
Delineating Purinergic Signaling in Drosophila



Abstract
:1. Drosophila Made to Model and Understand
2. Benefits, Barriers, and Uncertainties in Drosophila Research
3. An Ensemble Approach to Purinergic Signaling
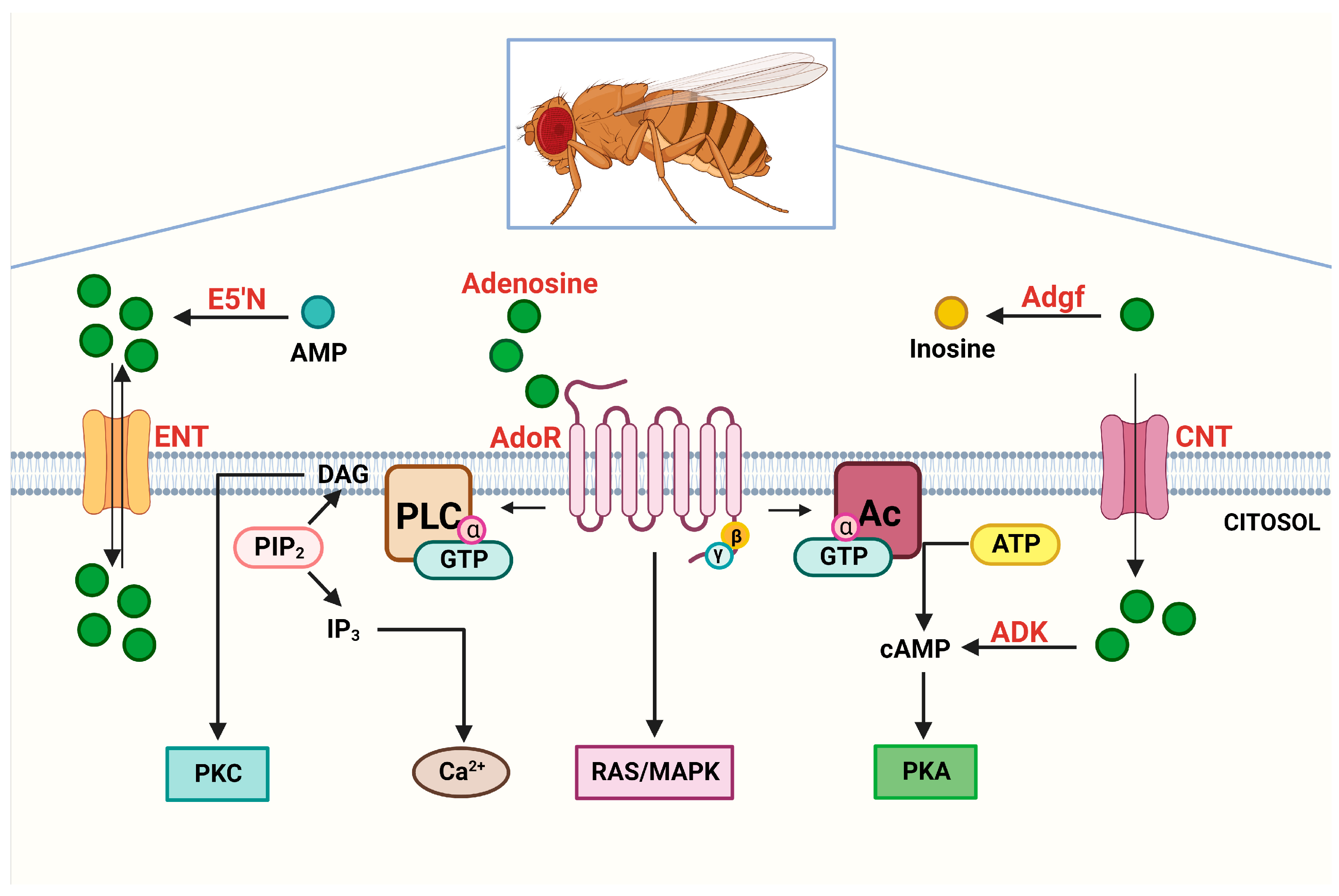
4. Underpinning Purinergic Signaling in Drosophila
4.1. Evidence about Adenosine Receptors
4.2. Evidence about Adenosine Transporters and Metabolic Enzymes
4.3. Evidence from Exogenous ATP Receptors Expression
5. Future Challenges and Opportunities
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Basic Notions about Drosophila
Appendix B. Basic Notions about Purinergic Transporters, Ectonucleotidases, and Receptors
References
- Verheyen, E.M. The power of Drosophila in modeling human disease mechanisms. Dis. Model. Mech. 2022, 15, dmm049549. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoskins, R.A.; Carlson, J.W.; Wan, K.H.; Park, S.; Mendez, I.; Galle, S.E.; Booth, B.W.; Pfeiffer, B.D.; George, R.A.; Svirskas, R.; et al. The Release 6 reference sequence of the Drosophila melanogaster genome. Genome Res. 2015, 25, 445–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şentürk, M.; Bellen, H.J. Genetic strategies to tackle neurological diseases in fruit flies. Curr. Opin. Neurobiol. 2018, 50, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Link, N.; Bellen, H.J. Using Drosophila to drive the diagnosis and understand the mechanisms of rare human diseases. Development 2020, 147, dev191411. [Google Scholar] [CrossRef]
- Flatt, T. Life-History Evolution and the Genetics of Fitness Components in Drosophila melanogaster. Genetics 2020, 214, 3–48. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liu, Z.; Rong, Y.S. Genome Editing: From Drosophila to Non-Model Insects and Beyond. J. Genet. Genom. 2016, 43, 263–272. [Google Scholar] [CrossRef]
- Burnstock, G.; Campbell, G.; Satchell, D.; Smythe, A. Evidence that adenosine triphosphate or a related nucleotide is the transmitter substance released by non-adrenergic inhibitory nerves in the gut. Br. J. Pharmacol. 1970, 40, 668–688. [Google Scholar] [CrossRef]
- Vultaggio-Poma, V.; Falzoni, S.; Salvi, G.; Giuliani, A.L.; Di Virgilio, F. Signalling by extracellular nucleotides in health and disease. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119237. [Google Scholar] [CrossRef]
- IJzerman, A.P.; Jacobson, K.A.; Müller, C.E.; Cronstein, B.N.; Cunha, R.A. International Union of Basic and Clinical Pharmacology. CXII: Adenosine Receptors: A Further Update. Pharmacol. Rev. 2022, 74, 340–372. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.; Iqbal, J. Purinergic receptors modulators: An emerging pharmacological tool for disease management. Med. Res. Rev. 2022, 42, 1661–1703. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.E.; Namasivayam, V. Recommended tool compounds and drugs for blocking P2X and P2Y receptors. Purinergic Signal. 2021, 17, 633–648. [Google Scholar] [CrossRef] [PubMed]
- Volonté, C.; Amadio, S.; D’Ambrosi, N.; Colpi, M.; Burnstock, G. P2 receptor web: Complexity and fine-tuning. Pharmacol. Ther. 2006, 112, 264–280. [Google Scholar] [CrossRef]
- Volonté, C.; D’Ambrosi, N. Membrane compartments and purinergic signalling: The purinome, a complex interplay among ligands, degrading enzymes, receptors and transporters. FEBS J. 2009, 276, 318–329. [Google Scholar] [CrossRef] [Green Version]
- Volonté, C.; Amadio, S. Rethinking purinergic concepts and updating the emerging role of P2X7 and P2X4 in amyotrophic lateral sclerosis. Neuropharmacology 2022, 221, 109278. [Google Scholar] [CrossRef]
- Riegel, J.A.; Maddrell, S.H.; Farndale, R.W.; Caldwell, F.M. Stimulation of fluid secretion of malpighian tubules of drosophila melanogaster meig. by cyclic nucleotides of inosine, cytidine, thymidine and uridine. J. Exp. Biol. 1998, 201 Pt 24, 3411–3418. [Google Scholar] [CrossRef]
- Brody, T.; Cravchik, A. Drosophila melanogaster G protein-coupled receptors. J. Cell Biol. 2000, 150, F83–F88. [Google Scholar] [CrossRef]
- Dolezelova, E.; Nothacker, H.P.; Civelli, O.; Bryant, P.J.; Zurovec, M. A Drosophila adenosine receptor activates cAMP and calcium signaling. Insect Biochem. Mol. Biol. 2007, 37, 318–329. [Google Scholar] [CrossRef]
- Kucerova, L.; Broz, V.; Fleischmannova, J.; Santruckova, E.; Sidorov, R.; Dolezal, V.; Zurovec, M. Characterization of the Drosophila adenosine receptor: The effect of adenosine analogs on cAMP signaling in Drosophila cells and their utility for In Vivo experiments. J. Neurochem. 2012, 121, 383–395. [Google Scholar] [CrossRef]
- Wu, M.N.; Ho, K.; Crocker, A.; Yue, Z.; Koh, K.; Sehgal, A. The effects of caffeine on sleep in Drosophila require PKA activity, but not the adenosine receptor. J. Neurosci. 2009, 29, 11029–11037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendricks, J.C.; Finn, S.M.; A Panckeri, K.; Chavkin, J.; A Williams, J.; Sehgal, A.; Pack, A. Rest in Drosophila is a sleep-like state. Neuron 2000, 25, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolezal, T.; Dolezelova, E.; Zurovec, M.; Bryant, P.J. A role for adenosine deaminase in Drosophila larval development. PLoS Biol. 2005, 3, e201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Franklin, B.; Tang, H.W.; Regimbald-Dumas, Y.; Hu, Y.; Ramos, J.; Bosch, J.A.; Villalta, C.; He, X.; Perrimon, N. An in vivo RNAi screen uncovers the role of AdoR signaling and adenosine deaminase in controlling intestinal stem cell activity. Proc. Natl. Acad. Sci. USA 2020, 117, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Poernbacher, I.; Vincent, J.P. Epithelial cells release adenosine to promote local TNF production in response to polarity disruption. Nat. Commun. 2018, 9, 4675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankar, N.; Machado, J.; Abdulla, P.; Hilliker, A.J.; Coe, I.R. Comparative genomic analysis of equilibrative nucleoside transporters suggests conserved protein structure despite limited sequence identity. Nucleic Acids Res. 2002, 30, 4339–4350. [Google Scholar] [CrossRef] [Green Version]
- Knight, D.; Harvey, P.J.; Iliadi, K.G.; Klose, M.K.; Iliadi, N.; Dolezelova, E.; Charlton, M.P.; Zurovec, M.; Boulianne, G.L. Equilibrative nucleoside transporter 2 regulates associative learning and synaptic function in Drosophila. J. Neurosci. 2010, 30, 5047–5057. [Google Scholar] [CrossRef] [Green Version]
- Zurovec, M.; Dolezal, T.; Gazi, M.; Pavlova, E.; Bryant, P.J. Adenosine deaminase-related growth factors stimulate cell proliferation in Drosophila by depleting extracellular adenosine. Proc. Natl. Acad. Sci. USA 2002, 99, 4403–4408. [Google Scholar] [CrossRef] [Green Version]
- Dolezelova, E.; Zurovec, M.; Dolezal, T.; Simek, P.; Bryant, P.J. The emerging role of adenosine deaminases in insects. Insect Biochem. Mol. Biol. 2005, 35, 381–389. [Google Scholar] [CrossRef]
- Machado, J.; Abdulla, P.; Hanna, W.J.; Hilliker, A.J.; Coe, I.R. Genomic analysis of nucleoside transporters in Diptera and functional characterization of DmENT2, a Drosophila equilibrative nucleoside transporter. Physiol. Genom. 2007, 28, 337–347. [Google Scholar] [CrossRef]
- Fenckova, M.; Hobizalova, R.; Fric, Z.F.; Dolezal, T. Functional characterization of ecto-5′-nucleotidases and apyrases in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2011, 41, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Prado Lopez, S.; Villamarìn, A.; Ibarguren, I. Simultaneous determination of adenosine and related purines in tissues and hemolymph of mussel by hplc. J. Liq. Chromatogr. Relat. Technol. 2012, 36, 470–485. [Google Scholar] [CrossRef]
- Fleischmannova, J.; Kucerova, L.; Sandova, K.; Steinbauerova, V.; Broz, V.; Simek, P.; Zurovec, M. Differential response of Drosophila cell lines to extracellular adenosine. Insect Biochem. Mol. Biol. 2012, 42, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Maaroufi, H.O.; Pauchova, L.; Lin, Y.H.; Wu, B.C.; Rouhova, L.; Kucerova, L.; Vieira, L.C.; Renner, M.; Sehadova, H.; Hradilova, M.; et al. Mutation in Drosophila concentrative nucleoside transporter 1 alters spermatid maturation and mating behavior. Front. Cell Dev. Biol. 2022, 10, 945572. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Maaroufi, H.O.; Kucerova, L.; Rouhova, L.; Filip, T.; Zurovec, M. Adenosine Receptor and Its Downstream Targets, Mod(mdg4) and Hsp70, Work as a Signaling Pathway Modulating Cytotoxic Damage in Drosophila. Front. Cell Dev. Biol. 2021, 9, 651367. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, T.; Gazi, M.; Zurovec, M.; Bryant, P.J. Genetic analysis of the ADGF multigene family by homologous recombination and gene conversion in Drosophila. Genetics 2003, 165, 653–666. [Google Scholar] [CrossRef]
- Sidorov, R.; Kucerova, L.; Kiss, I.; Zurovec, M. Mutation in the Drosophila melanogaster adenosine receptor gene selectively decreases the mosaic hyperplastic epithelial outgrowth rates in wts or dco heterozygous flies. Purinergic Signal. 2015, 11, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Zuberova, M.; Fenckova, M.; Simek, P.; Janeckova, L.; Dolezal, T. Increased extracellular adenosine in Drosophila that are deficient in adenosine deaminase activates a release of energy stores leading to wasting and death. Dis. Model. Mech. 2010, 3, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Chadwick, B.P.; Frischauf, A.M. The CD39-like gene family: Identification of three new human members (CD39L2, CD39L3, and CD39L4), their murine homologues, and a member of the gene family from Drosophila melanogaster. Genomics 1998, 50, 357–367. [Google Scholar] [CrossRef]
- Knowles, A.F. The GDA1_CD39 superfamily: NTPDases with diverse functions. Purinergic Signal. 2011, 7, 21–45. [Google Scholar] [CrossRef]
- Littleton, J.T.; Ganetzky, B. Ion channels and synaptic organization: Analysis of the Drosophila genome. Neuron 2000, 26, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Metpally, R.P.; Sowdhamini, R. Cross genome phylogenetic analysis of human and Drosophila G protein-coupled receptors: Application to functional annotation of orphan receptors. BMC Genom. 2005, 6, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, A.; Zhang, W.; Wang, Z. Functional feedback from mushroom bodies to antennal lobes in the Drosophila olfactory pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 10262–10267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.; Copeland, J.M.; Venton, B.J. Real-Time Measurement of Stimulated Dopamine Release in Compartments of the Adult. Anal. Chem. 2020, 92, 14398–14407. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.T.; Kallman, B.R.; Mullaney, B.C.; Scott, K. Representations of Taste Modality in the Drosophila Brain. Neuron 2015, 86, 1449–1460. [Google Scholar] [CrossRef] [Green Version]
- Grimes, L.; Griffiths, J.; Pasqualetto, G.; Brancale, A.; Kemp, P.J.; Young, M.T.; van der Goes van Naters, W. Drosophila taste neurons as an agonist-screening platform for P2X receptors. Sci. Rep. 2020, 10, 8292. [Google Scholar] [CrossRef]
- Lima, S.Q.; Miesenböck, G. Remote control of behavior through genetically targeted photostimulation of neurons. Cell 2005, 121, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Xiao, N.; Venton, B.J. Characterization of dopamine releasable and reserve pools in Drosophila larvae using ATP/P2X2 -mediated stimulation. J. Neurochem. 2015, 134, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Volonté, C.; Amadio, S.; Liguori, F.; Fabbrizio, P. Duality of P2X7 Receptor in Amyotrophic Lateral Sclerosis. Front. Pharmacol. 2020, 11, 1148. [Google Scholar] [CrossRef]
- Morgan, T.H. Sex limited inheritance in drosophila. Science 1910, 32, 120–122. [Google Scholar] [CrossRef]

{kind=link}
| Effector | Biological Target and/or Function | Ref. |
|---|---|---|
| Adenosine | Fluid secretion from Malpighian tubules | [17] |
| Adenosine | Cytotoxic effect | [28,29] |
| Adenosine | Concentration below 300 nM in haemolymph | [19] |
| Adenosine | Hyperglycaemia in larval haemolymph | [38] |
| Adenosine | Synergistic signaling of AdoR, Ent2, Adgf-A | [37] |
| Adenosine deaminases | Stimulation of cell proliferation | [28,29] |
| ADGF-A | Null mutation is lethal in homozygosis | [23,36] |
| AdoR | CG9753 gene identified as AdoR | [18] |
| AdoR | Endogenous brain expression and stress, immune, wake-cycle involvement | [22,23] |
| AdoR | CG9753 homology with ADORA2A | [19] |
| AdoR | AdoR mutants are viable | [21] |
| AdoR | AdoR coupling to adenylate cyclase | [20] |
| AdoR | Activation of TNF, boosting of JunK | [25] |
| AdoR | Multipotent intestinal stem cell maintenance | [24] |
| Caffeine | cAMP increase, PKA activation | [21] |
| CNT1 | Male infertility caused by mutations | [34] |
| ENT2 | Nucleoside transporters genomic analysis | [26] |
| ENT2 | Nucleoside transporter synaptic function, excitatory potentials increased by mutations | [27] |
| ENT2/AdoR | Amelioration of mutant Huntingtin-induced oxidative and heat stress response by knockdown | [35] |
| ENT2/CNT1/AdoR | Sequence homology with humans | [30] |
| ENT2/CNT1/AdoR | Rescue of imaginal discs cell deathby ablation | [33] |
| Ecto-5′-nucleotidases | Identification of five genes with alternative transcripts | [31] |
| NTDPase | Presence of just one CD39-like gene | [39] |
| NTDPase | Localization in the endoplasmic reticulum | [40] |
| P2X | Absence of P2X genes | [41] |
| P2Y | Absence of P2Y genes | [42] |
| Effector | Function induced by ectopic expression | Ref. |
| P2X2 | Locomotor activity and patterns | [47] |
| P2X2 | Olfactory information processing | [43] |
| P2X2 | Appetitive and aversive taste | [45] |
| P2X2 | Dopamine releasable and reserve pools | [48] |
| P2X2 | Novel agonists identification | [46] |
| P2X2 | Dopaminergic signaling in mushroom bodies | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volonté, C.; Alberti, F.; Vitale, G.; Liguori, F. Delineating Purinergic Signaling in Drosophila. Int. J. Mol. Sci. 2022, 23, 15196. https://doi.org/10.3390/ijms232315196
Volonté C, Alberti F, Vitale G, Liguori F. Delineating Purinergic Signaling in Drosophila. International Journal of Molecular Sciences. 2022; 23(23):15196. https://doi.org/10.3390/ijms232315196
Chicago/Turabian StyleVolonté, Cinzia, Francesca Alberti, Giuseppe Vitale, and Francesco Liguori. 2022. "Delineating Purinergic Signaling in Drosophila" International Journal of Molecular Sciences 23, no. 23: 15196. https://doi.org/10.3390/ijms232315196
APA StyleVolonté, C., Alberti, F., Vitale, G., & Liguori, F. (2022). Delineating Purinergic Signaling in Drosophila. International Journal of Molecular Sciences, 23(23), 15196. https://doi.org/10.3390/ijms232315196