu, H. LARGE2 facilitates the maturation of alpha-dystroglycan more effectively than LARGE. Biochem. Biophys. Res. Commun. 2005, 329, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Hohenester, E. Structural biology of laminins. Essays Biochem. 2019, 63, 285–295. [Google Scholar] [PubMed]
- Khan, M.I.; Collin, R.W.; Arimadyo, K.; Micheal, S.; Azam, M.; Qureshi, N.; Faradz, S.M.; den Hollander, A.I.; Qamar, R.; Cremers, F.P. Missense mutations at homologous positions in the fourth and fifth laminin A G-like domains of eyes shut homolog cause autosomal recessive retinitis pigmentosa. Mol. Vis. 2010, 16, 2753–2759. [Google Scholar]
- Siemiatkowska, A.M.; Arimadyo, K.; Moruz, L.M.; Astuti, G.D.; de Castro-Miro, M.; Zonneveld, M.N.; Strom, T.M.; de Wijs, I.J.; Hoefsloot, L.H.; Faradz, S.M.; et al. Molecular genetic analysis of retinitis pigmentosa in Indonesia using genome-wide homozygosity mapping. Mol. Vis. 2011, 17, 3013–3024. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age of Animals | +/+ | +/− | −/− | p Value * |
|---|---|---|---|---|
| 7-dpf | 23 | 20 | 12 | 0.59 |
| 14-dpf | 14 | 26 | 15 | 0.70 |
| 21-dpf | 20 | 21 | 15 | 0.76 |
| 1-mpf | 45 | 83 | 30 | 0.081 |
| 2-mpf | 28 | 62 | 14 | 0.0066 |
| 3–6-mpf | 169 | 264 | 38 | 2.13 × 10−17 |
| Age Range | +/+ | +/− | −/− | p Value * |
|---|---|---|---|---|
| 21 to 35 dpf | 2 | 17 | 12 | 0.034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Rittershaus, J.M.; Yu, M.; Sager, R.; Hu, H. Deletion of POMT2 in Zebrafish Causes Degeneration of Photoreceptors. Int. J. Mol. Sci. 2022, 23, 14809. https://doi.org/10.3390/ijms232314809
Liu Y, Rittershaus JM, Yu M, Sager R, Hu H. Deletion of POMT2 in Zebrafish Causes Degeneration of Photoreceptors. International Journal of Molecular Sciences. 2022; 23(23):14809. https://doi.org/10.3390/ijms232314809
Chicago/Turabian StyleLiu, Yu, Jaclyn M. Rittershaus, Miao Yu, Rachel Sager, and Huaiyu Hu. 2022. "Deletion of POMT2 in Zebrafish Causes Degeneration of Photoreceptors" International Journal of Molecular Sciences 23, no. 23: 14809. https://doi.org/10.3390/ijms232314809