


Understanding Life at High Temperatures: Relationships of Molecular Channels in Enzymes of Methanogenic Archaea and Their Growth Temperatures

Abstract
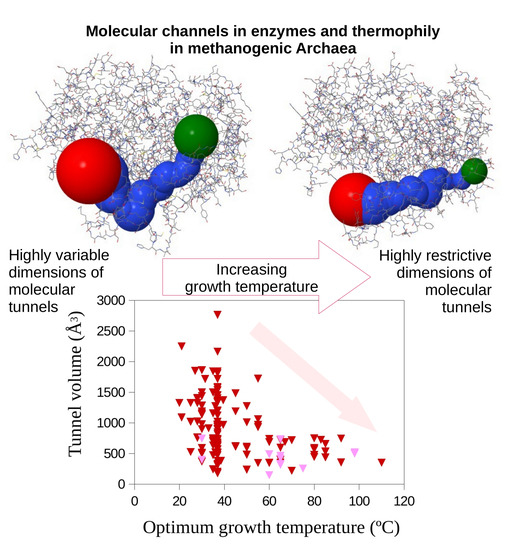
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miles, E.W.; Rhee, S.; Davies, D.R. The molecular basis of substrate channeling. J. Biol. Chem. 1999, 274, 12193–12196. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.X.; McCammon, J.A. The gates of ion channels and enzymes. Trends Biochem. Sci. 2010, 35, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Hubner, C.A.; Jentsch, T.J. Ion channel diseases. Hum. Mol. Genet. 2002, 11, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Holden, H.M.; Raushel, F.M. Channeling of substrates and intermediates in enzyme-catalyzed reactions. Annu. Rev. Biochem. 2001, 70, 149–180. [Google Scholar] [CrossRef]
- Kingsley, L.; Lill, M.A. Substrate tunnels in enzymes: Structure-function relationships and computational methodology. Proteins 2015, 83, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Pravda, L.; Sehnal, D.; Vařeková, R.S.; Navrátilova, V.; Toušek, D.; Berka, K.; Otyepka, M.; Koca, J. ChannelsDB: Database of biomacromolecular tunnels and pores. Nucleic Acids Res. 2018, 46, D399–D405. [Google Scholar] [CrossRef]
- Pravda, L.; Berka, K.; Svobodová Vařeková, R.; Sehnal, D.; Banáš, P.; Laskowski, R.A.; Koča, J.; Otyepka, M. Anatomy of enzyme channels. BMC Bioinform. 2014, 15, 379. [Google Scholar] [CrossRef] [PubMed]
- Milani, M.; Pesce, A.; Bolognesi, M.; Bocedi, A.; Ascenzi, P. Substrate channeling. Biochem. Mol. Biol. Educ. 2003, 31, 228–233. [Google Scholar] [CrossRef]
- Raushel, F.M.; Thoden, J.B.; Holden, H.M. Enzymes with molecular tunnels. Acc. Chem. Res. 2003, 36, 539–548. [Google Scholar] [CrossRef]
- Furmanová, K.; Vavra, O.; Kozlíková, B.; Damborský, J.; Vonásek, V.; Bednái, D.; Byska, J. DockVis: Visual analysis of molecular docking trajectories. Comput. Graph. Forum 2020, 39, 452–464. [Google Scholar] [CrossRef]
- Vavra, O.; Damborsky, J.; Bednar, D. Fast approximative methods for study of ligand transport and rational design of improved enzymes for biotechnologies. Biotech. Adv. 2022, 60, 108009. [Google Scholar] [CrossRef] [PubMed]
- Cuecas, A.; Cruces, J.; Galisteo-López, J.F.; Peng, X.; Gonzalez, J.M. Cellular viscosity in Prokaryotes and thermal stability of low molecular weight biomolecules. Biophys. J. 2016, 111, 875–882. [Google Scholar] [CrossRef]
- Grogan, D.W. Hyperthermophiles and the problem of DNA instability. Mol. Microbiol. 1998, 28, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev. 2000, 65, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Berezovsky, I.N.; Shakhnovich, E.I. Physics and evolution of thermophilic adaptation. Proc. Natl. Acad. Sci. USA 2005, 102, 12742–12747. [Google Scholar] [CrossRef]
- Daniel, R.M.; Cowan, D.A. Biomolecular stability and life at high temperatures. Cell. Mol. Life Sci. 2000, 57, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Cowan, D.A. The upper temperature of life—Where do we draw the line? Trends Microbiol. 2004, 12, 58–60. [Google Scholar] [CrossRef]
- Brock, T.D. Life at high temperatures. Science 1967, 158, 1012–1019. [Google Scholar] [CrossRef]
- Stetter, K.O.; Fiala, G.; Huber, G.; Huber, R.; Segerer, A. Hyperthermophilic microorganisms. FEMS Microbiol. Rev. 1990, 75, 117–124. [Google Scholar] [CrossRef]
- Gonzalez, J.M. Molecular tunnels in enzymes and thermophily: A case study on the relationship to growth temperature. Microorganisms 2018, 6, 109. [Google Scholar] [CrossRef]
- Ferry, J.G. Fundamentals of methanogenic pathways that are key to the biomethanation of complex biomass. Curr. Opin. Biotechnol. 2011, 22, 351–357. [Google Scholar] [CrossRef] [PubMed]
- González, J.M.; Stres, B. Trace element enzymes in reactions essential for anaerobic digestion. In Trace Elements in Anaerobic Biotechnologies; Chapter 3; Fermoso, F.G., van Hullebusch, E., Collins, G., Roussel, J., Mucha, A.P., Esposito, G., Eds.; IWA Publishing: London, UK, 2019; pp. 51–72. ISBN 9781789060225. [Google Scholar] [CrossRef]
- Bapteste, E.; Brochier, C.; Boucher, Y. Higher-level classification of the Archaea: Evolution of methanogenesis and methanogens. Archaea 2005, 1, 353–363. [Google Scholar] [CrossRef]
- Vishnivetskaya, T.A.; Buongiorno, J.; Bird, J.; Krivushin, K.; Spirina, E.V.; Oshurkova, V.; Shcherbakova, V.A.; Wilson, G.; Lloyd, K.G.; Rivkina, E.M. Methanogens in the Antartic dry Valley permafrost. FEMS Microbiol. Ecol. 2018, 94, fiy109. [Google Scholar] [CrossRef]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. USA 2008, 105, 10949–10954. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Robb, F.T.; Adams, M.W. Purification and characterization of NADP-specific alcohol dehydrogenase and glutamate dehydrogenase from the hyperthermophilic archaeon Thermococcus litoralis. Appl. Environ. Microbiol. 1994, 60, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Britton, K.L.; Yip, K.S.; Sedelnikova, S.E.; Stillman, T.J.; Adams, M.W.; Ma, K.; Maeder, D.L.; Robb, F.T.; Tolliday, N.; Vetriani, C.; et al. Structure determination of the glutamate dehydrogenase from the hyperthermophile Thermococcus litoralis and its comparison with that from Pyrococcus furiosus. J. Mol. Biol. 1999, 12, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G. Enzymology of one carbon metabolism in methanogenic pathways. FEMS Microbiol. Rev. 1999, 23, 13–38. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Petřek, M.; Košinová, P.; Koča, J.; Otyepka, M. MOLE: A voronoi diagrama-based explorer of molecular channels, pores, and tunnels. Structure 2007, 15, 1357–1363. [Google Scholar] [CrossRef]
- Masood, T.B.; Sandhya, S.; Chandra, N.; Natarajan, V. CHEXVIS: A tool for molecular channel extraction and visualization. BMC Bioinform. 2015, 16, 119. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. The Principles and Practice of Statistics in Biological Research, 7th ed.; W.H. Freeman and Co.: New York, NY, USA, 2001. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ginsbach, L.F.; Gonzalez, J.M. Understanding Life at High Temperatures: Relationships of Molecular Channels in Enzymes of Methanogenic Archaea and Their Growth Temperatures. Int. J. Mol. Sci. 2022, 23, 15149. https://doi.org/10.3390/ijms232315149
Ginsbach LF, Gonzalez JM. Understanding Life at High Temperatures: Relationships of Molecular Channels in Enzymes of Methanogenic Archaea and Their Growth Temperatures. International Journal of Molecular Sciences. 2022; 23(23):15149. https://doi.org/10.3390/ijms232315149
Chicago/Turabian StyleGinsbach, Laura F., and Juan M. Gonzalez. 2022. "Understanding Life at High Temperatures: Relationships of Molecular Channels in Enzymes of Methanogenic Archaea and Their Growth Temperatures" International Journal of Molecular Sciences 23, no. 23: 15149. https://doi.org/10.3390/ijms232315149
APA StyleGinsbach, L. F., & Gonzalez, J. M. (2022). Understanding Life at High Temperatures: Relationships of Molecular Channels in Enzymes of Methanogenic Archaea and Their Growth Temperatures. International Journal of Molecular Sciences, 23(23), 15149. https://doi.org/10.3390/ijms232315149