Abstract
Camptothecin (CPT) and its derivatives from Camptotheca acuminata have antitumor effects as a DNA topoisomerase I inhibitor. Previous studies have shown that application of exogenous abscisic acid (ABA) significantly promoted the accumulation level of CPT and induced the expression of CPT biosynthetic genes, which revealed that ABA signaling is effectively involved in regulating CPT biosynthesis in C. acuminata. In this study, an ABA transporter, CaABAT, which encodes a plasma membrane protein belonging to the ABCG subfamily, was identified in C. acuminata, and its ABA import activity was confirmed by transport assay in yeast cells. Real-time PCR analysis showed that CaABAT was predominately expressed in C. acuminata leaves and its expression could be significantly upregulated by exogenous ABA treatment. Silencing of CaABAT down-regulated the expression of ABA response genes, which indicated that translocation of ABA by CaABAT should initiate changes in plant physiological status in response to ABA signaling, thus leading to decreased expression of CPT biosynthesis pathway genes and low accumulation levels of CPT in C. acuminata.
1. Introduction
Camptothecin (CPT), a monoterpene indole alkaloid (MIA), was initially isolated and identified from Camptotheca acuminata. Later investigation showed that CPT and its derivatives could inhibit DNA topoisomerase I activity, which led to it being used as a promising anti-tumor drug for treating cancer diseases in clinical therapy [1,2,3]. However, similarly to other valuable specialized metabolites in plants, low accumulation of CPT in C. acuminata limits its supply in the market [4]. In order to promote the production of CPT in plants, many strategies have been applied, such as the overexpression of pathway genes, and application of elicitors and/or precursor compounds in suspension cells and/or hairy root cultures. Some plant hormones, such as jasmonic acid (JA) [5,6], gibberellic acid (GA) [7], salicylic acid (SA) [8], and abscisic acid (ABA) [9], have already been identified to be involved in regulating plant specialized metabolite biosynthesis as effective elicitors, and many previous reports also revealed that application of these plant hormones could successfully induce the expression of CPT biosynthesis genes, thus promoting CPT accumulation in C. acuminata [5,6,7,8,9,10].
ABA, as the most effective elicitor for promoting CPT biosynthesis according to a previous report [10], is principally biosynthesized in vascular tissues and then transported to the target cells (such as the guard cells) through specific transporters to achieve its biological functions [10,11,12]. In Arabidopsis thaliana, several ABA transporters have been identified, such as AtABCG25, AtABCG31, AtABCG30, AtABCG40, AIT1/NRT1.2, DTX50, and NPF5.1 [13,14,15,16,17].
Among these ABA transporters, AtABCG40 is localized at the plasma membrane and functions as an ABA importer for importing ABA into guard cells [18], where CPT could be effectively synthesized in C. acuminata [19]. Plants lacking functional AtABCG40 exhibit less sensitivity to ABA, including reduced expression of ABA-responsive genes and impaired stomatal closure [18].
In the present study, a homolog of AtABCG40, CaABAT, was isolated from C. acuminata. Its ABA transport activity was determined in yeast cells and its effects on CPT biosynthesis were also investigated in C. acuminata plants. Our results strongly suggest that regulation of plant hormone translocation by certain specific transporters could be a feasible strategy for effective production of valuable plant specialized metabolites.
2. Results
2.1. Isolation and Characterization of CaABAT
Sequencing results indicate that CaABAT contains 4374 bp of coding region, and it encodes 1458 amino acids with 168.12 kDa. For analyzing conserved domains of CaABAT, the amino acid sequences of CaABAT with AtABCG40 were aligned using BLAST and InterPro. The results showed that the homology of the two sequences reached 70%, and the homologous proteins contained specific domains for transporters, an ABC transporter domain (81–143 aa), ATP binding cassette transporter PDR-like subfamily G domain 1 (152–425 aa) and domain 2 (857–1112 aa), ABC-2-type transporter (504–716 and 1185–1399 aa), plant PDR ABC transporter-associated (721–784 aa), and ATPase domain (177–409 and 897–1089 aa) (Figure 1A). Furthermore, phylogenetic analysis of CaABAT with other ABA transporters was performed (Figure 1B), and a prediction of the secondary structure suggested that the protein had six transmembrane structures outside and inside the bilayer region (Figure 1C and Figure S1). These results demonstrate that CaABAT encodes a plasma membrane protein and belongs to the ABCG subfamily.
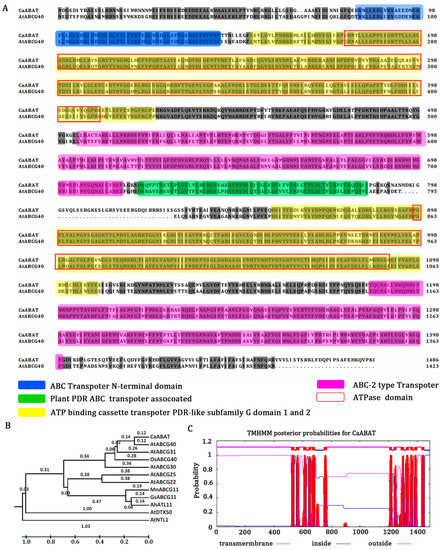
Figure 1.
CaABAT is a member of the ABC transporter family. (A) Alignment of amino acid sequences of CaABAT with AtABCG40. ABC transporter domain (CaABAT 81–143 aa), ATP binding cassette transporter PDR-like subfamily G domain 1 (CaABAT 152–425 aa) and domain 2 (857–1112 aa), ABC-2 type transporter (504–716 aa, 1185–1399 aa), ABC transporter associated (CaABAT 721–784 aa), ATPase domain (CaABAT 177–409 aa, 897–1089 aa). (B) Phylogenetic relationships of CaABAT with other ABA transporters by MEGA6. AtABCG31: Q7PC88.1; AtABCG30: Q8GZ52.2; GsABCG11: KHN26506; MnABCG11: XP_010094142; AtABCG11: Q8RXN0.1; AtABCG40: Q9M9E1.1; AtABCG25: Q84TH5.1; AtNTL1: Q8H157.1; AtDTX50: Q9FJ87.1; AtABCG22: Q93YS4.1; AhATL11: AQW44869; CaABAT: AUB45106; OsABCG40: Q8GU85. (C) Transmembrane structure of CaABAT was analyzed using TMHMM 2.0 (online at http://www.cbs.dtu.dk/services/, accessed on 2 November 2017). CaABAT protein contains a total of six transmembrane structures. Transmembrane helical structures are represented by amino acid residues 520–542, 555–577, 605–627, 640–659, 664–686, 747–769, 1204–1223, 1236–1253, 1284–1306, 1319–1341, 1351–1370, 1377–1399, and 1428–1450 of the CaABAT protein. Consequently, residues 543–554, 628–639, 687–746, 1224–1235, 1307–1318, 1371–1376, and 1451–1487 are located inside the bilayer (i.e., are intracellular). Amino acid residues 1–519, 578–604, 660–663, 770–1203, 1254–1283, 1342–1350, and 1400–1427 are outside the bilayer.
2.2. Functional Identification of CaABAT in Yeast Cells
To examine whether CaABAT functions as an ABA transporter, CaABAT was expressed in the yeast strain AD12345678, which lacks eight major yeast ABC transporter genes [20]. The results showed that ABA could inhibit yeast growth, and this inhibition was more obvious by overexpressing CaABAT in yeast cells, which indicated that CaABAT might have ABA import activity, thus increasing the accumulation level of ABA in cells to raise the inhibitory effect of ABA on yeast growth (Figure 2A). To further confirm ABA transport activity of CaABAT, a transport assay was carried out by analyzing uptake amounts in yeast cells with overexpressing CaABAT compared to that with overexpressing empty vector. Results showed that the accumulation levels of ABA were much higher in yeast cells by overexpressing CaABAT in comparison with empty vector control, which validated the ABA import activity of CaABAT in yeast cells (Figure 2B).
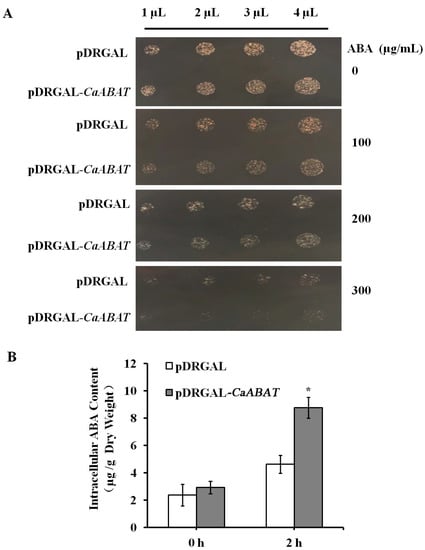
Figure 2.
CaABAT is an ABA influx transporter. (A) The growth of CaABAT yeast transformants in a medium containing ABA. 1 to 4 μL of yeast cultures (OD600 = 0.1) were spotted on a 1/2 SG/-URA medium plate for 24 h at 28 °C. (B) ABA import activity of CaABAT. Yeast cells transformed with pDRGAL and pDRGAL-CaABAT were suspended in 1/2 SG/-URA medium supplemented with 100 µM ABA. The error bars represent standard deviations from three biological replicates, and asterisks indicate statistically significant differences compared with pDRGAL. * p < 0.05.
2.3. Expression Analysis and Subcellular Localization of CaABAT
For analyzing the expression profile of CaABAT, three tissues (root, stem, and leaf) from C. acuminata plant were collected for gene expression analysis by real-time PCR. Results showed that CaABAT was predominately expressed in leaves, while the relative expression levels of CaABAT in root and stem were about 10 times lower than that in leaves (Figure 3A). Furthermore, an ABA responsive gene [21], CaRD29B, which could be induced by ABA treatment, was also investigated for its expression profile in C. acuminata plant. Results showed that the highest expression level of CaRD29B was observed in leaves, which was similar to the expression profile of CaABAT. These similar expression profiles of CaABAT and CaRD29B imply that CaABAT might participate in ABA signaling transduction in response to abiotic stresses by translocating ABA to trigger increased expression of CPT biosynthesis genes for promoting CPT accumulation in C. acuminata leaves (Figure 3B).
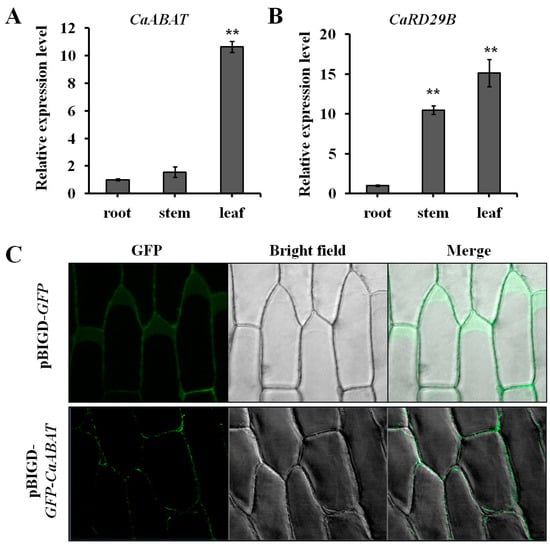
Figure 3.
Expression analysis and subcellular localization of CaABAT. (A) Tissue-specific expression analysis of CaABAT in C. acuminata. (B) Tissue-specific expression analysis of CaRD29B in C. acuminata. (C) Subcellular localization of CaABAT in onion epidermal cells. The error bars represent standard deviations from three biological replicates, and asterisks indicate statistically significant differences compared with pDRGAL. ** p < 0.01.
To determine the subcellular localization of CaABAT, both pBIGD-GFP and pBIGD-GFP-CaABAT vectors were transformed into Agrobacterium for treating onion epidermal cells. GFP signals were detected by a confocal microscope, and results showed that GFP-CaABAT fusion protein localized at plasma membrane in onion epidermal cells (Figure 3C). Considering the ABA import activity of CaABAT in yeast cells, CaABAT should function as an ABA importer in C. acuminata.
2.4. Exogenous Plant Hormone Treatments on C. acuminata Seedlings
Exogenous ABA was applied to treat C. acuminata seedlings for analyzing the expression of CaABAT and CPT biosynthesis pathway genes in response to ABA treatments. The seedlings were treated with ABA for 2 and 4 h, separately, and the expression of CaABAT and CaRD29B could be successfully induced (Figure 4A), which indicated that exogenous ABA triggered a plant response to ABA signaling. For investigating the effect of exogenous ABA on the expression of CPT biosynthesis genes, six pathway genes were selected for the examination. Results showed that four pathway genes (CaCYC1, CaG8O, Ca7DLS, and Ca7DLGT) could be successfully induced by ABA treatment, in which the transcript level of CaCYC1 increased about 50 times in response to exogenous ABA treatment in 4 h (Figure 4B). For analyzing the accumulation levels of CPT, seedlings were treated with ABA for 24, 48, and 72 h. It was shown that the highest accumulation level could be observed after 24 h treatment, and then decreased to original levels (Figure 4C).
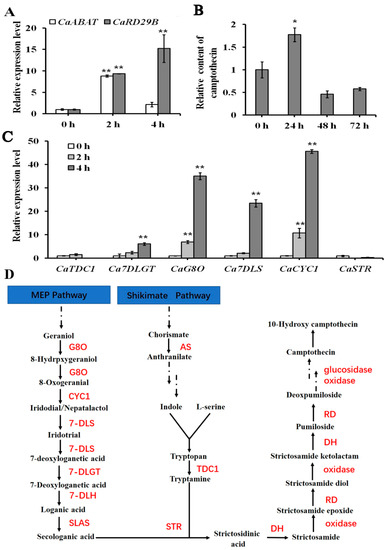
Figure 4.
Gene expression and CPT accumulation analysis in response to exogenous ABA treatment in C. acuminata leaves. (A) Relative expression levels of CaABAT and CaRD29B in response to ABA treatments. (B) Accumulation levels of CPT in response to ABA treatments. (C) Relative expression levels of CPT biosynthesis genes in response to ABA treatments. The error bars represent standard deviations from three biological replicates, and asterisks indicate statistically significant differences compared with 0 h. * p < 0.05, ** p < 0.01. (D) Proposed CPT biosynthesis pathway in C. acuminata [22]. G8O, geraniol-8-oxidase; CYC1, Cyclase 1; 7DLS, 7-deoxyloganetic acid synthase; 7-DLGT, 7-deoxyloganetic acid glucosyltransferase; 7-DLH, 7-deoxyloganic acid hydroxylase; SLAS, secologanic acid synthase; AS, anthranilic acid synthetase; TDC, tryptophan decarboxylase; STR, strictosidinic acid synthase; DH, dehydration; RD, reduction.
Besides ABA, two other important plant hormones, MeJA and SA, were also selected for testing their effects on the expression of CaABAT. Results showed that both MeJA and SA had only slight effects for inducing the expression of CaABAT, while obvious inducing the expression of responding genes, CaMYC2 and CaNPR1, could be observed (Figure S2). These gene expression profiles of CaABAT in responding to different plant hormone treatments demonstrated that CaABAT should be mainly involved in ABA signaling for regulating CPT biosynthesis in C. acuminata.
2.5. Silencing and Overexpression of CaABAT in C. acuminata
To further investigate the effect of CaABAT on the biosynthesis of CPT, the VIGS approach was carried out for silencing the expression of CaABAT in C. acuminata leaves (Figure 5). Results showed that the accumulation level of CPT decreased (by approximately 33%) in CaABAT-silenced leaves in comparison with that in empty vector control leaves (Figure 5B), while the expression levels of CPT biosynthesis pathway genes (CaCYC1, CaG8O, and Ca7DLS) were down-regulated in CaABAT-silenced leaves (Figure 5C). Silencing of CaABAT also resulted in decreased expression of ABA responding gene, CaRD29B (Figure 5A), which indicated that inefficient import of ABA mediated by CaABAT could affect translocation of ABA in cells, thus reducing CPT production by repressing ABA signaling in C. acuminata leaves. Interestingly, when CaABAT was overexpressed in C. acuminata leaves, no obvious variation in CPT accumulation levels could be observed between CaABAT-overexpressing and empty vector control leaves (Figure S3A). Since the biosynthesis of CPT in C. acuminata reveals complicated cell-type specific compartmentation [19], overexpression of CaABAT in all cell types might not guarantee endogenous that ABA is efficiently moved into target cell types, where CPT is synthesized, for promoting CPT biosynthesis. Down-regulation of CPT biosynthesis genes and the ABA biosynthesis gene, CaNCED, and up-regulation of the ABA responding gene, CaRD29B (Figure S3B), also verified cell-type specific compartmentation for CPT biosynthesis in C. acuminata.
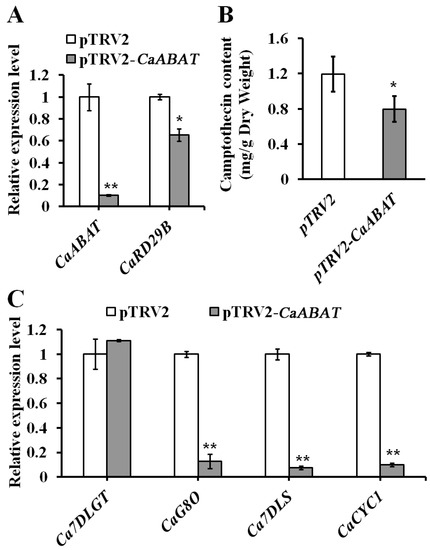
Figure 5.
Virus-induced gene silencing (VIGS) of CaABAT in C. acuminata leaves. (A) Relative expression levels of CaABAT and CaRD29B in pTRV2 control and CaABAT-silenced leaves. (B) CPT accumulation levels in pTRV2 control and CaABAT-silenced leaves. (C) Relative expression levels of CPT biosynthesis genes in pTRV2 control and CaABAT-silenced leaves. The error bars represent standard deviations from three biological replicates, and asterisks indicate statistically significant differences compared with pTRV2 control. * p < 0.05, ** p < 0.01.
3. Discussion
3.1. CaABAT Belongs to the ABCG Subfamily and Is Involved in ABA Transport
As an important plant stress hormone, ABA pervasively exists in plants. The level of endogenous ABA is a major determinant of ABA sensing that is maintained by ABA biosynthesis, catabolism, and transport. At present, ABA transporters from Arabidopsis thaliana, such as AtDTX50, AtABCG25, AtABCG40, AtABCG31, AtABCG30, AIT1, and NPF5.1, have been thoroughly studied [13,14,15,16,17,18]. These transporters belong to three protein families: the multidrug and toxin efflux transporter family, nitrate transporter 1/peptide transporter family, and ABCG family [13,14,15,16]. Among these ABA transporters, AtABCG40 was identified to be importing ABA into guard cells [18], which is the major place for CPT biosynthesis in C. acuminata according to a previous report [19]. Recently, a homolog of AtABCG40 from Artemisia annua, AaABCG40, was also identified as an ABA transporter, which demonstrates conserved function of ABCG40 homologs for transporting ABA in planta [23].
In the present study, we successfully isolated an ABCG40 homolog (CaABAT) from C. acuminata, and sequence analysis showed that this identified CaABAT has high homology and contains the same conserved domain with AtABCG40 (Figure 1). For further understanding of the structure and possible function of CaABAT, a 3D model of CaABAT was constructed through subsection homology modeling using a Swiss-model server. The results showed that the crystal structure of ATP-binding cassette subfamily G member 2 (PDB code: 5nj3), as the template, shares 31.55% sequence identity and 36% similarity with 167–635 aa of CaABAT (as Model 1 in Figure S1A). Moreover, ATP-binding cassette sub-family G member 5 (PDB code: 5do7) shares 32.13% sequence identity and 35% similarity with 857–1455 aa of CaABAT (as Model 2 in Figure S1B). These templates were then artificially assembled into 3D models (Figure S1C). ATP-binding cassette transporters are a highly conserved family of ATP-driven pump proteins consisting of two hydrophobic transmembrane domains (TMDs) and two cytosolic domains known as the nucleotide-binding domains (NBDs) or nucleotide-binding folds (NBFs). The two hydrophobic TMDs constitute a membrane-spanning pore (the red boxes), while the two cytosolic domains contain the ATP-binding motifs (the yellow box). The structure of this protein is consistent with a previous study [24]. Thus, we hypothesized that ABA may cause partial conformational changes in TMD and NBD by binding to the TMD of the transporter and activating NBD to release ADP.
3.2. Effect of CaABAT on ABA-Regulated Camptothecin Biosynthesis
Once abiotic stressors or developmental cues up-regulate the accumulation levels of endogenous ABA, PYR/PYL/RCAR (an ABA receptor) binds ABA and interacts with PP2C (a negative regulator) to inhibit protein phosphatase activity [25]. Then, SnRK2 (a positive regulator) is released from PP2C-dependent regulation and activated to phosphorylate downstream factors, such as the AREB/ABF bZIP-type transcription factor or membrane proteins involving ion channels [26]. After the phosphorylation of the bZIP transcription factor binding response element ABRE, the expression of ABA response genes was activated. Subsequently, ABA response was launched to resist external stimuli, such as the regulation of seed germination, the biosynthesis of secondary metabolites, stomatal closure, and drought resistance [26]. Although ABA transporters have been validated as signal cascade amplifiers in ABA signaling, efficient function of these transporters still depends on ABA accumulation levels in plants. In this work, since there was no excessive ABA accumulation in plants, only reduced accumulation levels of CPT could be observed in CaABAT-silenced leaves, while no variation in CPT accumulation levels could be found in CaABAT-overexpressing leaves. Therefore, appropriate regulation of ABA biosynthesis could be very important for ABA transporter-mediated promotion of CPT biosynthesis in C. acuminata.
By silencing the expression of CaABAT, four CPT biosynthesis pathway genes in the iridoid pathway could be obviously regulated, which demonstrates that the iridoid pathway should be the target of ABA signaling for regulating CPT biosynthesis in C. acuminata. Further, promoter sequence analysis showed that ABA responding element (ACGTG) could be found in Ca7DLGT (from −987 to −992) and CaG8O (from −263 to −268) genes (Table S2), which additionally indicates the expression profile of these genes for responding to ABA signaling.
It is well known that ABA, as a signaling molecule, plays very important roles in plants, and it could be produced in response to stress, such as drought and low temperature. In Artemisia annu, it has been reported that the ABCG40 homolog AaABCG40 could enhance artemisinin accumulation and modulate drought tolerance [23]. In this work, we validated the importance of ABA signaling to CPT production mediated by the ABCG40 homolog, CaABAT. Considering that plants respond to most abiotic stresses through the ABA signaling pathway, translocation of endogenous ABA by ABA transporters, such as CaABAT, could be an important biological process for connecting the correlation of abiotic stresses and CPT biosynthesis in C. acuminata.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
C. acuminata seedlings were obtained by in vitro rapid propagation described in our previous report [22]. After 45 days of growth, seedlings were transferred to soil and grown at 25 °C with a 16 h photoperiod provided by cool white fluorescent light (40 μmol m−2s−l). After 4 weeks of growth, the new leaves were selected for the following experiments.
4.2. Isolation and Characterization of CaABAT
A homolog of AtABCG40 in C. acuminata was obtained by blast analysis in the C. acuminata transcriptome database (http://medicinalplantgenomics.msu.edu/, accessed on 8 March 2017), and then this ABA transporter was designated as Camptotheca acuminata ABA transporter (CaABAT). The full-length cDNA of CaABAT with KpnI/SacI at both ends was amplified by nested PCR using the following primers (Table S1): first pair, CaABAT-FL-F and CaABAT-FL-R; second pair: CaABAT-KpnI-F and CaABAT-SacI-R. PCR products were then ligated into the pGEM-T easy vector (Promega, Beijing) to produce plasmid T-CaABAT. The CaABAT amino acid sequence and transmembrane structure were analyzed using MEGA6 and TMHMM 2.0, respectively. Then, conserved domains of CaABAT were analyzed using InterPro 91.0 [27,28].
4.3. RNA Extraction and Real-Time PCR Analysis
Plant tissues were ground into a fine powder within liquid nitrogen, and the total RNA was extracted using TRIzol reagent (Invitrogen, Shanghai, China). The RNA pellets were dissolved in DEPC-treated water, and the quality and quantity of total RNA were determined at OD 260 and 280 nm using a UV-2400 spectrophotometer (Shanghai Sunny Hengping Scientific Instrument Co., Ltd., Shanghai, China). After the extracted RNA was digested with DNase I, first-strand cDNA was synthesized using an M-MLV RTase cDNA Synthesis kit (TaKaRa Biotech Co., Ltd., Dalian, China) for real-time PCR analysis. The expression levels of target genes were analyzed using LightCycler 480 software (Roche) and normalized to CaUBC through the 2–ΔΔCT method [29]. Primers were designed based on sequences from CaUBC and other target genes (Table S1).
4.4. Transient Overexpression and Virus-Induced Gene Silencing (VIGS) of CaABAT in C. acuminata
For overexpression vector construction, full-length cDNA fragment of CaABAT was obtained from T-CaABAT digested by KpnI/SacI and cloned to pBIGD overexpression vector pre-digested with KpnI/SacI to produce pBIGD-CaABAT vector. Both empty vector pBIGD and pBIGD-CaABAT were then transformed into Agrobacterium tumefaciens strain GV3101 to treat C. acuminata leaves [22]. The plants were grown for 3 weeks until transformed leaves were harvested for further gene expression and CPT accumulation analysis. For constructing VIGS vectors, a 543 bp CaABAT fragment with KpnI/SacI restriction sites at both ends was PCR-amplified by using T-CaABAT plasmid as the template. The PCR product was then cloned into pGEM-T easy vector, and the CaABAT fragment was then obtained by KpnI/SacI double digestion. The CaABAT fragment was mobilized to the pTRV2 vector pre-digested with KpnI/SacI to produce pTRV2-CaABAT. The pTRV1, pTRV2, and pTRV2-CaABAT vectors were transformed into Agrobacterium strain GV3101, and VIGS experiments were carried out according to the method developed previously in the lab [30]. The plants were grown for 3 weeks until VIGS-treated leaves were harvested for further gene expression and CPT accumulation analysis.
4.5. Functional Identification of CaABAT in Yeast Cells
For yeast overexpression vector construction, pDRGAL vector was digested with NotI, followed by dephosphorylation of the digested backbone of pDRGAL. Full-length cDNA of CaABAT was obtained by NotI digestion of T-CaABAT plasmid and cloned to pre-treated pDRGAL vector mentioned above to produce pDRGAL-CaABAT. The pDRGAL and pDRGAL-CaABAT vectors were then transformed into the yeast strain AD12345678. The transformants containing pDRGAL and pDRGAL-CaABAT vectors were grown in SD/-URA medium until OD 600 nm = 0.8. A 1–4 µL amount of yeast cultures was then spotted on 1/2 SG/-URA medium plates containing 0, 100, 200, 300, or 500 µg/mL [20] of ABA, and the inhibitory effect of ABA on yeast growth was observed.
For further confirming ABA transport activity of CaABAT, transport assay in yeast cells was carried out. Yeast transformants containing pDRGAL and pDRGAL-CaABAT vectors were grown on SD/-URA medium until OD600 = 0.8. Yeast cells were then collected by centrifugation and re-suspended in 1/2 SG/-URA medium containing 100 µM of ABA with shaking at 180 rpm. Cells were harvested at the indicated time by centrifugation and lysed in a TissueLyser after being washed with distilled water three times. ABA accumulated inside yeast cells was then extracted in methanol, and ABA amounts were determined by HPLC analysis [22].
4.6. Subcellular Localization of CaABAT
The coding region of GFP (with stop codon) was PCR-amplified and cloned to pBIGD vector to produce pBIGD-GFP. For GFP-CaABAT fusion protein expression, both coding regions of GFP (without stop codon) and CaABAT were fused by PCR and then cloned to pBIGD vector to produce pBIGD-GFP-CaABAT. The constructed pBIGD-GFP and pBIGD-GFP-CaABAT vectors were transformed into Agrobacterium tumefaciens GV3101 to treat onion epidermal cells according to our previous report [20]. The GFP signal of the transformed cells was detected by laser scanning confocal microscopy (LSCM, Zeiss, Oberkochen, Germany).
4.7. Exogenous Plant Hormone Treatment
For evaluating the effect of exogenous plant hormones on pathway gene expression in C. acuminata leaves, seedlings were gently removed from soil, soaked in 100 µM ABA, 20 µM MeJA, or 20 µM SA for 0, 2, or 4 h. The leaves were harvested at the indicated time for target gene expression analysis. For analyzing the effect of ABA on CPT accumulation in C. acuminata leaves, seedlings were sprayed with 100 µM of ABA containing 0.1% DMSO (ABA was pre-dissolved in DMSO) for 0, 24, 48, or 72 h, while DMSO was used as negative control. Leaves were then harvested at the indicated time, and CPT contents were determined by HPLC analysis [22].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232416120/s1.
Author Contributions
Y.W. (Yanyan Wang) designed the study. Y.W. (Yang Wang), H.B. and Y.H. performed the experiments. Y.W. (Yanyan Wang) and Y.H. performed data analyses. Y.W. (Yanyan Wang) and F.Y. wrote the manuscript and critically reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (no. 31970324 and 42177112), Science and Technology Basic Resources Investigation Program of China (no. 2019FY100500), and Department of Education of Liaoning Province (no. J2020042).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ko, A.H.; Tempero, M.A.; Shan, Y.S.; Su, W.C.; Lin, Y.L.; Dito, E.; Ong, A.; Wang, Y.W.; Yeh, C.G.; Chen, L.T. A multinational phase 2 study of nanoliposomal irinotecan sucrosofate (PEP02, MM-398) for patients with gemcitabine-refractory metastatic pancreatic cancer. Br. J. Cancer 2013, 109, 920–925. [Google Scholar] [CrossRef]
- Hamaguchi, T.; Doi, T.; Eguchi-Nakajima, T.; Kato, K.; Yamada, Y.; Shimada, Y.; Fuse, N.; Ohtsu, A. Phase I study of NK012, a novel SN-38- incorporating micellar nanoparticle, in adult patients with solid Tumors. Clin. Cancer Res. 2010, 16, 5058–5066. [Google Scholar] [CrossRef]
- Wen, Y.; Wang, Y.; Liu, X.; Zhang, W.; Xiong, X.; Han, Z.; Liang, X. Camptothecin based nanodrug delivery systems. Cancer Biol. Med. 2017, 14, 363–370. [Google Scholar]
- Lorence, A.; Nessler, C.L. Camptothecin, over four decades of surprising findings. Phytochemistry 2004, 65, 2735–2749. [Google Scholar] [CrossRef]
- Curaba, J.; Singh, M.B.; Bhalla, P.L. miRNAs in the crosstalk between phytohormone signalling pathways. J. Exp. Bot. 2014, 65, 1425–1438. [Google Scholar] [CrossRef]
- El-Sayed, M.; Verpoorte, R. Methyl jasmonate accelerates catabolism of monoterpenoid indole alkaloids in Catharanthus roseus during leaf processing. Fitoterapia 2005, 76, 83–90. [Google Scholar] [CrossRef]
- Amini, A.; Glevarec, G.; Andreu, F.; Rideau, M.; Crèche, J. Low levels of gibberellic acid control the biosynthesis of ajmalicine in Catharanthus roseus cell suspension cultures. Planta Med. 2009, 75, 187–191. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Khodakovskaya, M.V.; Glazunov, V.P.; Radchenko, S.V.; Zvereva, E.V.; Fedoreyev, S.A.; Zhuravlev, Y.N. Effect of salicylic acid, methyl jasmonate, ethephon and cantharidin on anthraquinone production by Rubia cordifolia callus cultures transformed with the rolB and rolC genes. J. Biotechnol. 2002, 97, 213–221. [Google Scholar] [CrossRef]
- Brookbank, B.P.; Patel, J.; Gazzarrini, S.; Nambara, E. Role of Basal ABA in Plant Growth and Development. Genes 2021, 12, 1936. [Google Scholar] [CrossRef]
- Kai, G.; Teng, X.; Cui, L.; Li, S.; Hao, X.; Shi, M.; Yan, B. Effect on three plant hormone elicitors on the Camptothecin accumulation and gene transcript profiling in Camptptheca Acuminata Seelings. Int. J. Sci. Res. 2014, 3, 86–95. [Google Scholar]
- Boursiac, Y.; Léran, S.; Corratgé-Faillie, C.; Gojon, A.; Krouk, G.; Lacombe, B. ABA transport and transporters. Trends Plant Sci. 2013, 18, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Sugimoto, E.; Shinozaki, K. Intertissue signal transfer of abscisic acid from vascular cells to guard cells. Plant Physiol. 2014, 164, 1587–1592. [Google Scholar] [CrossRef] [PubMed]
- Kuromorim, T.; Shinozaki, K. ABA transport factors found in Arabidopsis ABC transporters. Plant Signal. Behav. 2010, 5, 1124–1126. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Miyaji, T.; Yabuuchi, H.; Shimizu, H.; Sugimoto, E.; Kamiya, A.; Moriyama, Y.; Shinozaki, K. ABC transporter AtABCG25 is involved in abscisic acid transport and responses. Proc. Natl. Acad. Sci. USA 2010, 107, 2361–2366. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, H.; Pan, Y.; Yu, Y.; Luan, S.; Li, L. A DTX/MATE type transporter facilitates abscisic acid efflux and modulates ABA sensitivity and drought tolerance in Arabidopsis. Mol. Plant 2014, 7, 1522–1532. [Google Scholar] [CrossRef] [PubMed]
- Merilo, E.; Jalakas, P.; Laanemets, K.; Mohammadi, O.; Hõrak, H.; Kollist, H.; Brosché, M. Abscisic acid transport and homeostasis in the context of stomatal regulation. Mol. Plant 2015, 8, 1321–1333. [Google Scholar] [CrossRef]
- Shimizu, T.; Kanno, Y.; Watanabe, S.; Seo, M. Arabidopsis NPF5.1 regulates ABA homeostasis and seed germination by mediating ABA uptake into the seed coat. Plant Signal. Behav. 2022, 17, 2095488. [Google Scholar] [CrossRef]
- Kang, J.; Hwang, J.U.; Lee, M.; Kim, Y.Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef]
- Valletta, A.; Trainotti, L.; Santamaria, A.R.; Pasqua, G. Cell-specific expression of tryptophan decarboxylase and 10-hydroxygeraniol oxidoreductase, key genes involved in camptothecin biosynthesis in Camptotheca acuminata Decne (Nyssaceae). BMC Plant Biol. 2010, 10, 69. [Google Scholar] [CrossRef]
- Yu, F.; De Luca, V. ATP-binding cassette transporter controls leaf surface secretion of anticancer drug components in Catharanthus roseus. Proc. Natl. Acad. Sci. USA 2013, 110, 15830–15835. [Google Scholar] [CrossRef]
- Nakashima, K.; Fujita, Y.; Katsura, K.; Maruyama, K.; Narusaka, Y.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulation of ABI3- and ABA-responsive genes including RD29B and RD29A in seeds, germinating embryos, and seedlings of Arabidopsis. Plant Mol. Biol. 2006, 60, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Liu, Z.; Wang, Y.; Tang, Z.; Yu, F. A bZIP transcription factor, CaLMF, mediated light-regulated camptothecin biosynthesis in Camptotheca acuminata. Tree Physiol. 2019, 39, 372–380. [Google Scholar] [CrossRef]
- Fu, X.; Liu, H.; Hassani, D.; Peng, B.; Yan, X.; Wang, Y.; Wang, C.; Li, L.; Liu, P.; Pan, Q.; et al. AaABCG40 Enhances Artemisinin Content and Modulates Drought Tolerance in Artemisia annua. Front. Plant Sci. 2020, 11, 950. [Google Scholar] [CrossRef]
- Ma, Y.; Cao, J.; He, J.; Chen, Q.; Li, X.; Yang, Y. Molecular mechanism for the regulation of ABA homeostasis during plant development and stress responses. Int. J. Mol. Sci. 2018, 19, 3643. [Google Scholar] [CrossRef] [PubMed]
- Beis, K. Structural basis for the mechanism of ABC transporters. Biochem. Soc. Trans. 2015, 43, 889–893. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J.; Sauter, A.; Wichert, K.; Messdaghi, D.; Hartung, W. Extracellular beta-glucosidase activity in barley involved in the hydrolysis of ABA glucose conjugate in leaves. J. Exp. Bot. 2000, 51, 937–944. [Google Scholar] [CrossRef]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381e5. [Google Scholar] [CrossRef]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195e201. [Google Scholar] [CrossRef]
- Jin, Z.; Cong, Y.; Zhu, S.; Xing, R.; Zhang, D.; Yao, X.; Wan, R.; Wang, Y.; Yu, F. Two classes of cytochrome P450 reductase genes and their divergent functions in Camptotheca acuminate Decne. Int. J. Biol. Macromol. 2019, 138, 1098–1108. [Google Scholar] [CrossRef]
- Jin, Z.; Yan, T.; Chang, C.; Liu, Z.; Wang, Y.; Tang, Z.; Yu, F. Application of virus-induced gene silencing approach in Camptotheca acuminata. Plant Cell Tiss. Organ Cult. 2016, 126, 533–540. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).