Abstract
Understanding which intracellular signaling pathways are activated by manganese stress is crucial to decipher how metal overload compromise cellular integrity. Here, we unveil a role for oxidative and cell wall stress signaling in the response to manganese stress in yeast. We find that the oxidative stress transcription factor Yap1 protects cells against manganese toxicity. Conversely, extracellular manganese addition causes a rapid decay in Yap1 protein levels. In addition, manganese stress activates the MAPKs Hog1 and Slt2 (Mpk1) and leads to an up-regulation of the Slt2 downstream transcription factor target Rlm1. Importantly, Yap1 and Slt2 are both required to protect cells from oxidative stress in mutants impaired in manganese detoxification. Under such circumstances, Slt2 activation is enhanced upon Yap1 depletion suggesting an interplay between different stress signaling nodes to optimize cellular stress responses and manganese tolerance.
1. Introduction
Manganese is an essential, redox active metal of medical relevance. It serves as a metal co-factor for enzymes located in all cellular compartments and environmental exposure to manganese has been associated with the development of Parkinson-like neurological symptoms in human [1]. Budding yeast has been pivotal to identify genetic mutations affecting manganese metabolism and cellular availability. Manganese is sequestered by the vacuole or the Golgi, and Golgi manganese detoxification is mainly provided by the P-type Ca2+/Mn2+ ATPase Pmr1 [2]. The Pmr1 protein can be replaced by its human homolog SPCA1 encoded by the ATP2C1 gene [3] and mutations affecting calcium and/or manganese transport activities of SPCA1 are linked to Hailey-Hailey disease (HHD, [4]). Noteworthy, yeast cells lacking Pmr1 are sensitive to peroxide (H2O2; [5]) but at the same time, the absence of Pmr1 suppress oxygen-mediated phenotypes of superoxide dismutase 1 (Sod1) mutants [6].
To adapt to diverse extracellular stimuli and environmental changes, cells activate stress response pathways known to operate through sequential phosphorylation events that are termed protein kinase cascades. The stress-activated MAP kinases (SAPK) signaling cascades are evolutionary conserved prototypes of this kind of stress response signaling pathways. In yeast, the Hog1 SAPK plays a key role in reprogramming the gene expression pattern required for cell survival upon osmostress [7], while the Slt2 SAPK is the downstream kinase of the so-called cell wall integrity (CWI) pathway [8]. The human Hog1 paralog p38 plays a critical role in adaptive responses to environmental stress [9]. Slt2 is a functional homolog of human extracellular signal-regulated kinase 5 (ERK5), a MAPK that is activated in response to growth factors as well as physical and chemical stresses [10]. Together, Hog1 and Slt2 are needed to coordinate metabolic needs with cell cycle progression, therewith contributing to maintain genetic stability in yeast [11,12].
In an adaptive response to hydrogen peroxide (H2O2) treatment-mediated oxidative stress, the transcription factor yeast activator (AP1-like) protein (Yap1) is activated and translocates into the nucleus to induce the expression of a protective transcriptional program [13,14]. During metal and heat-shock induced oxidative stress, Yap1 transcriptionally regulates the expression of GSH1, which encodes the first enzyme (γ-glutamylcysteine synthetase) involved in glutathione biosynthesis, leading to an increase in intracellular glutathione level [15,16].
A growing body of evidence provides a link between the loss of P-type Golgi Ca2+/Mn2+ ATPases and genome instability in yeast and human. Loss of Pmr1 function leads to enhanced sensibility to a variety of DNA damaging agents, DNA damage formation and telomere shortening [5,17,18], while depletion of ATP2C1 in human cells was shown to boost the formation of reactive oxygen species (ROS) and to down-regulate the expression of DNA damage response (DDR) genes [19]. Finally, a genetic screen in Kluyveromyces lactis identified the Glutathione S-transferase ϴ-subunit (GTT1) as an oxidative stress suppressor in cells lacking Pmr1 [20], thereby providing a link between manganese and oxidative stress response.
These findings prompted us to further investigate stress response pathways related to oxidative damage and to determine if they contribute to manganese stress resistance in yeast. Here, we reveal that impaired manganese homeostasis leads to oxidative stress and that cellular tolerance to MnCl2 requires the Yap1 transcription factor, although MnCl2 addition stimulates Yap1 decay. Compromised manganese detoxification leads to a constitutive activation of the CWI effector MAPK Slt2, and Slt2 activation is further stimulated in the absence of Yap1. The activation of different stress signaling nodes pinpoints to a multifaceted impact of manganese on cellular metabolism that requires the concerted action of signaling kinases and transcription factors for manganese and oxidative stress tolerance.
2. Results
2.1. Impaired Manganese Homeostasis Leads to Oxidative Stress
Several observations relate Golgi Ca2+/Mn2+ transport to oxidative stress. The viability of yeast cells lacking a functional Pmr1 Golgi Ca2+/Mn2+ ATPase is compromised upon treatment with H2O2 [5] and oxidative stress and Notch1 activation were increased upon inactivation of the PMR1 homologue ATP2C1 in human cultured keratinocytes [19]. In yeast, several pathways participate in oxidative stress signaling, including the activation of the AP-1-like transcription factor Yap1 [21], and the SAPKs Hog1 and Slt2 [22,23] (see Figure 1A).
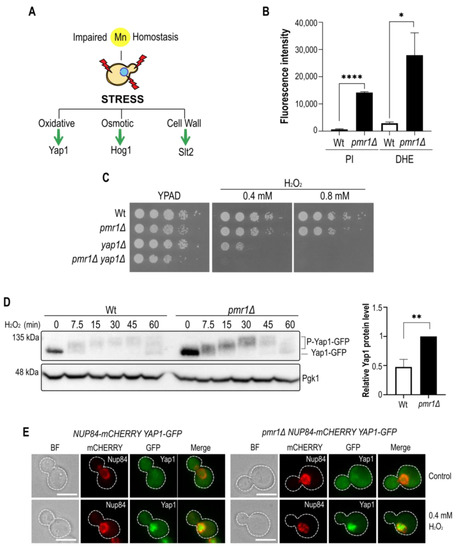
Figure 1.
The manganese tolerance and oxidative stress resistance of Pmr1 depleted cells depends on Yap1. (A) The schematic outline of stress signaling pathways linked to oxidative stress. The effector transcription factor (Yap1) or MAPKs (Hog1, Slt2) are indicated. (B) Flow cytometric analysis of DHE and PI positive cells. The fluorescence intensity of positive Wt and pmr1Δ mutant cells is plotted. Standard error of the mean (SEM, with bars) and p-values are indicated (p-value 0.05 *; 0.01 **; 0.001 ****). (C) Growth on peroxide-containing (H2O2) YPAD (Yeast Extract-Peptone-Adenine-Dextrose) medium. 10-fold dilutions of exponentially growing cells are shown. Strains and H2O2 concentrations are indicated. (D) Immunoblot of Yap1-GFP modifications after H2O2 addition in Wt and pmr1Δ mutants using an anti-GFP antibody. H2O2 concentration and incubation times (min) prior to protein isolation are indicated. Pgk1 (3-phosphoglycerate kinase 1) protein levels, probed with anti-Pgk1 antibodies, served as a loading control. Quantification of relative Yap1-GFP levels in untreated Wt and pmr1Δ cells (right panel). Standard deviation (SD, with bars) and p-value are indicated. (E) Analysis of Yap1-GFP localization by fluorescence microscopy. Cells were analyzed before (control) and after 7.5 min treatment with 0.4 mM H2O2. Nup84-mCherry was used as a nuclear marker. Scale bar represents 5 µm.
First, we assessed cell viability and ROS formation in pmr1Δ mutants using the fluorescent dyes propidium iodide (PI) and dihydroethidium (DHE), respectively (Figure 1B). As compared to wild-type cells (Wt), we found that loss of Pmr1 led to a considerable increase in the fluorescence signal with either dyes indicative of loss of plasma membrane selective permeability or increased ROS formation in pmr1Δ mutants. The later finding is in concordance with previous observations showing that DHE fluorescence is increased in pmr1Δ mutants [24]. Next, we wondered if oxidative stress tolerance in cells lacking Pmr1 may depend on Yap1, a transcription factor needed to activate the transcription of antioxidant genes in response to H2O2 [25,26]. In our strain background, no differences between Wt and pmr1Δ cells were observed in growth assays with H2O2 supplemented media (Figure 1C). However, mutant cells lacking Yap1 alone were very sensitive to H2O2 and pmr1Δ yap1Δ double mutants became even more H2O2-sensitive than yap1Δ single mutants, supporting the previous notion that pmr1Δ mutants are prone to oxidative stress. We then analyzed whether H2O2 induced Yap1 posttranslational modifications, observed as a shift in the electrophoretic mobility (Figure 1D). The cells were treated for up to 1 h with 400 µM H2O2 and protein samples were taken at various time points. In accordance with previous reports [27], Western blotting analysis revealed a rapid shift in Yap1 migration (7.5 min after treatment), suggesting an increase in Yap1 phosphorylation. While the kinetics of Yap1 protein modification was very similar in Wt and pmr1Δ cells, the initial levels of Yap1 protein appeared to be increased in cells lacking Pmr1 (Figure 1D). In addition, we monitored intracellular Yap1-GFP protein localization to assess if the cytoplasmic-nuclear Yap1 transition in response to H2O2 treatment is altered in pmr1Δ mutants (Figure 1E). Fluorescence microscopy analysis showed that this was not the case as the Yap1-GFP protein localization was mainly cytoplasmic in untreated Wt and pmr1Δ mutant cells but became principally nuclear in both cell types shortly after H2O2 treatment.
2.2. Manganese-Dependent Yap1 Decay Is Calcineurin B-Independent
Our finding that Yap1 is needed for oxidative stress resistance in the absence of Pmr1 led us to investigate whether or not Yap1 is required for growth on MnCl2 supplemented medium. To do so, we performed a cell growth analysis of cells lacking Pmr1 and/or Yap1 in growth medium supplemented with 1, 2.5 or 5 mM MnCl2 (Figure 2A). As expected, the growth of pmr1Δ mutant cells was strongly inhibited by MnCl2 addition. MnCl2 addition also impaired the growth of yap1Δ mutants as compared to Wt control cells. We therefore wondered if MnCl2 treatment would have an impact on Yap1 protein levels. Consequently, we performed Western blotting analyses of Wt and pmr1Δ cells expressing a Yap1-GFP protein upon treatment with MnCl2 (Figure 2B). MnCl2 treatment caused a significant reduction of Yap1-GFP protein levels both in Wt and pmr1Δ cells after the addition of 10 mM MnCl2. MnCl2 treatment did not alter the Yap1-GFP migration pattern suggesting that MnCl2 is not associated with Yap1 posttranslational modifications. In order to assess if the reduction of Yap1 protein levels is associated with a change in its subcellular localization, we monitored Yap1-GFP distribution by fluorescence microscopy (Figure 2C). However, in contrast to the nuclear Yap1-GFP accumulation observed in H2O2 treated cells, Yap1-GFP did not accumulate in the nucleus upon MnCl2 addition.
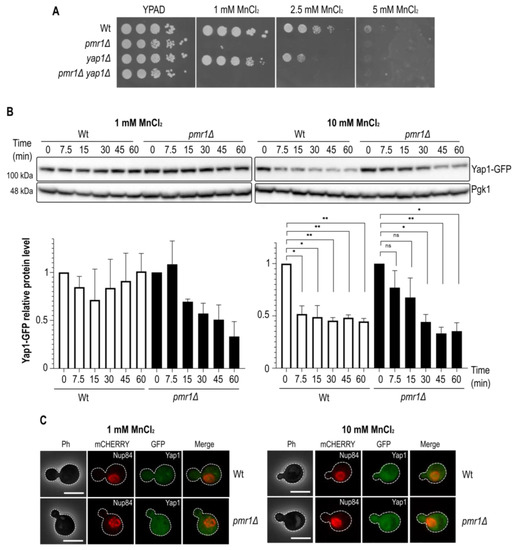
Figure 2.
Yap1 is a target for manganese stress response. (A) Growth on MnCl2-containing medium. 10-fold dilutions of exponentially growing cells are shown. The strains and MnCl2 concentrations are indicated. The pictures were taken after 3 days of growth. (B) Immunoblot of Yap1-GFP protein levels after MnCl2 addition in Wt and pmr1Δ mutants. MnCl2 concentration and incubation times (min) prior to protein isolation are indicated. Pgk1 (3-phosphoglycerate kinase 1) protein levels, probed with anti-Pgk1 antibodies, served as a loading control. Quantification of Yap1 protein levels is shown below. SEM with bars and p-value are indicated (p-value 0.05 *; 0.01 **). Time point 0 was set to 1. (C) Fluorescence microscopy analysis of Yap1-GFP localization. The cells were analyzed before and after 15 min treatment with 1 or 10 mM MnCl2. Nup84-mCherry was used as a nuclear marker. Scale bar represents 5 µm.
In order to distinguish whether Yap1 was down-regulated at the translational or post-translational level, we examined the effect of MnCl2 on the stability of Yap1 in the presence of protein synthesis inhibitor cycloheximide (CHX, Figure 3A). Notably, the decrease of Yap1-GFP protein levels was very similar in the presence of MnCl2 or CHX. No additive effect on Yap1 protein decay was observed in the presence of both MnCl2 and CHX suggesting that MnCl2 interferes with Yap1 protein synthesis. We next wondered if MnCl2-induced Yap1 decay is linked to the previously described CaCl2-dependent Yap1 decay due to activation of the calcium/calmodulin regulated serine/threonine protein phosphatase calcineurin [28]. Calcineurin is a heterodimeric enzyme comprising of a catalytic A and a regulatory B subunit, and is required for Crz1 transcription factor dependent gene expression, ion homeostasis and viability in yeast [29]. By taking advantage of calcineurin B (cnb1Δ) mutant cells we confirmed that the calcineurin B subunit is required for MnCl2 tolerance, in accordance with impaired activation of Crz1 target genes related to ion homeostasis (Figure 3B). However, in our strain background the CHX-induced Yap1 decay was barely reduced in cnb1Δ mutant cells, and MnCl2-dependent Yap1 decay was similar in Wt cells as compared to cnb1Δ mutants (Figure 3C). Further experimental settings will be needed to determine all the aspects of calcineurin-independent, MnCl2-driven Yap1-GFP decay.
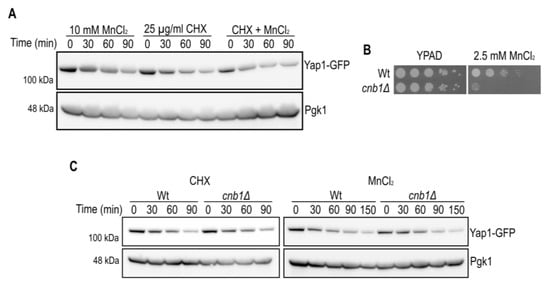
Figure 3.
Calcineurin B is not involved in MnCl2-driven Yap1 decay. (A) Immunoblot of Yap1-GFP protein levels after MnCl2 and/or CHX addition. The incubation times (min) prior to cell collection are indicated. Pgk1 (3-phosphoglycerate kinase 1) protein levels served as a loading control. (B) Growth on MnCl2-containing medium. 10-fold dilutions of exponentially growing cells are shown. Strains and MnCl2 concentrations are indicated. The pictures were taken after 2 days of growth. (C) Immunoblot of Yap1-GFP protein levels in Wt and cnb1Δ mutant cells after CHX (left panel) or MnCl2 (right panel) addition. See (A) for further description.
2.3. Pmr1 Depletion Leads to Constitutive Activation of the Slt2 MAP Kinase Pathway
MAPK cascades are among the major pathways by which extracellular stimuli are transduced into intracellular responses in eukaryotic cells. The yeast cell wall integrity (CWI) pathway operates through the sequential activation of protein kinases Bck1, Mkk1/2 and the effector MAPK Slt2 (outlined in Figure 4A). Interestingly, CWI pathway activation occurs in cells exposed to oxidative stress inducing agents, such as H2O2 [23]. In addition to Slt2, fungal resistance to a variety of stresses depends on the MAPK Hog1 [22]. Since ROS are increased in cells lacking Pmr1, we asked whether Slt2 or Hog1 are activated in pmr1Δ mutants. We therefore analyzed the levels of phosphorylated Slt2 and Hog1 in Wt and pmr1Δ mutant cells (Figure 4B). In contrast to Hog1, the levels of Slt2 and phosphorylated Slt2 were enhanced in the absence of Pmr1 indicating a constitutive activation of the Slt2 MAPK pathway in this strain. Thus, we addressed whether extracellular MnCl2 addition would be sufficient to stimulate Slt2 activation (Figure 4C). Wt and pmr1Δ mutants were grown for 1 h in the presence of increasing amounts of MnCl2 prior to protein extraction. Subsequent immunoblotting revealed a MnCl2 concentration-dependent increase in Slt2 and Hog1 phosphorylation. However, MnCl2 concentration-dependent phosphorylation was more evident for Hog1 and we did not observe a difference in Hog1 activation in pmr1Δ mutants as compared to Wt cells. To verify that the constitutive Slt2 phosphorylation observed in pmr1Δ mutants depends upon its upstream kinases Mkk1 and Mkk2, we compared MnCl2- and Congo red (CR)-induced Slt2 phosphorylation in cells devoid of these kinases (Figure 4D). Notably, the phosphorylated Slt2 signal was absent in mkk1Δ mkk2Δ (mkk1,2Δ) double as well as in pmr1Δ mkk1Δ mkk2Δ triple mutants excluding the possibility that MnCl2 could drive Slt2 phosphorylation by non-canonical kinases. Moreover, Hog1 phosphorylation was not affected by the concomitant absence of Mkk1 and Mkk2, as expected for Slt2-independent Hog1 phosphorylation in pmr1Δ mutants. These findings suggest that Slt2 phosphorylation is a hallmark of Pmr1 deficiency.
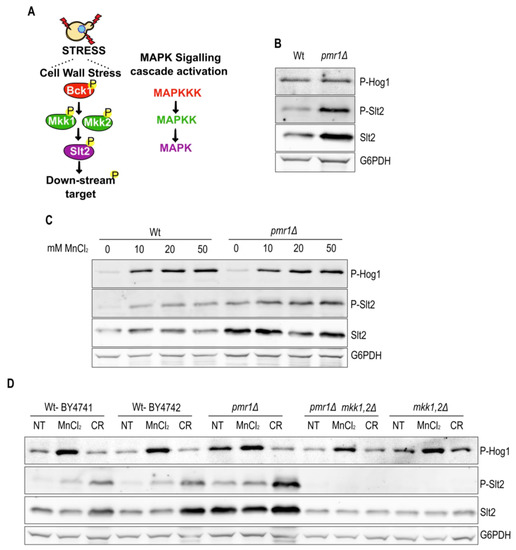
Figure 4.
The constitutive Slt2 phosphorylation is a consequence of Pmr1 depletion. (A) The schematic outline of the MAPK cell wall integrity signaling cascade in yeast. The relevant kinases are shown. (B) Hog1 and Slt2 phosphorylation of exponentially growing Wt and pmr1Δ mutant were analyzed by immunoblotting using phospho-specific antibodies (anti-P-p38 MAPK Thr180/182 and anti-p42/44 MAPK Erk1/2, respectively). The total Slt2 levels were detected with anti-Slt2 antibodies. Glucose-6-phosphate dehydrogenase (G6PDH) levels were used as a loading control. (C) Analysis of Hog1 and Slt2 phosphorylation levels in cells treated for 1 h with different MnCl2 concentration. Description as in B. (D) Comparison of MnCl2-induced Hog1 and Slt2 phosphorylation in cells lacking the Slt2 upstream kinases Mkk1/2. The non-treated cells (NT) were used as negative control. Congo red (CR) treatment was used as a control to selectively induce Slt2 phosphorylation. Description as in (B).
2.4. The Slt2 Target Rlm1 Is Up-Regulated in Cells Lacking Pmr1
SLT2 expression is known to be subjected to a feedback mechanism mediated by the CWI pathway specific transcription factor Rlm1 [30]. The observation that Slt2 protein levels are increased in pmr1Δ mutants (Figure 4) suggests that Rlm1 activates SLT2 gene expression. To test for this possibility, we generated pmr1Δ rlm1Δ double mutants and analyzed the levels of phosphorylated and total Slt2 (Figure 5A). The total Slt2 protein signal was reduced while phosphorylated Slt2 levels remained high in cells lacking Pmr1 and Rlm1. This finding is consistent with a model in which phosphorylated Slt2 activates Rlm1 to promote transcription of the SLT2 gene. We therefore took advantage of previously published microarray data [17], to figure out whether the expression of further Rlm1 target genes was increased in pmr1Δ mutants. This was indeed the case for a number of known Rlm1 target genes, including the SLT2 paralog KDX1 (MLP1), which was highly expressed in the absence of Pmr1 (Figure 5B).
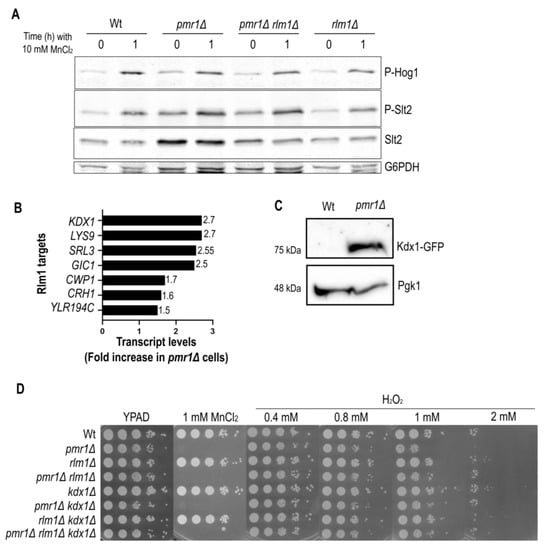
Figure 5.
Slt2-downstream targets are up-regulated in cells lacking Pmr1. (A) Exponentially growing Wt and indicated mutant strains were treated for up to 1 h with 10 mM MnCl2. The levels of Slt2, phosphorylated Slt2 (P-Slt2) and Hog1 (P-Hog1) were analyzed by immunoblotting as in Figure 4. Glucose-6-phosphate dehydrogenase (G6PDH) was used as a loading control. (B) Up-regulation of Rlm1 target gene transcripts in pmr1Δ mutants and (C) Kdx1-GFP protein level, analyzed by immunoblotting with anti-GFP antibodies, in Wt and pmr1Δ mutants. Pgk1 was used as loading control. (D) The growth of 10-fold dilutions of exponentially growing cells is shown. Strains, MnCl2 and H2O2 concentrations are indicated. The pictures were taken after 2 days of growth.
We performed a Western blotting analysis and confirmed that Kdx1 protein levels are highly increased in pmr1Δ mutants (Figure 5C). Kdx1 has been shown to interact with Rlm1 to drive the expression of stress responsive genes such as RCK1, which codes for a protein kinase involved in oxidative stress response [31]. It is noteworthy that Rck1 overexpression has been proposed to modulate Hog1 and Slt2 activation [32]. Next, we investigated if cells lacking Rlm1 and/or Kdx1 become more sensitive to H2O2 or MnCl2 (Figure 5D). As compared to Wt and pmr1Δ mutants, colony formation efficiency was similar in the presence or absence of Rlm1 and/or Kdx1, suggesting that these factors are dispensable for manganese and oxidative stress resistance.
2.5. Slt2 Contributes to Oxidative Stress Resistance of pmr1Δ Mutants
Having excluded a role for Rlm1 and Kdx1 in the oxidative stress resistance of pmr1Δ mutants, we strove to assess the impact of Slt2 itself on oxidative stress resistance. Yeast cells have been previously shown to become highly sensitive to H2O2 in the absence of Slt2 [23,33] and we therefore analyzed colony formation in the presence of H2O2 (Figure 6A). Notably, we did not observe a difference between the growth of Wt and slt2Δ mutants in H2O2 supplemented medium, probably because different strain backgrounds were used in our and previous studies. Importantly, pmr1Δ slt2Δ double mutants were much more sensitive to H2O2 than Wt, pmr1Δ or slt2Δ single mutant cells, providing evidence that Slt2 is required to withstand oxidative stress in the absence of Pmr1. The concomitant absence of Pmr1 and Yap1 resulted in further increased Slt2 phosphorylation compared to the pmr1Δ single mutant (Figure 6B). These results suggest that manganese stress signalling is channeled into the CWI pathway in the absence of Yap1 and led us to assess whether oxidative stress is responsible for Slt2 phosphorylation. To this end, cells were treated with the antioxidant N-acetyl cystein [4] to see if Slt2 phosphorylation can be diminished (Figure 6C). This was indeed the case, as N-acetyl cystein treatment strongly reduced Slt2 phosphorylation of pmr1Δ simple and pmr1Δ yap1Δ double mutants to similar levels. Since composite docking sites confer substrate recognition by both calcineurin and MAP kinases [34], we wondered if Slt2 regulates Yap1 protein levels upon MnCl2 exposure. Therefore, cells lacking Pmr1 and/or Slt2 were grown for 1 h in MnCl2 supplemented medium and Yap1-GFP signals analyzed by Western blotting (Supplementary Figure S1). However, MnCl2 leads to a similar drop in Yap1 protein levels in the absence of Pmr1 and/or Slt2 and, thus, these proteins are not involved in Yap1 stability or synthesis. Taken together, our results define a relevant role for Slt2 and Yap1 in the response to manganese stress and open the possibility of an additional mechanism involved in the regulation of Yap1 transcription, translation or protein stability.
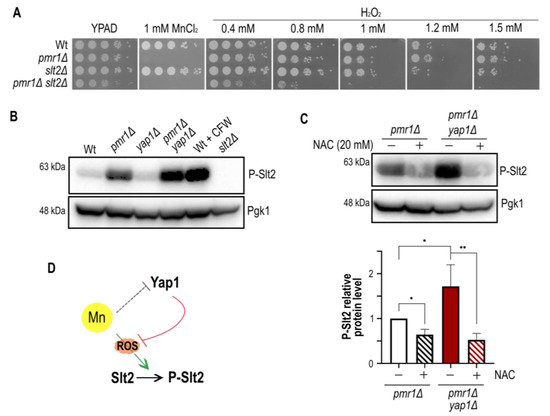
Figure 6.
Slt2 has a dual role in ROS and manganese stress defense. (A) The growth of 10-fold dilutions of exponentially growing cells is shown. Strains, MnCl2 and H2O2 concentrations are indicated. Pictures were taken after 2 days of growth. (B) Comparison of Slt2 phosphorylation levels in cells lacking Pmr1 and/or Yap1. Wt cells treated with calcofluor white (CFW) and slt2Δ mutants were used as positive and negative controls, respectively. (C) Comparison of Slt2 phosphorylation levels in N-acetyl cystein (NAC) treated cells. The quantification of Slt2 phosphorylation is shown (bottom). SD (with bars) and p-value are indicated (p-value 0.05 *; 0.01 **). The value obtained for untreated pmr1Δ mutants was set as 1. (D) Model for manganese stress-induced Slt2 activation and Yap1 protein decay, respectively. See text for details.
3. Discussion
3.1. Yap1 Has a Hitherto Unknown Role in Manganese Stress Response
Manganese is a redox-active metal that can mimic superoxide dismutases by catalyzing the decomposition of O2•− to H2O2 and O2 in vitro [35]. The same effect can be achieved in vivo as impaired manganese detoxification in cells devoid of Pmr1 can bypass the lack of Sod1 [6]. If intracellular manganese stimulates H2O2 formation, intracellular manganese overload upon Pmr1 depletion would provide an explanation for why mutant cells suffer from increased ROS formation. Our findings corroborate the previously observed increase in ROS levels in ATP2C1 depleted human keratinocytes [19]. We find that Yap1 protein levels are increased in cells lacking Pmr1. However, there are substantial differences between oxidative stress activation of the mammalian stress transcription factor Nrf2 as compared to yeast Yap1, because a complex stress-sensing system mechanism is needed for Yap1 nuclear retention including the assistance of glutathione peroxidase 3 (Gpx3) [27,36,37]. Regarding this unique activation mechanism, the Yap1 system might stem from a different evolutionary origin than the Keap1-Nrf2 system (reviewed in [38]).
To our knowledge, the impact of manganese stress on Yap1 activation has not been thoroughly explored. Interestingly, we find that Yap1 is needed for MnCl2 resistance, although MnCl2 addition leads to a rapid and dramatic drop in Yap1 protein levels (see Figure 2 and Figure 3). The nuclear accumulation of Yap1 upon oxidant challenge or due to impaired nuclear Yap1 export promotes its proteolytic degradation in a E3 Ubiquitin Ligase Not4-dependent manner [39]. MnCl2 may stimulate cytoplasmic to nuclear Yap1 turnover, but there is no evidence of a MnCl2-dependent change in Yap1 localization suggesting another mechanistic cause for MnCl2-driven Yap1 decay. It is possible that manganese interferes with Yap1 translation as MnCl2 was found to reduce total rRNA levels in a dose-dependent manner and to alter overall ribosome profiling [40], or one might even speculate that the 5′-UTR of YAP1 mRNA can form a riboswitch-like structure [41], whose interaction with manganese modulates YAP1 gene expression or mRNA stability. Yap1 has been shown to be regulated post-translationally in a calcineurin B-dependent manner [28], opening the possibility that MnCl2 could alter the enzymatic activity of calcineurin to promote Yap1 decay [42]. Calcineurin B-independent Yap1 down-regulation upon H2O2 [39] or MnCl2 addition suggests that manganese could drive the modulation of other phosphatase or kinase activities that promote Yap1 decay. Along this line, we recently found that manganese stimulates the enzymatic activity of the TORC1 complex in vitro and in vivo [43]. In any case, although calcineurin does not mediate MnCl2-triggered Yap1 decay this phosphatase is needed to activate the stress response transcription factor Crz1. Calcineurin-driven Crz1 dephosphorylation is required for nuclear Crz1 translocation, and impaired Crz1 activation in cna1Δ/cnb1Δ mutants renders these cells sensitive to manganese or arsenic stress [2,44,45]. Thus, it is likely that the balanced action of Yap1 and Crz1 activation is required for manganese stress tolerance.
3.2. SAPKs Activation Is a Read-Out for Manganese Stress
Deciphering the activation of different signaling pathways is important to define a manganese stress signature. The depletion of Pmr1 did not stimulate phosphorylation of the osmotic stress effector Hog1. In contrast, extracellular MnCl2 did activate Hog1 phosphorylation in a dose-dependent manner. Notably, MnCl2 addition results in a transient and similar induction of Hog1 phosphorylation in Wt and pmr1Δ mutants. Thus, Hog1 is required for an adaptive response to MnCl2 extracellular addition.
In addition to Hog1, extracellular MnCl2 supplementation promotes Slt2 phosphorylation in Wt and pmr1Δ mutants in a dose-dependent manner (see Figure 4). The total Slt2 levels and Slt2 phosphorylation were steadily increased in cells lacking Pmr1. The increase in Slt2 protein levels was dependent on the Slt2 downstream target Rlm1, a transcription factor that is part of a feedback loop to enhance SLT2 expression [30]. A hallmark for impaired cell wall integrity is the hypersensitivity to the cell wall damaging agents, a phenotype associated with the loss of Pmr1 function [46]. However, Slt2 activation could be a consequence of Golgi manganese depletion and consequential protein glycosylation and trafficking defects in the absence of Pmr1 [47].
Rlm1 has been shown to interact with the Slt2 paralog Kdx1 (Mlp1) to drive the expression of protein kinase Rck1 involved in oxidative stress response [31]. Indeed, we confirmed that Kdx1 protein levels were up-regulated in cells lacking Pmr1. However, genetic analysis failed to detect a growth defect if cells devoid of Pmr1 and/or Rml1/Kdx1 were grown in the presence of H2O2. Thus, it is unlikely that Rlm1/Kdx1 have a significant role in oxidative stress tolerance linked to manganese.
3.3. Activation of the CWI Stress Response in the Absence of Yap1
Corroborating previous observations that Slt2 protects cells from oxidative stress, a concomitant lack of Pmr1 and Slt2 renders cells hypersensitive to H2O2 treatment. Slt2 has been shown to promote transcription activation and the elongation of stress-induced genes by catalytic and non-catalytic mechanisms [48,49,50].
It is likely that in the absence of Yap1, the oxidative stress response is channeled into the CWI pathway. The molecular bases of Slt2 activation in the absence of Yap1 remains to be explored, but Slt2 activation may result from impaired Slt2 dephosphorylation, as has occurred with genotoxic stress [51]. It will be interesting to determine if reduced phosphatase activity is linked to enhanced Slt2 phosphorylation in cells lacking Pmr1, and if Slt2 indeed drives the transcriptional activation of oxidative stress-induced genes.
3.4. Concluding Remarks and Perspectives
Each kind of stress requires an adequate response to optimize cell survival. How stress signaling networks manage to crosstalk with each other is not well understood, but mechanistic evidence have been provided on how oxidative stress inhibits pheromone signaling [14]. Here, we report the results on the stress response signature of cells supplemented with extracellular manganese and/or lacking the Mn2+/Ca2+ Golgi transporter Pmr1. Thereby, we reveal the need for concomitant activation of various stress signaling pathways including Yap1 and SAPKs driven signaling as outlined in Figure 6D. A yet unanswered question is how stress signaling is channeled into the MAPK Slt2 in the absence of Yap1. In addition to the hitherto unknown role of Yap1 in manganese tolerance, we find that manganese induces a rapid reduction of Yap1 protein levels. However, the molecular bases of this manganese-driven Yap1 decay still remain to be explored in detail.
4. Materials and Methods
4.1. Yeast Strains, Plasmids, and Growth Conditions
Yeast strains and plasmids used in this study are listed in Table 1 and Table 2, respectively. Yeast transformants were grown in liquid or solid, adenine supplemented YPD medium (YPAD). Yeast mutants were generated by according to standard protocols by direct gene knock-out or N-terminal tagging.

Table 1.
Yeast strains used in this work.
Table 2.
Plasmids used in this work.
4.2. Drug Sensitivity Assays
Yeast cells were adjusted in concentration to an initial A600 of 0.5, and were then serially diluted 1:10 and spotted onto plates supplemented with MnCl2 (Sigma, St. Louis, MO, USA; 244589), or hydrogen peroxide (Alfa Aesar, Heysham, UK; L14000) at the indicated concentrations. The plates were incubated at 26 °C over the course of 2–3 days.
4.3. Yap1-GFP Localization
The cells were grown to exponential phase prior to fixation with 2.8% paraformaldehyde. Bright field images (DM-6000B, Leica, Wetzlar, Germany) were obtained at a 100× magnification, respectively. Fluorescence was detected using standard filters for mCherry (595 nm excitation/645–675 nm emission) and GFP (480 nm excitation/527 nm emission) and a digital charge-coupled device camera (DFC350, Leica) and pictures were processed with LAS AF (Leica).
4.4. Preparation of Yeast Extracts and Immunoblot Analysis
Yeast cells were grown overnight in the appropriate medium, diluted to A600 = 0.3 and grown for 3 h at 26 °C prior to addition of MnCl2, H2O2 or Cycloheximide (CHX). Protein extracts were isolated as previously described [43], separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS/PAGE), using 8% (w/v) polyacrylamide (37.5:1), and transferred to nitrocellulose membranes (Hybond, GE Healthcare, Amersham, UK) according to standard protocols. The Slt2 TEY phosphorylation site was marked a rabbit anti-phospho-p44/42 antibody (Thr-202/Tyr-204; Cell Signaling, Danvers, MA, USA, catalog# 9101), Hog1 phosphorylation with a p38 MAPK Thr180/182 antibody (Cell Signaling, catalog# 9101), Slt2 with an anti-Slt2 mouse antibody (Santa Cruz, Dallas, TX, USA, catalog# sc-374440), GFP with a anti-GFP mouse antibody (JL-8, Clontech, Shiga, Japan, catalog# 632381), G6DPH by an anti-G6DPH rabbit antibody (Sigma, catalog# A9521) and Pgk1 with an anti-Pgk1 mouse antibody (Invitrogen, Waltham, MA, USA, catalog# 459250) prior to inmuno-detection with a secondary antibody. Western blot images were acquired using a Chemidoc™ MP system and Image Lab software (Bio-Rad, Hercules, CA, USA). All immunoblots were reproduced at least twice in independent experiments with representative images shown.
4.5. Microarray Analysis
Microarray Analysis—Gene expression profiles were determined by using the “3′-expression microarray” technology by Affymetrix platform at the Genomics Unit of CABIMER (Seville, Spain) as described previously [5,17,18]. The microarray data can be derived from the GEO database using the identifier GSE29420.
4.6. ROS Detection Assay
For the detection of oxygen free radicals (ROS) and variations in the mitochondrial membrane potential or cell viability, yeast cells were grown on YPD and cultured as usual, and 2.5 µg/mL of dihydroethidium (DHE) or 5 µg/mL of propidium iodide (PI), respectively, were added to 1 mL of each sample and incubated at 24 °C for 5 and 30 min. Next, the samples were diluted 1:10 in PBS and analyzed on a FACScalibur flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA). The data were analyzed with FlowJo software (Becton Dickinson).
Supplementary Materials
The supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232415706/s1.
Author Contributions
Conceptualization, H.M., M.M. and R.E.W.; methodology and investigation, I.G.d.O., H.G. and E.J.-G.; writing—original draft preparation, H.M., M.M. and R.E.W.; writing—review and editing, I.G.d.O., H.G., E.J.-G., H.M., M.M. and R.E.W.; supervision, H.M., M.M. and R.E.W.; project administration, H.M., M.M. and R.E.W.; funding acquisition, H.M., M.M. and R.E.W. All authors have read and agreed to the published version of the manuscript.
Funding
R.E.W. research was funded by grants from the University of Seville (2020/00001326), Junta de Andalucía/European Union Regional Funds (P20-RT-01220), M.M. and H.M. research was funded by grant PID2019-105342GB-I00/AEI/10.13039/501100011033 from the Ministerio de Ciencia e Innovación (Spain). E.J.-G. was recipient of a predoctoral contract from Universidad Complutense de Madrid.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The row data of the experiments shown in this study are available from the corresponding author upon request. The microarray data is openly available in the GEO database (www.ncbi.nlm.nih.gov/geo) under accession number GSE29420.
Acknowledgments
We thank Elisabet Fernández-García, Néstor García-Rodríguez, Helle Ulrich and Woo-Hyun Chung for their generous supply with yeast plasmids and strains. We thank the Genomics Unit (Genomics and Proteomics Center, UCM) for sequencing reactions and the Flow Cytometry Unit (Cytometry and Fluorescence Microscopy Center, UCM).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Peres, T.V.; Schettinger, M.R.C.; Chen, P.; Carvalho, F.; Avila, D.S.; Bowman, A.B.; Aschner, M. Manganese-induced neurotoxicity: A review of its behavioral consequences and neuroprotective strategies. BMC Pharmacol. Toxicol. 2016, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.; Muend, S.; Zhang, Y.; Rao, R. A phenomics approach in yeast links proton and calcium pump function in the Golgi. Mol. Biol. Cell 2007, 18, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Kellermayer, R. Hailey–Hailey disease as an orthodisease of PMR1 deficiency in Saccharomyces cerevisiae. FEBS Lett. 2005, 579, 2021–2025. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.; Healy, E.; Strachan, T.; Sudbrak, R.; Perrussel, M.; Lehrach, H.; Munro, C.; Burge, S.; Hovnanian, A.; Monaco, A. Hailey-Hailey disease is caused by mutations in ATP2C1 encoding a novel Ca2+ pump. Hum. Mol. Genet. 2000, 9, 1131–1140. [Google Scholar]
- Ando, A.; Nakamura, T.; Murata, Y.; Takagi, H.; Shima, J. Identification and classification of genes required for tolerance to freeze–thaw stress revealed by genome-wide screening of Saccharomyces cerevisiae deletion strains. FEMS Yeast Res. 2007, 7, 244–253. [Google Scholar] [CrossRef]
- Lapinskas, P.J.; Cunningham, K.W.; Liu, X.F.; Fink, G.R.; Culotta, V.C. Mutations in PMR1 suppress oxidative damage in yeast cells lacking superoxide dismutase. Mol. Cell. Biol. 1995, 15, 1382–1388. [Google Scholar] [CrossRef]
- Nadal Clanchet, E.D.; Posas Garriga, F. Osmostress-induced gene expression--a model to understand how stress-activated protein kinases (SAPKs) regulate transcription. FEBS J. 2015, 282, 3275–3285. [Google Scholar] [CrossRef]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef]
- Duch, A.; De Nadal, E.; Posas, F. The p38 and Hog1 SAPKs control cell cycle progression in response to environmental stresses. FEBS Lett. 2012, 586, 2925–2931. [Google Scholar] [CrossRef]
- Yan, C.; Luo, H.; Lee, J.D.; Abe, J.; Berk, B.C. Molecular cloning of mouse ERK5/BMK1 splice variants and characterization of ERK5 functional domains. J. Biol. Chem. 2001, 276, 10870–10878. [Google Scholar] [CrossRef]
- Duch, A.; Felipe-Abrio, I.; Barroso, S.; Yaakov, G.; García-Rubio, M.; Aguilera, A.; de Nadal, E.; Posas, F. Coordinated control of replication and transcription by a SAPK protects genomic integrity. Nature 2013, 493, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Torres, M.; Jaquenoud, M.; De Virgilio, C. TORC1 controls G1–S cell cycle transition in yeast via Mpk1 and the greatwall kinase pathway. Nat. Commun. 2015, 6, 8256. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Pousada, C.; Devaux, F.; Caetano, S.M.; Pimentel, C.; da Silva, S.; Cordeiro, A.C.; Amaral, C. Yeast AP-1 like transcription factors (Yap) and stress response. Microb. Cell 2019, 6, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Jeong, S.-G.; Jin, S.H.; Mishra, R.; Peter, M.; Lee, C.-S.; Lee, S.S. Quantitative analysis of yeast MAPK signaling networks and crosstalk using a microfluidic device. Lab Chip 2020, 20, 2646–2655. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, K.-i.; Izawa, S.; Inoue, Y. The Yap1p-dependent induction of glutathione synthesis in heat shock response of Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 15535–15540. [Google Scholar] [CrossRef]
- Wu, A.-L.; Moye-Rowley, W.S. GSH1, which encodes gamma-glutamylcysteine synthetase, is a target gene for yAP-1 transcriptional regulation. Mol. Cell. Biol. 1994, 14, 5832–5839. [Google Scholar] [PubMed]
- Garcia-Rodriguez, N.; Diaz de la Loza, M.C.; Andreson, B.; Monje-Casas, F.; Rothstein, R.; Wellinger, R.E. Impaired manganese metabolism causes mitotic misregulation. J. Biol. Chem. 2012, 287, 18717–18729. [Google Scholar] [CrossRef]
- Bolton, E.C.; Mildvan, A.S.; Boeke, J.D. Inhibition of reverse transcription in vivo by elevated manganese ion concentration. Mol. Cell 2002, 9, 879–889. [Google Scholar] [CrossRef]
- Cialfi, S.; Le Pera, L.; De Blasio, C.; Mariano, G.; Palermo, R.; Zonfrilli, A.; Uccelletti, D.; Palleschi, C.; Biolcati, G.; Barbieri, L.; et al. The loss of ATP2C1 impairs the DNA damage response and induces altered skin homeostasis: Consequences for epidermal biology in Hailey-Hailey disease. Sci. Rep. 2016, 6, 31567. [Google Scholar] [CrossRef]
- Ficociello, G.; Zanni, E.; Cialfi, S.; Aurizi, C.; Biolcati, G.; Palleschi, C.; Talora, C.; Uccelletti, D. Glutathione S-transferase ϴ-subunit as a phenotypic suppressor of pmr1Δ strain, the Kluyveromyces lactis model for Hailey-Hailey disease. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 2650–2657. [Google Scholar] [CrossRef]
- Moye-Rowley, W.; Harshman, K.; Parker, C. Yeast YAP1 encodes a novel form of the jun family of transcriptional activator proteins. Genes Dev. 1989, 3, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Proft, M.; Mas, G.; de Nadal, E.; Vendrell, A.; Noriega, N.; Struhl, K.; Posas, F. The stress-activated Hog1 kinase is a selective transcriptional elongation factor for genes responding to osmotic stress. Mol. Cell 2006, 23, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Krasley, E.; Cooper, K.F.; Mallory, M.J.; Dunbrack, R.; Strich, R. Regulation of the oxidative stress response through Slt2p-dependent destruction of cyclin C in Saccharomyces cerevisiae. Genetics 2006, 172, 1477–1486. [Google Scholar] [CrossRef]
- Neklesa, T.K.; Davis, R.W. Superoxide anions regulate TORC1 and its ability to bind Fpr1: Rapamycin complex. Proc. Natl. Acad. Sci. USA 2008, 105, 15166–15171. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.T.; Epping, E.A.; Steggerda, S.M.; Moye-Rowley, W.S. Yap1p activates gene transcription in an oxidant-specific fashion. Mol. Cell. Biol. 1999, 19, 8302–8313. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rowe, L.A.; Degtyareva, N.; Doetsch, P.W. Yap1: A DNA damage responder in Saccharomyces cerevisiae. Mech. Ageing Dev. 2012, 133, 147–156. [Google Scholar] [CrossRef]
- Delaunay, A.; Isnard, A.-D.; Toledano, M.B. H2O2 sensing through oxidation of the Yap1 transcription factor. EMBO J. 2000, 19, 5157–5166. [Google Scholar] [CrossRef]
- Yokoyama, H.; Mizunuma, M.; Okamoto, M.; Yamamoto, J.; Hirata, D.; Miyakawa, T. Involvement of calcineurin-dependent degradation of Yap1p in Ca2+-induced G2 cell-cycle regulation in Saccharomyces cerevisiae. EMBO Rep. 2006, 7, 519–524. [Google Scholar] [CrossRef]
- Cyert, M.S.; Philpott, C.C. Regulation of cation balance in Saccharomyces cerevisiae. Genetics 2013, 193, 677–713. [Google Scholar] [CrossRef]
- García, R.; Sanz, A.B.; Rodríguez-Peña, J.M.; Nombela, C.; Arroyo, J. Rlm1 mediates positive autoregulatory transcriptional feedback that is essential for Slt2-dependent gene expression. J. Cell Sci. 2016, 129, 1649–1660. [Google Scholar] [CrossRef]
- Chang, M.; Kang, H.-J.; Baek, I.-J.; Kang, C.-M.; Park, Y.-S.; Yun, C.-W. Kdx1 regulates RCK1 gene expression by interacting with Rlm1 in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2013, 435, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Kang, H.-J.; Baek, I.-J.; Kang, C.-M.; Park, Y.-S.; Yun, C.-W. Rck1 up-regulates Hog1 activity by down-regulating Slt2 activity in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2013, 440, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Pujol, N.; Petkova, M.I.; Serrano, L.; Torre-Ruiz, M.A.D.L. The MAP kinase Slt2 is involved in vacuolar function and actin remodeling in Saccharomyces cerevisiae mutants affected by endogenous oxidative stress. Appl. Environ. Microbiol. 2013, 79, 6459–6471. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.; Roy, J.; Bodenmiller, B.; Wanka, S.; Landry, C.R.; Aebersold, R.; Cyert, M.S. The calcineurin signaling network evolves via conserved kinase-phosphatase modules that transcend substrate identity. Mol. Cell 2014, 55, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.P.; Lennon, P.J.; Neumann, W.L.; Weiss, R.H. Toward the rational design of superoxide dismutase mimics: Mechanistic studies for the elucidation of substituent effects on the catalytic activity of macrocyclic manganese (II) complexes. J. Am. Chem. Soc. 1997, 119, 6522–6528. [Google Scholar] [CrossRef]
- Delaunay, A.; Pflieger, D.; Barrault, M.-B.; Vinh, J.; Toledano, M.B. A thiol peroxidase is an H2O2 receptor and redox-transducer in gene activation. Cell 2002, 111, 471–481. [Google Scholar] [CrossRef]
- Okazaki, S.; Tachibana, T.; Naganuma, A.; Mano, N.; Kuge, S. Multistep disulfide bond formation in Yap1 is required for sensing and transduction of H2O2 stress signal. Mol. Cell 2007, 27, 675–688. [Google Scholar] [CrossRef]
- Fuse, Y.; Kobayashi, M. Conservation of the Keap1-Nrf2 system: An evolutionary journey through stressful space and time. Molecules 2017, 22, 436. [Google Scholar] [CrossRef]
- Gulshan, K.; Thommandru, B.; Moye-Rowley, W.S. Proteolytic degradation of the Yap1 transcription factor is regulated by subcellular localization and the E3 ubiquitin ligase Not4. J. Biol. Chem. 2012, 287, 26796–26805. [Google Scholar] [CrossRef]
- Luo, X.; Li, S.; Lu, L.; Liu, B.; Kuang, X.; Shao, G.; Yu, S. Gene expression of manganese-containing superoxide dismutase as a biomarker of manganese bioavailability for manganese sources in broilers. Poult. Sci. 2007, 86, 888–894. [Google Scholar] [CrossRef]
- Vilela, C.; Linz, B.; Rodrigues-Pousada, C.; McCarthy, J.E. The yeast transcription factor genes YAP1 and YAP2 are subject to differential control at the levels of both translation and mRNA stability. Nucleic Acids Res. 1998, 26, 1150–1159. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Martin, B.L.; Jurado, L.A.; Hengge, A.C. Comparison of the reaction progress of calcineurin with Mn2+ and Mg2+. Biochemistry 1999, 38, 3386–3392. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, R.; Gaillard, H.; Zarzuela, L.; Péli-Gulli, M.-P.; Fernández-García, E.; Tomé, M.; García-Rodríguez, N.; Durán, R.V.; De Virgilio, C.; Wellinger, R.E. Manganese is a physiologically relevant TORC1 activator in yeast and mammals. elife 2022, 11, e80497. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Li, H.; Hogan, P.G.; Cyert, M.S. A conserved docking site modulates substrate affinity for calcineurin, signaling output, and in vivo function. Mol. Cell 2007, 25, 889–901. [Google Scholar] [CrossRef][Green Version]
- Ferreira, R.T.; Silva, A.R.C.; Pimentel, C.; Batista-Nascimento, L.; Rodrigues-Pousada, C.; Menezes, R.A. Arsenic stress elicits cytosolic Ca2+ bursts and Crz1 activation in Saccharomyces cerevisiae. Microbiology 2012, 158, 2293–2302. [Google Scholar] [CrossRef]
- García-Rodríguez, N.; Manzano-López, J.; Muñoz-Bravo, M.; Fernández-García, E.; Muñiz, M.; Wellinger, R.E. Manganese redistribution by calcium-stimulated vesicle trafficking bypasses the need for P-type ATPase function. J. Biol. Chem. 2015, 290, 9335–9347. [Google Scholar] [CrossRef]
- Durr, G.; Strayle, J.; Plemper, R.; Elbs, S.; Klee, S.K.; Catty, P.; Wolf, D.H.; Rudolph, H.K. The medial-Golgi ion pump Pmr1 supplies the yeast secretory pathway with Ca2+ and Mn2+ required for glycosylation, sorting, and endoplasmic reticulum-associated protein degradation. Mol. Biol. Cell 1998, 9, 1149–1162. [Google Scholar] [CrossRef]
- Kim, K.-Y.; Truman, A.W.; Levin, D.E. Yeast Mpk1 MAPK activates transcription through Swi4/Swi6 by a non-catalytic mechanism that requires upstream signal. Mol. Cell. Biol. 2008, 28, 2579–2589. [Google Scholar] [CrossRef]
- Kim, K.-Y.; Levin, D.E. Mpk1 MAPK association with the Paf1 complex blocks Sen1-mediated premature transcription termination. Cell 2011, 144, 745–756. [Google Scholar] [CrossRef]
- Yurko, N.; Liu, X.; Yamazaki, T.; Hoque, M.; Tian, B.; Manley, J.L. MPK1/SLT2 links multiple stress responses with gene expression in budding yeast by phosphorylating Tyr1 of the RNAP II CTD. Mol. Cell 2017, 68, 913–925.e3. [Google Scholar] [CrossRef]
- Liu, L.; Levin, D.E. Intracellular mechanism by which genotoxic stress activates yeast SAPK Mpk1. Mol. Biol. Cell 2018, 29, 2898–2909. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).