


Investigation of the Interaction between Aloe vera Anthraquinone Metabolites and c-Myc and C-Kit G-Quadruplex DNA Structures

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
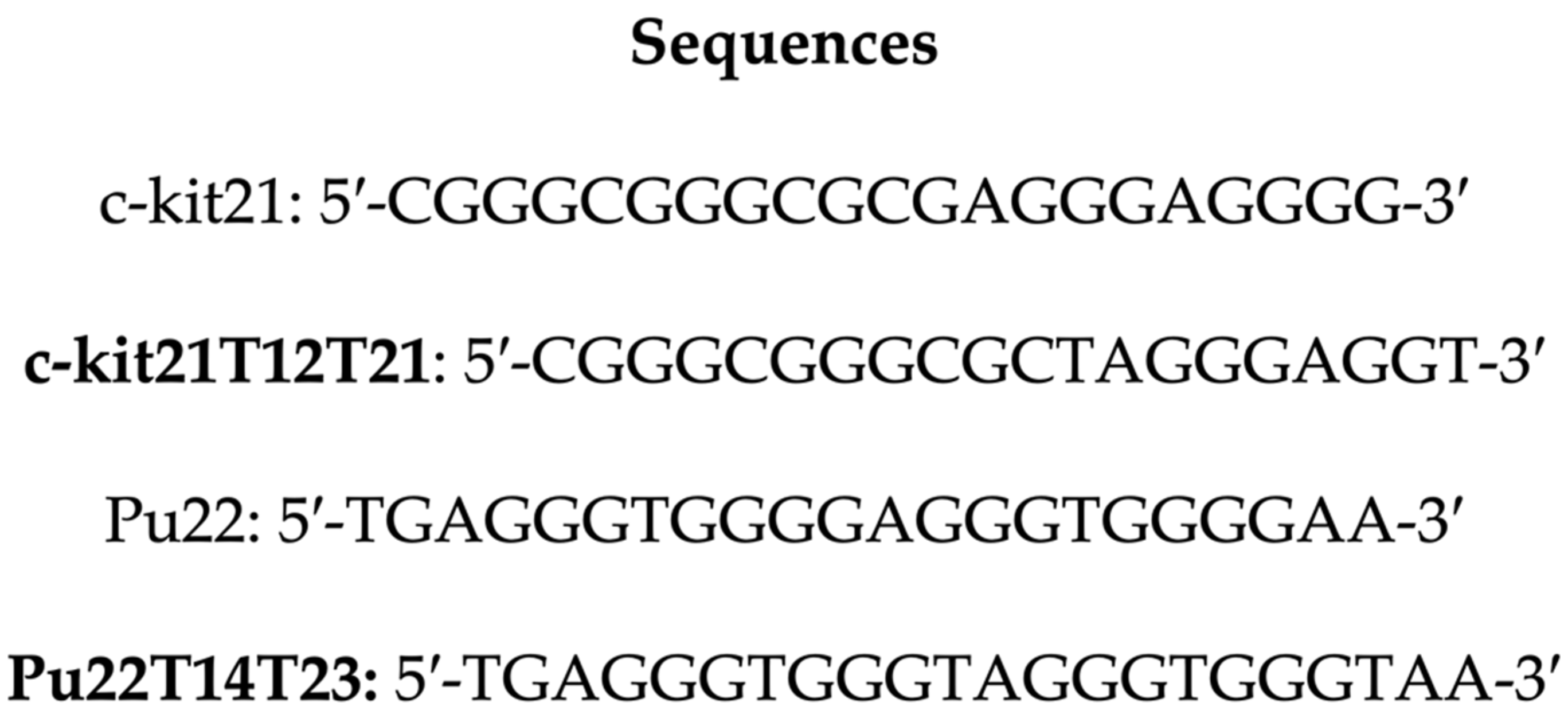
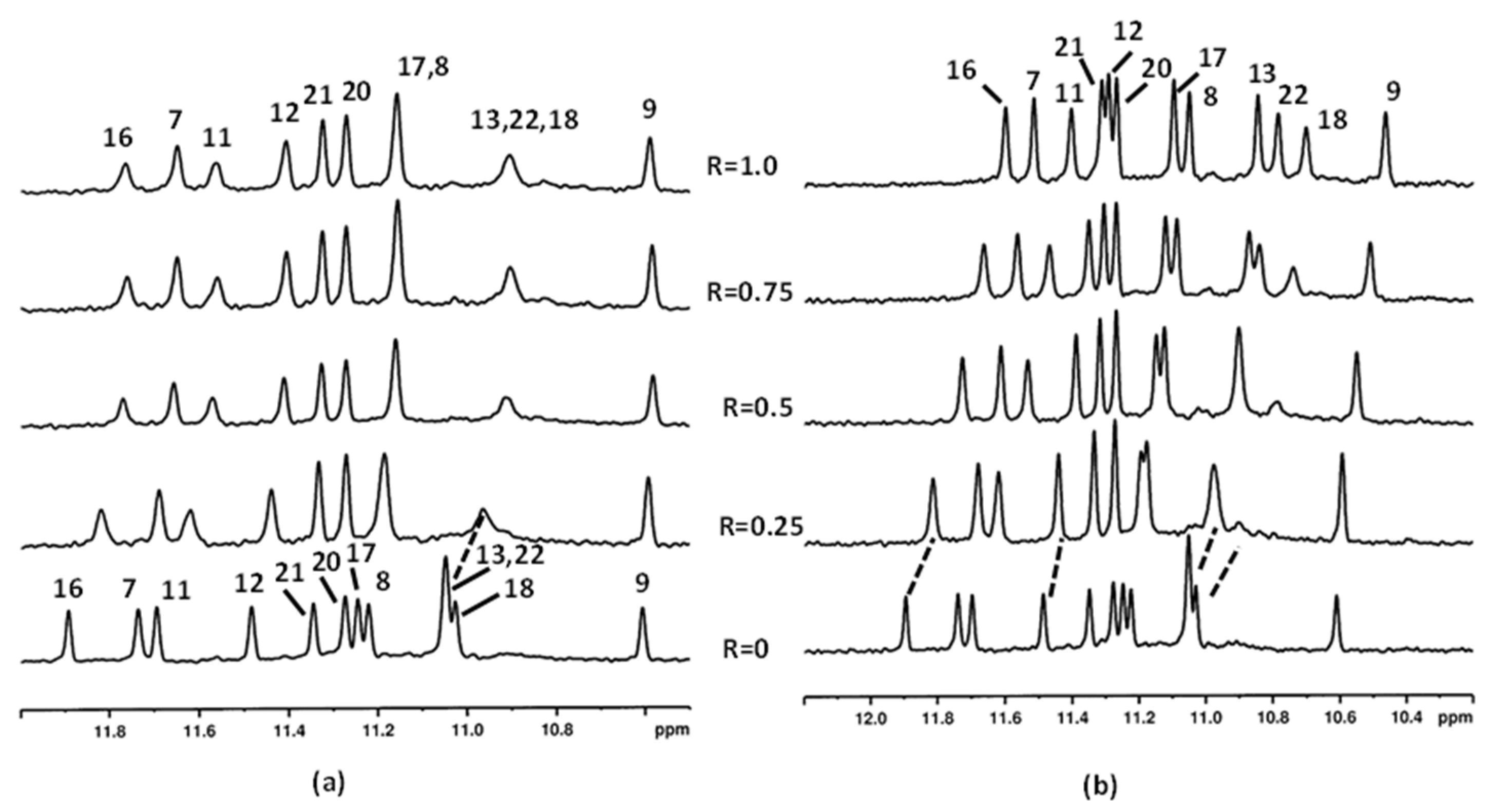
2.1. Interaction of Aloe Anthraquinones with c-kit21T12T21 and Pu22T14T23
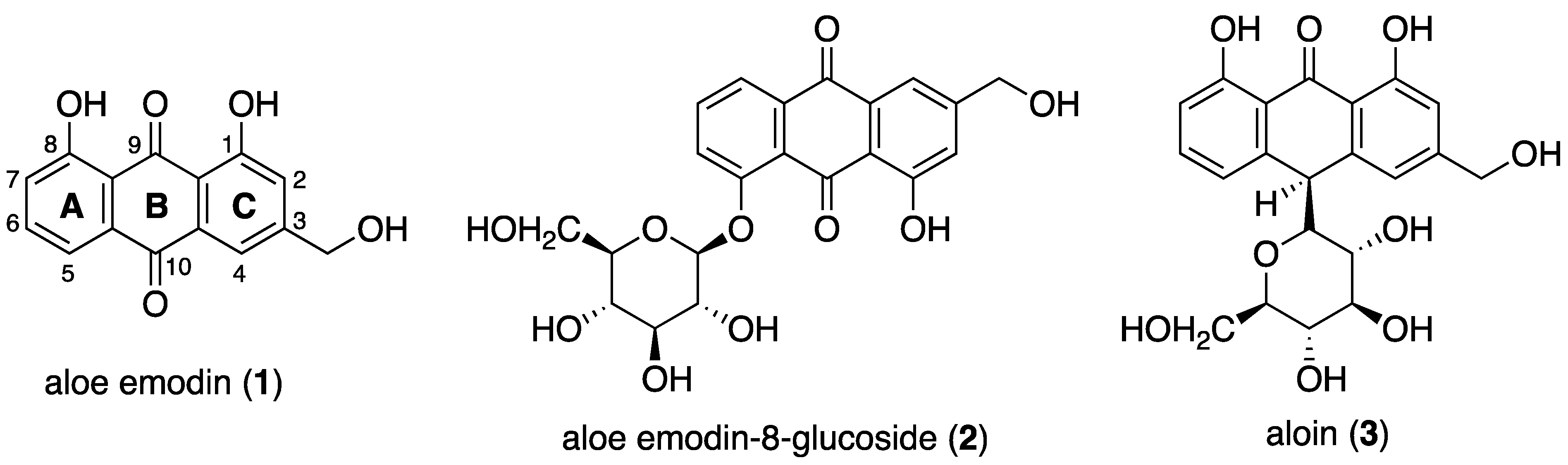
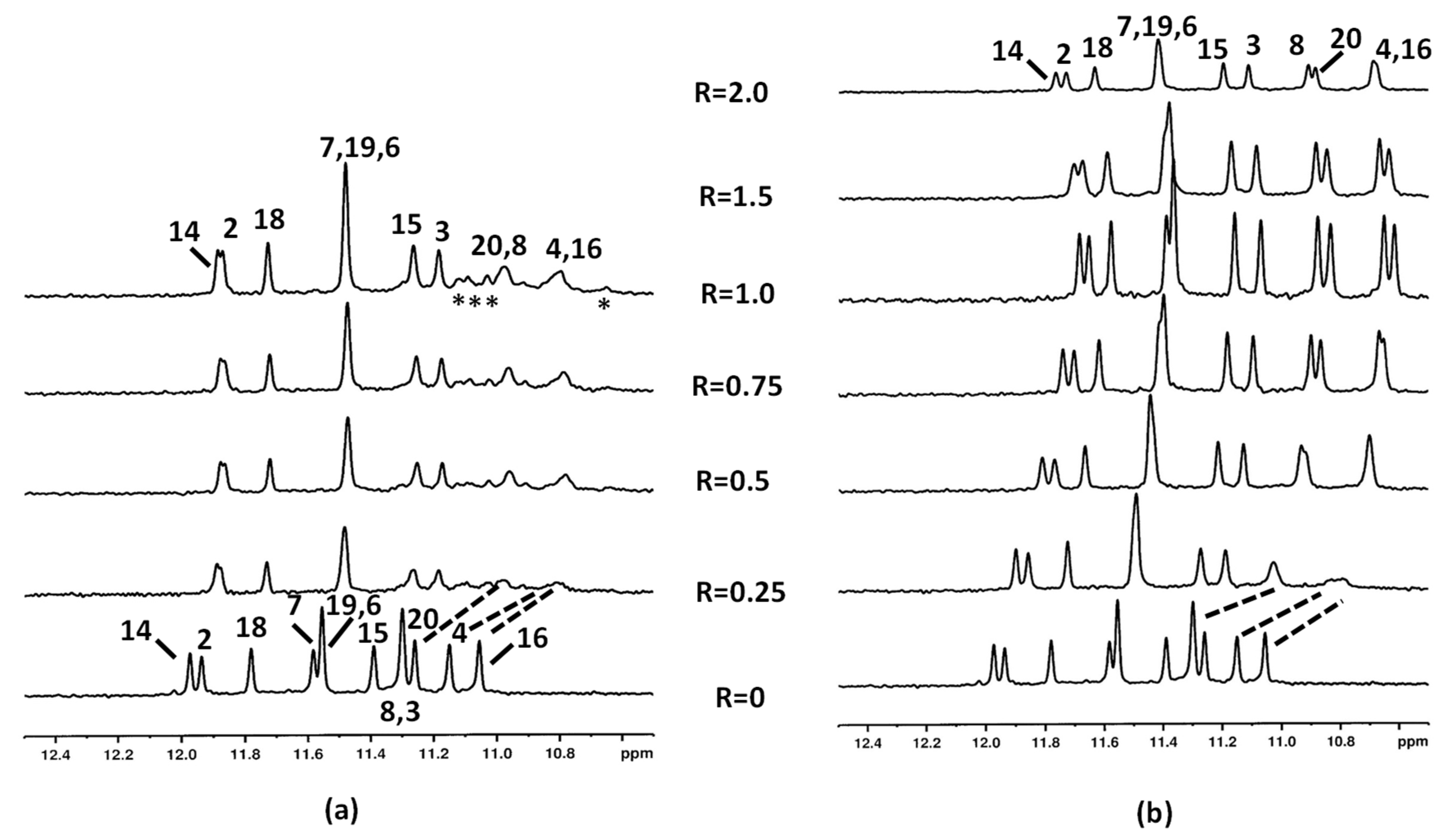
2.1.1. Interaction of Aloe Emodin (1) and Aloe Emodin-8-glucoside (2) with c-kit21T12T21
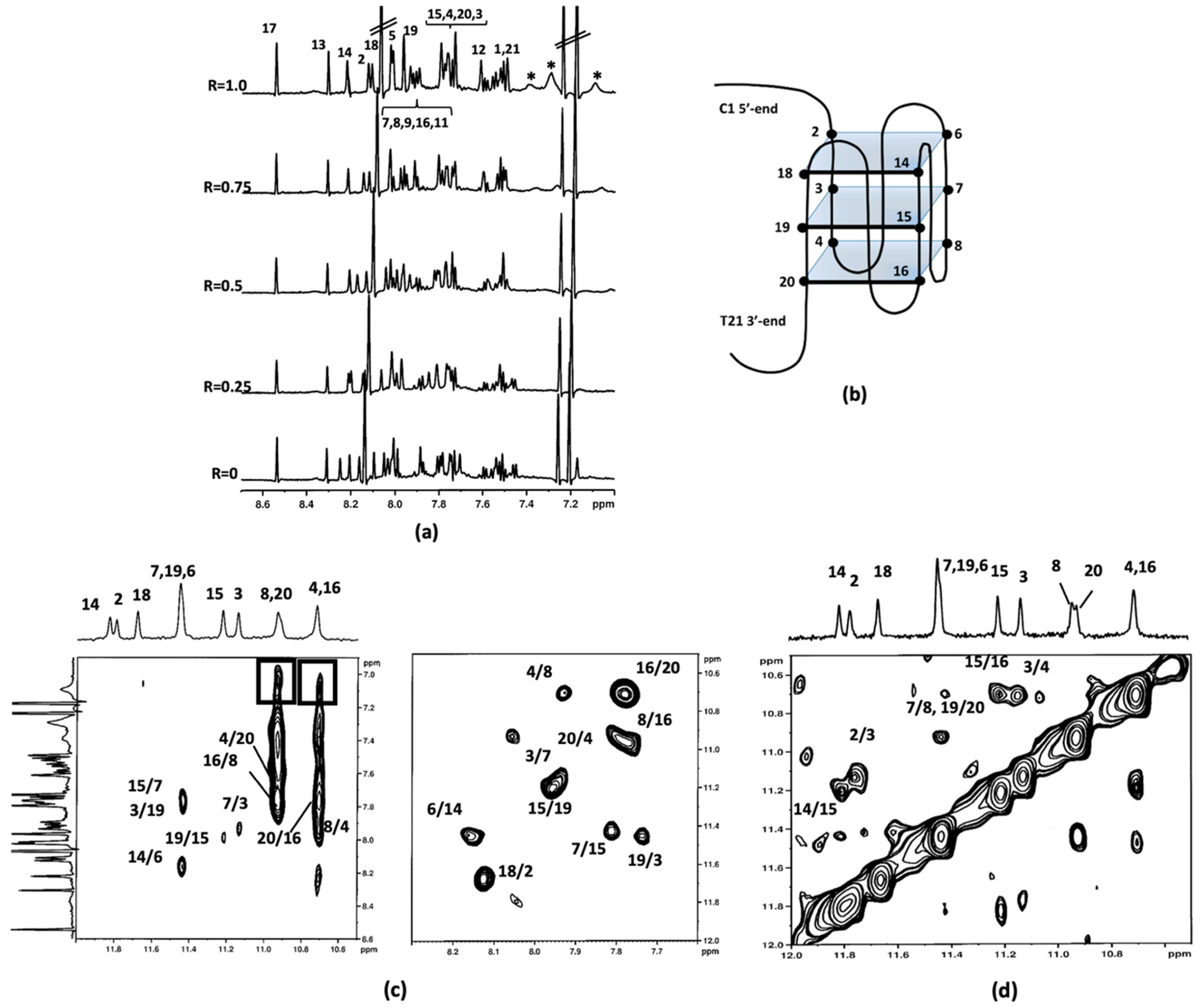
2.1.2. Interaction of Compounds 1–3 with Pu22T14T23
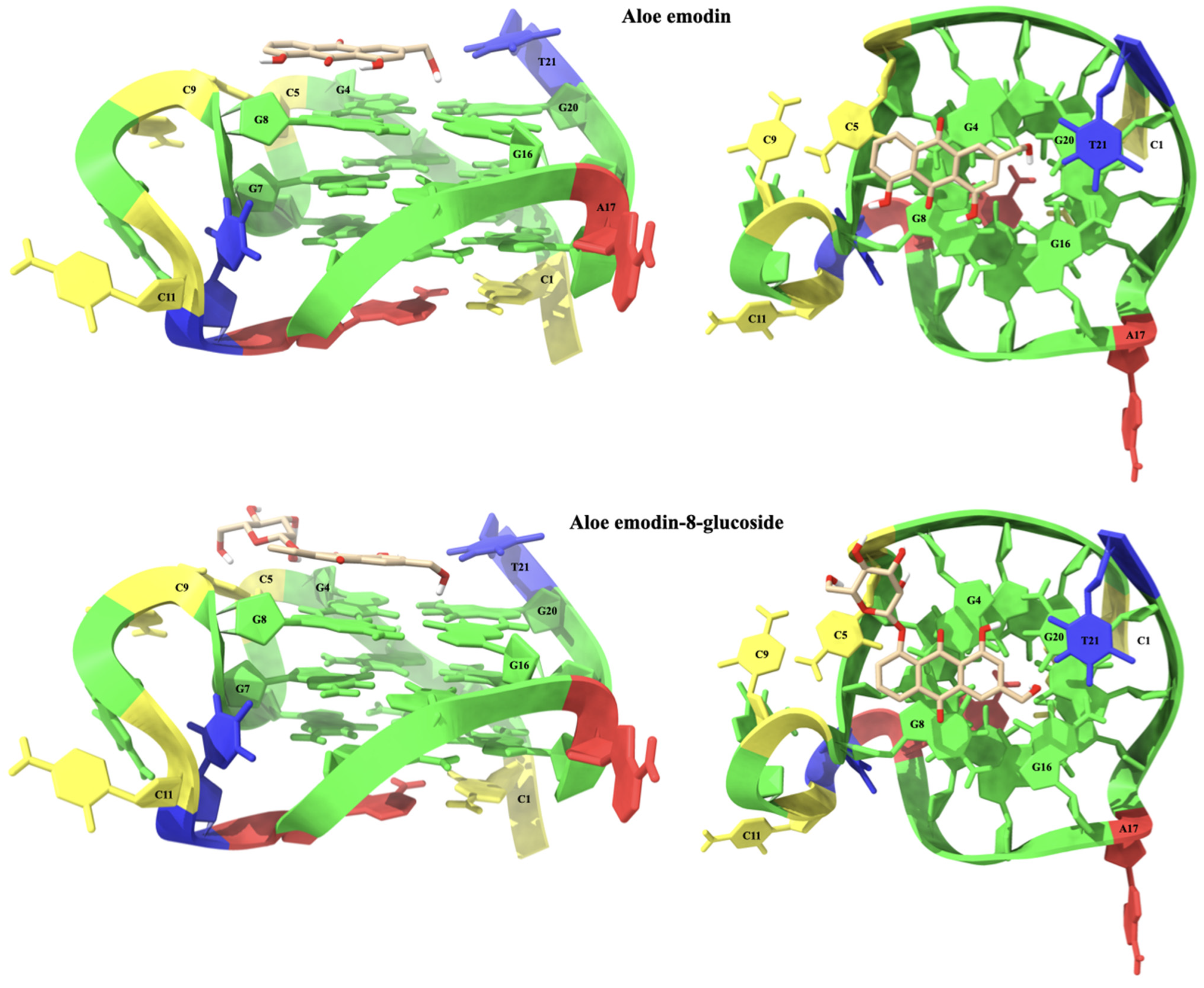
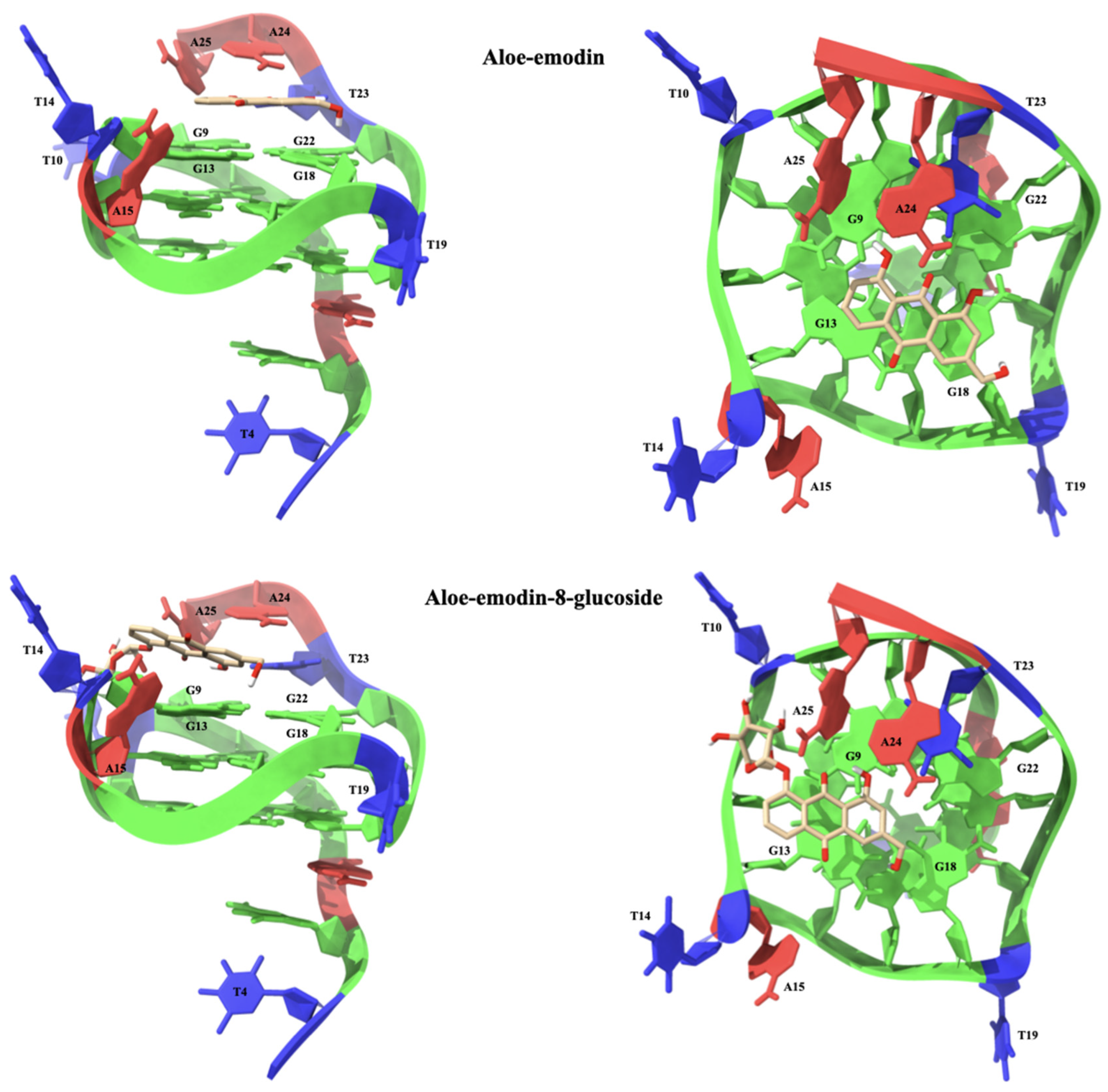
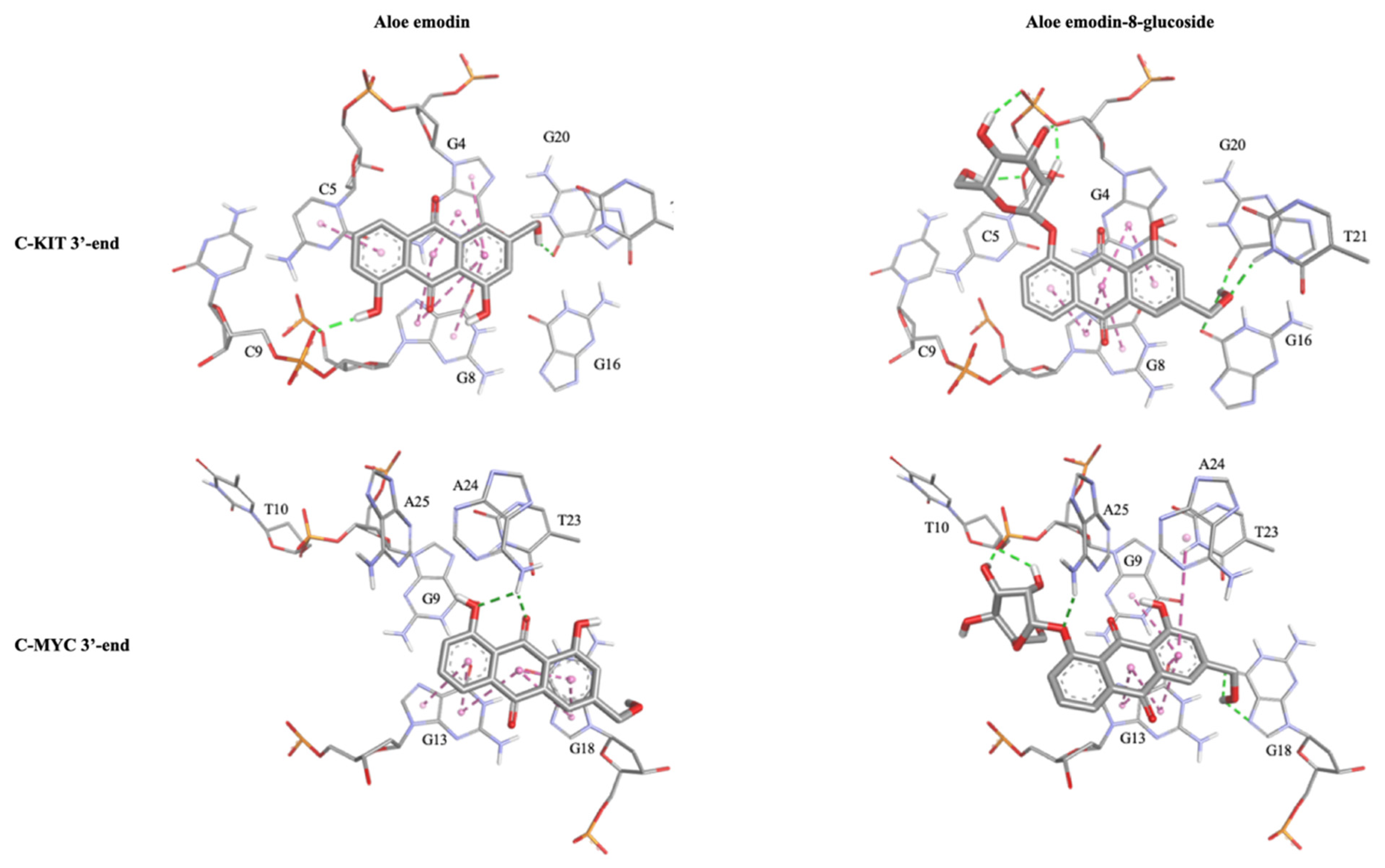
2.1.3. Molecular Modeling Studies
3. Materials and Methods
3.1. Ligands
3.2. NMR Experiments
3.3. Molecular Modeling Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chien, S.-C.; Wu, Y.-C.; Chen, Z.-W.; Yang, W.-C. Naturally Occurring Anthraquinones: Chemistry and Therapeutic Potential in Autoimmune Diabetes. Evid.-Based Complement. Altern. Med. 2015, 2015, 357357. [Google Scholar] [CrossRef] [PubMed]
- Berillo, D.; Kozhahmetova, M.; Lebedeva, L. Overview of the Biological Activity of Anthraquinons and Flavanoids of the Plant Rumex Species. Molecules 2022, 27, 1204. [Google Scholar] [CrossRef] [PubMed]
- Sebak, M.; Molham, F.; Greco, C.; Tammam, M.A.; Sobeh, M.; El-Demerdash, A. Chemical Diversity, Medicinal Potentialities, Biosynthesis, and Pharmacokinetics of Anthraquinones and Their Congeners Derived from Marine Fungi: A Comprehensive Update. RSC Adv. 2022, 12, 24887–24921. [Google Scholar] [CrossRef] [PubMed]
- Seigler, D.S. Plant Secondary Metabolism; Springer: Boston, MA, USA, 1998; ISBN 978-1-4613-7228-8. [Google Scholar]
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: Molecular Advances and Pharmacologic Developments in Antitumor Activity and Cardiotoxicity. Pharm. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, A.; Mahar, A.S. DNA targeted anthraquinone derivatives: An important anticancer agents. Int. J. Pharm. Pharm. Sci. 2016, 8, 17–25. [Google Scholar]
- Das, A.; Suresh Kumar, G.; Dutta, S. Interaction of Aloe Active Compounds with Calf Thymus DNA. J. Mol. Recognit. 2019, 32, e2786. [Google Scholar] [CrossRef]
- Huang, P.-H.; Huang, C.-Y.; Chen, M.-C.; Lee, Y.-T.; Yue, C.-H.; Wang, H.-Y.; Lin, H. Emodin and Aloe-Emodin Suppress Breast Cancer Cell Proliferation through ER α Inhibition. Evid.-Based Complement. Altern. Med. 2013, 2013, 376123. [Google Scholar] [CrossRef] [Green Version]
- Ahirwar, K.; Jain, S. Aloe-Emodin Novel Anticancer Herbal Drug. Int. J. Phytomed. 2011, 3, 27–31. [Google Scholar]
- Özenver, N.; Saeed, M.; Ömür Demirezer, L.; Efferth, T. Aloe-Emodin as Drug Candidate for Cancer Therapy. Oncotarget 2018, 9, 17770–17796. [Google Scholar] [CrossRef] [Green Version]
- Pecere, T.; Gazzola, M.V.; Mucignat, C.; Parolin, C.; Vecchia, F.D.; Cavaggioni, A.; Basso, G.; Diaspro, A.; Salvato, B.; Carli, M.; et al. Aloe-Emodin Is a New Type of Anticancer Agent with Selective Activity against Neuroectodermal Tumors1. Cancer Res. 2000, 60, 2800–2804. [Google Scholar]
- Anand, S.; Muthusamy, V.S.; Sujatha, S.; Sangeetha, K.N.; Bharathi Raja, R.; Sudhagar, S.; Poornima Devi, N.; Lakshmi, B.S. Aloe Emodin Glycosides Stimulates Glucose Transport and Glycogen Storage through PI3K Dependent Mechanism in L6 Myotubes and Inhibits Adipocyte Differentiation in 3T3L1 Adipocytes. FEBS Lett. 2010, 584, 3170–3178. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Pan, H.; Lou, H.; Xu, Y.; Tian, L. Inhibition of the Angiogenesis and Growth of Aloin in Human Colorectal Cancer in Vitro and in Vivo. Cancer Cell Int. 2013, 13, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmat, A.Y.; Said, M.M.; Hamdy, G.M.; Soliman, A.A.; Khalil, S.A. In Vivo and In Vitro Studies on the Antioxidant Activity of Aloin Compared to Doxorubicin in Rats. Drug Dev. Res 2012, 73, 154–165. [Google Scholar] [CrossRef]
- Niciforovic, A.; Adzic, M.; Spasic, S.D.; Radojcic, M.B. Antitumor Effects of a Natural Anthracycline Analog (Aloin) Involve Altered Activity of Antioxidant Enzymes in HeLaS3 Cells. Cancer Biol. 2007, 6, 1211–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallavalle, S.; Musso, L.; Artali, R.; Aviñó, A.; Scaglioni, L.; Eritja, R.; Gargallo, R.; Mazzini, S. G-Quadruplex Binding Properties of a Potent PARP-1 Inhibitor Derived from 7-Azaindole-1-Carboxamide. Sci. Rep. 2021, 11, 3869. [Google Scholar] [CrossRef] [PubMed]
- Dallavalle, S.; Princiotto, S.; Mattio, L.M.; Artali, R.; Musso, L.; Aviñó, A.; Eritja, R.; Pisano, C.; Gargallo, R.; Mazzini, S. Investigation of the Complexes Formed between PARP1 Inhibitors and PARP1 G-Quadruplex at the Gene Promoter Region. Int. J. Mol. Sci. 2021, 22, 8737. [Google Scholar] [CrossRef]
- Dallavalle, S.; Mattio, L.M.; Artali, R.; Musso, L.; Aviñó, A.; Fàbrega, C.; Eritja, R.; Gargallo, R.; Mazzini, S. Exploring the Interaction of Curaxin CBL0137 with G-Quadruplex DNA Oligomers. Int. J. Mol. Sci. 2021, 22, 6476. [Google Scholar] [CrossRef]
- Platella, C.; Mazzini, S.; Napolitano, E.; Mattio, L.M.; Beretta, G.L.; Zaffaroni, N.; Pinto, A.; Montesarchio, D.; Dallavalle, S. Plant-Derived Stilbenoids as DNA-Binding Agents: From Monomers to Dimers. Chem. Eur. J. 2021, 27, 8832–8845. [Google Scholar] [CrossRef]
- Mazzini, S.; Gargallo, R.; Musso, L.; de Santis, F.; Aviñó, A.; Scaglioni, L.; Eritja, R.; di Nicola, M.; Zunino, F.; Amatulli, A.; et al. Stabilization of C-KIT G-Quadruplex DNA Structures by the RNA Polymerase I Inhibitors BMH-21 and BA-41. Int. J. Mol. Sci. 2019, 20, 4927. [Google Scholar] [CrossRef] [Green Version]
- Musso, L.; Mazzini, S.; Rossini, A.; Castagnoli, L.; Scaglioni, L.; Artali, R.; di Nicola, M.; Zunino, F.; Dallavalle, S. C-MYC G-Quadruplex Binding by the RNA Polymerase I Inhibitor BMH-21 and Analogues Revealed by a Combined NMR and Biochemical Approach. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 615–629. [Google Scholar] [CrossRef]
- Lipps, H.J.; Rhodes, D. G-Quadruplex Structures: In Vivo Evidence and Function. Trends Cell Biol. 2009, 19, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Ganguly, A.; Chowdhury, S. Promise of G-Quadruplex Structure Binding Ligands as Epigenetic Modifiers with Anti-Cancer Effects. Molecules 2019, 24, 582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Dutta, S. Binding Studies of Aloe-Active Compounds with G-Quadruplex Sequences. ACS Omega 2021, 6, 18344–18351. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yan, W.-W.; He, M.; Wei, D.; Long, Z.-J.; Tao, Y.-M. Aloe Emodin Inhibits Telomerase Activity in Breast Cancer Cells: Transcriptional and Enzymological Mechanism. Pharmacol. Rep. 2020, 72, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Pagano, B.; Amato, J.; Iaccarino, N.; Cingolani, C.; Zizza, P.; Biroccio, A.; Novellino, E.; Randazzo, A. Looking for Efficient G-Quadruplex Ligands: Evidence for Selective Stabilizing Properties and Telomere Damage by Drug-Like Molecules. ChemMedChem 2015, 10, 640–649. [Google Scholar] [CrossRef]
- Jarosova, P.; Paroulek, P.; Rajecky, M.; Rajecka, V.; Taborska, E.; Eritja, R.; Aviñó, A.; Mazzini, S.; Gargallo, R.; Taborsky, P. Naturally Occurring Quaternary Benzo[c]Phenanthridine Alkaloids Selectively Stabilize G-Quadruplexes. Phys. Chem. Chem. Phys. 2018, 20, 21772–21782. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Carver, M.; Hurley, L.H.; Yang, D. Solution Structure of a 2:1 Quindoline–c-MYC G-Quadruplex: Insights into G-Quadruplex-Interactive Small Molecule Drug Design. J. Am. Chem. Soc. 2011, 133, 17673–17680. [Google Scholar] [CrossRef] [Green Version]
- Fernando, H.; Reszka, A.P.; Huppert, J.; Ladame, S.; Rankin, S.; Venkitaraman, A.R.; Neidle, S.; Balasubramanian, S. A Conserved Quadruplex Motif Located in a Transcription Activation Site of the Human C-Kit Oncogene. Biochemistry 2006, 45, 7854–7860. [Google Scholar] [CrossRef] [Green Version]
- Kuryavyi, V.; Phan, A.T.; Patel, D.J. Solution Structures of All Parallel-Stranded Monomeric and Dimeric G-Quadruplex Scaffolds of the Human c-Kit2 Promoter. Nucleic Acids Res. 2010, 38, 6757–6773. [Google Scholar] [CrossRef] [Green Version]
- Seenisamy, J.; Rezler, E.M.; Powell, T.J.; Tye, D.; Gokhale, V.; Joshi, C.S.; Siddiqui-Jain, A.; Hurley, L.H. The Dynamic Character of the G-Quadruplex Element in the c-MYC Promoter and Modification by TMPyP4. J. Am. Chem. Soc. 2004, 126, 8702–8709. [Google Scholar] [CrossRef]
- Ambrus, A.; Chen, D.; Dai, J.; Jones, R.A.; Yang, D. Solution Structure of the Biologically Relevant G-Quadruplex Element in the Human c-MYC Promoter. Implications for G-Quadruplex Stabilization. Biochemistry 2005, 44, 2048–2058. [Google Scholar] [CrossRef] [PubMed]
- Hay, J.E.; Haynes, L.J. 605. The Aloins. Part I. The Structure of Barbaloin. J. Chem. Soc. 1956, 3141–3147. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated Docking Using a Lamarckian Genetic Algorithm and an Empirical Binding Free Energy Function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A Programming Language for Software Integration and Development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Gasteiger, J.; Marsili, M. Iterative Partial Equalization of Orbital Electronegativity—A Rapid Access to Atomic Charges. Tetrahedron 1980, 36, 3219–3228. [Google Scholar] [CrossRef]
- Harrach, M.F.; Drossel, B. Structure and Dynamics of TIP3P, TIP4P, and TIP5P Water near Smooth and Atomistic Walls of Different Hydroaffinity. J. Chem. Phys. 2014, 140, 174501. [Google Scholar] [CrossRef]
- Galindo-Murillo, R.; Robertson, J.C.; Zgarbová, M.; Šponer, J.; Otyepka, M.; Jurečka, P.; Cheatham, T.E. Assessing the Current State of Amber Force Field Modifications for DNA. J. Chem. Theory Comput. 2016, 12, 4114–4127. [Google Scholar] [CrossRef]
- Ivani, I.; Dans, P.D.; Noy, A.; Pérez, A.; Faustino, I.; Hospital, A.; Walther, J.; Andrio, P.; Goñi, R.; Balaceanu, A.; et al. Parmbsc1: A Refined Force Field for DNA Simulations. Nat. Methods 2016, 13, 55–58. [Google Scholar] [CrossRef] [Green Version]
- Lamoureux, G.; Roux, B. Modeling Induced Polarization with Classical Drude Oscillators: Theory and Molecular Dynamics Simulation Algorithm. J. Chem. Phys. 2003, 119, 3025–3039. [Google Scholar] [CrossRef]
- Jiang, W.; Hardy, D.J.; Phillips, J.C.; MacKerell, A.D.; Schulten, K.; Roux, B. High-Performance Scalable Molecular Dynamics Simulations of a Polarizable Force Field Based on Classical Drude Oscillators in NAMD. J. Phys. Chem. Lett. 2011, 2, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable Molecular Dynamics on CPU and GPU Architectures with NAMD. J. Chem. Phys. 2020, 153, 44130. [Google Scholar] [CrossRef] [PubMed]
- Ryckaert, J.-P.; Ciccotti, G.; Berendsen, H.J.C. Numerical Integration of the Cartesian Equations of Motion of a System with Constraints: Molecular Dynamics of n-Alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, S.; Kollman, P.A. Settle: An Analytical Version of the SHAKE and RATTLE Algorithm for Rigid Water Models. J. Comput. Chem. 1992, 13, 952–962. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle Mesh Ewald: An N⋅log(N) Method for Ewald Sums in Large Systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure Visualization for Researchers, Educators, and Developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting Modern Challenges in Visualization and Analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligands | C-Kit 3′-End Tetrad | c-Myc 3′-End Tetrad | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G4 | C5 | G8 | C9 | G16 | G20 | G21 | G9 | T10 | G13 | G18 | A24 | A25 | |
| aloe emodin (1) | π-π | π-π | π-π | HB | HB | π-π | π-π | HB | |||||
| aloe emodin-8-glucoside (2) | HB, π-π | HB | π-π | HB | HB | HB | π-π | HB | π-π | HB | π-π | HB | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dallavalle, S.; Artali, R.; Princiotto, S.; Musso, L.; Borgonovo, G.; Mazzini, S. Investigation of the Interaction between Aloe vera Anthraquinone Metabolites and c-Myc and C-Kit G-Quadruplex DNA Structures. Int. J. Mol. Sci. 2022, 23, 16018. https://doi.org/10.3390/ijms232416018
Dallavalle S, Artali R, Princiotto S, Musso L, Borgonovo G, Mazzini S. Investigation of the Interaction between Aloe vera Anthraquinone Metabolites and c-Myc and C-Kit G-Quadruplex DNA Structures. International Journal of Molecular Sciences. 2022; 23(24):16018. https://doi.org/10.3390/ijms232416018
Chicago/Turabian StyleDallavalle, Sabrina, Roberto Artali, Salvatore Princiotto, Loana Musso, Gigliola Borgonovo, and Stefania Mazzini. 2022. "Investigation of the Interaction between Aloe vera Anthraquinone Metabolites and c-Myc and C-Kit G-Quadruplex DNA Structures" International Journal of Molecular Sciences 23, no. 24: 16018. https://doi.org/10.3390/ijms232416018
APA StyleDallavalle, S., Artali, R., Princiotto, S., Musso, L., Borgonovo, G., & Mazzini, S. (2022). Investigation of the Interaction between Aloe vera Anthraquinone Metabolites and c-Myc and C-Kit G-Quadruplex DNA Structures. International Journal of Molecular Sciences, 23(24), 16018. https://doi.org/10.3390/ijms232416018