
Common T-Cell-Receptor Motifs and Features in Patients with Cytomegalovirus (CMV)-Seronegative End-Stage Renal Disease Receiving a Peptide Vaccination against CMV

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
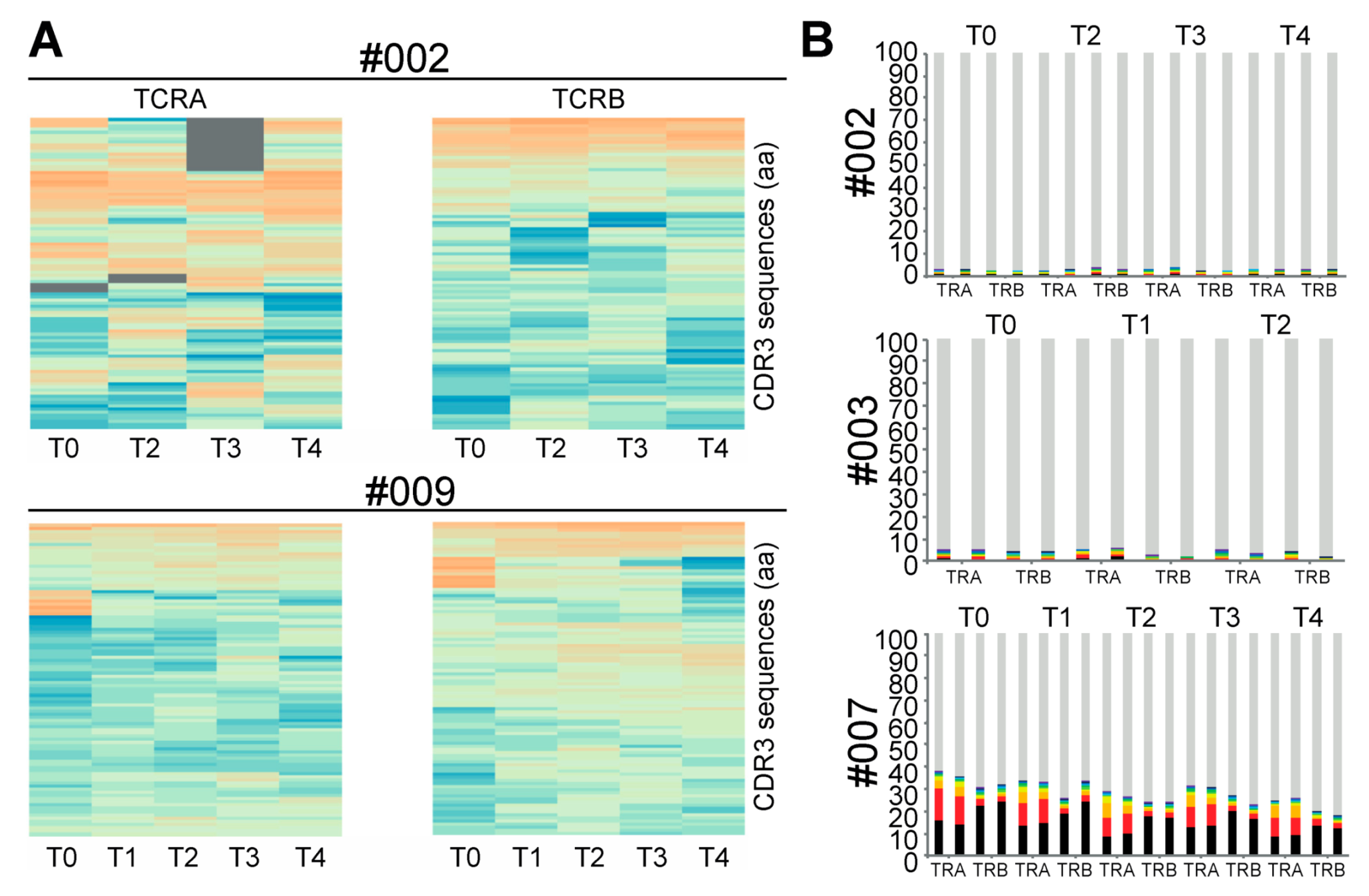
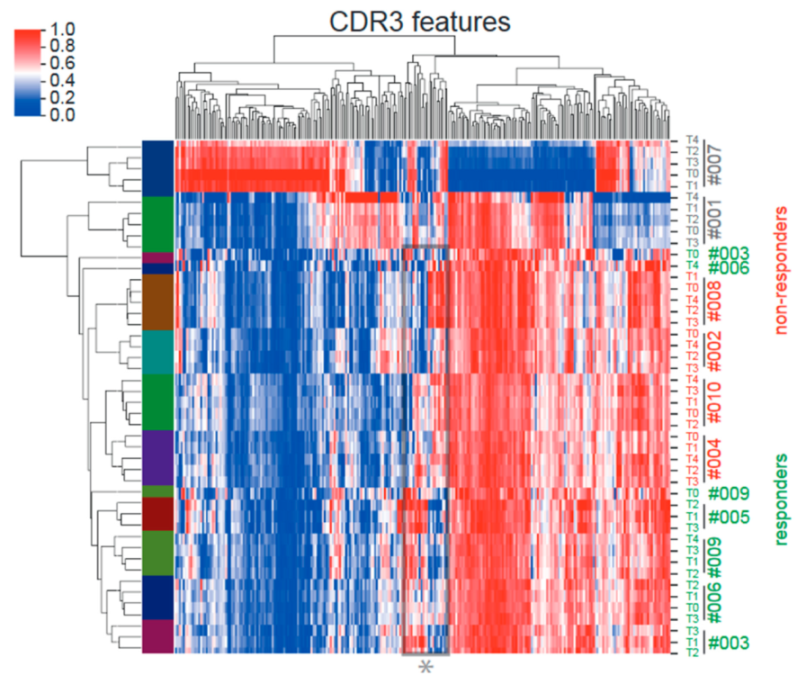
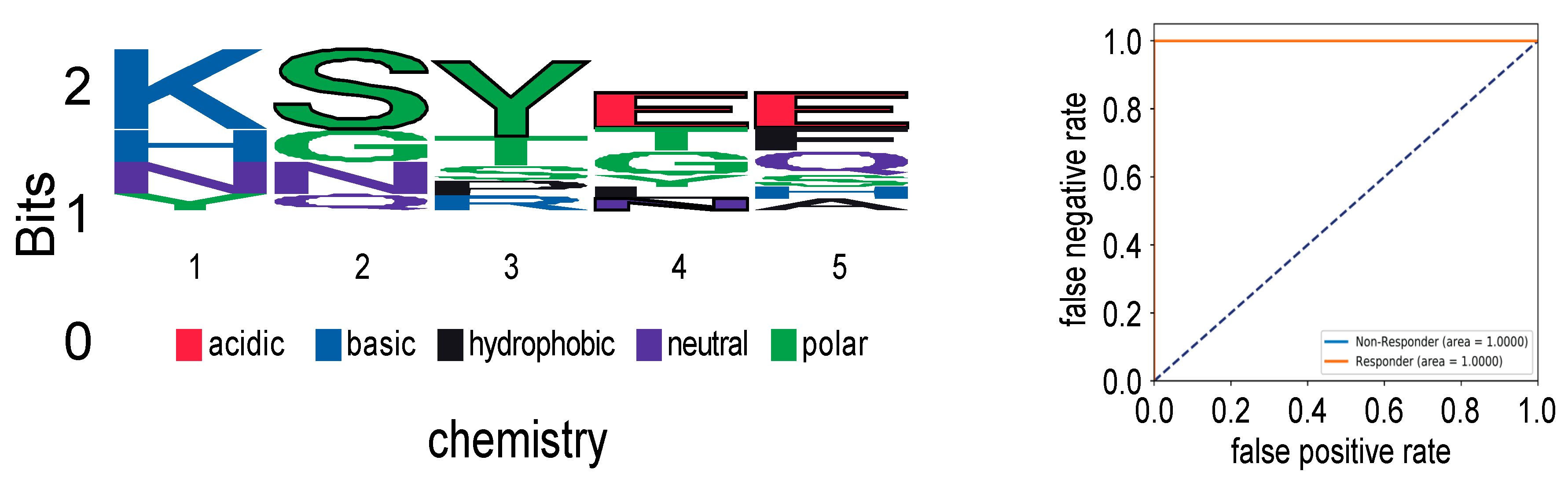
2.1. Impact of CMV-Peptide Vaccination on the TCR Repertoire
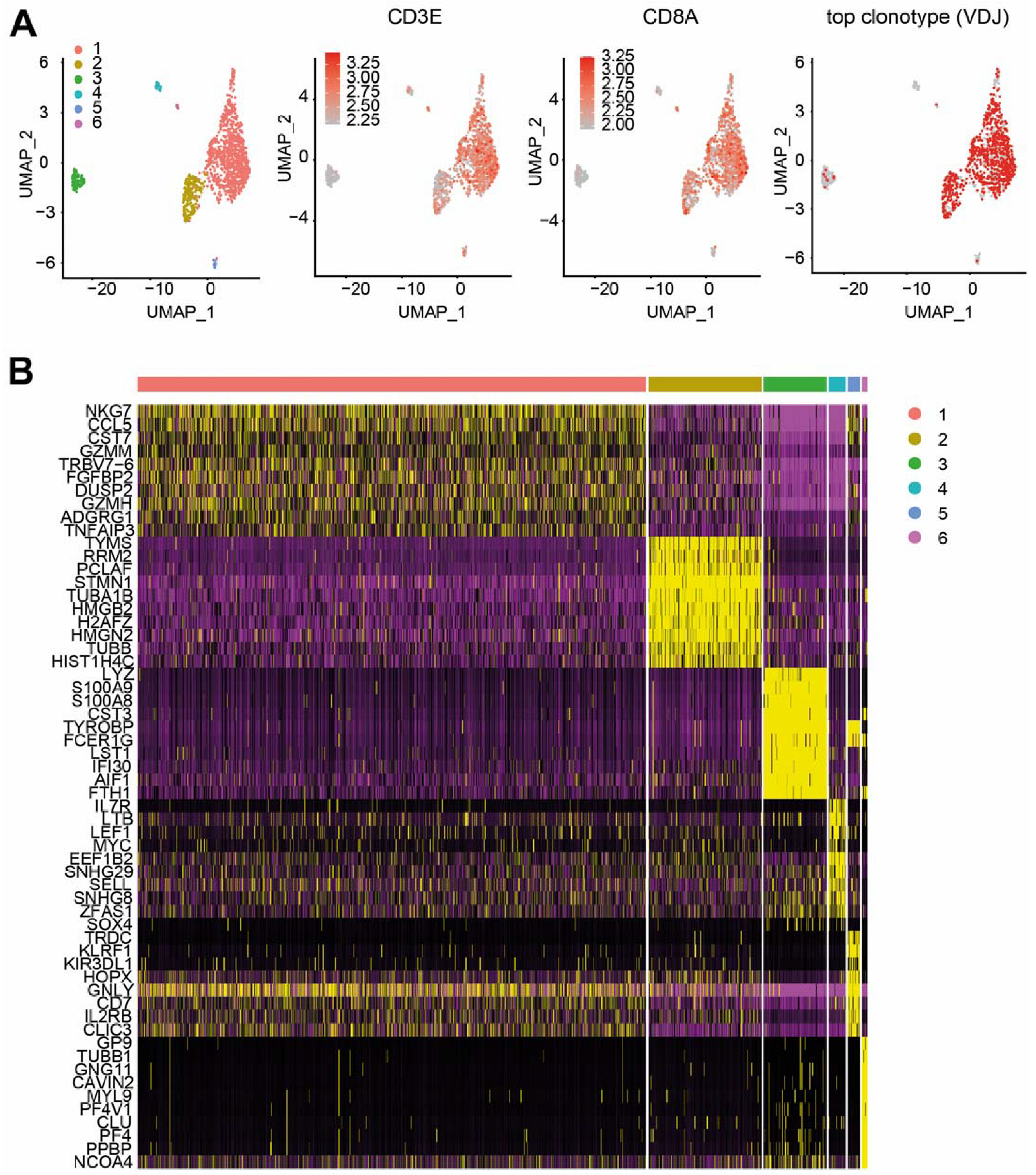
2.2. Combined Single RNA and TCR Sequencing in Follow-Up
3. Discussion
4. Materials and Methods
4.1. Underlying Clinical Study
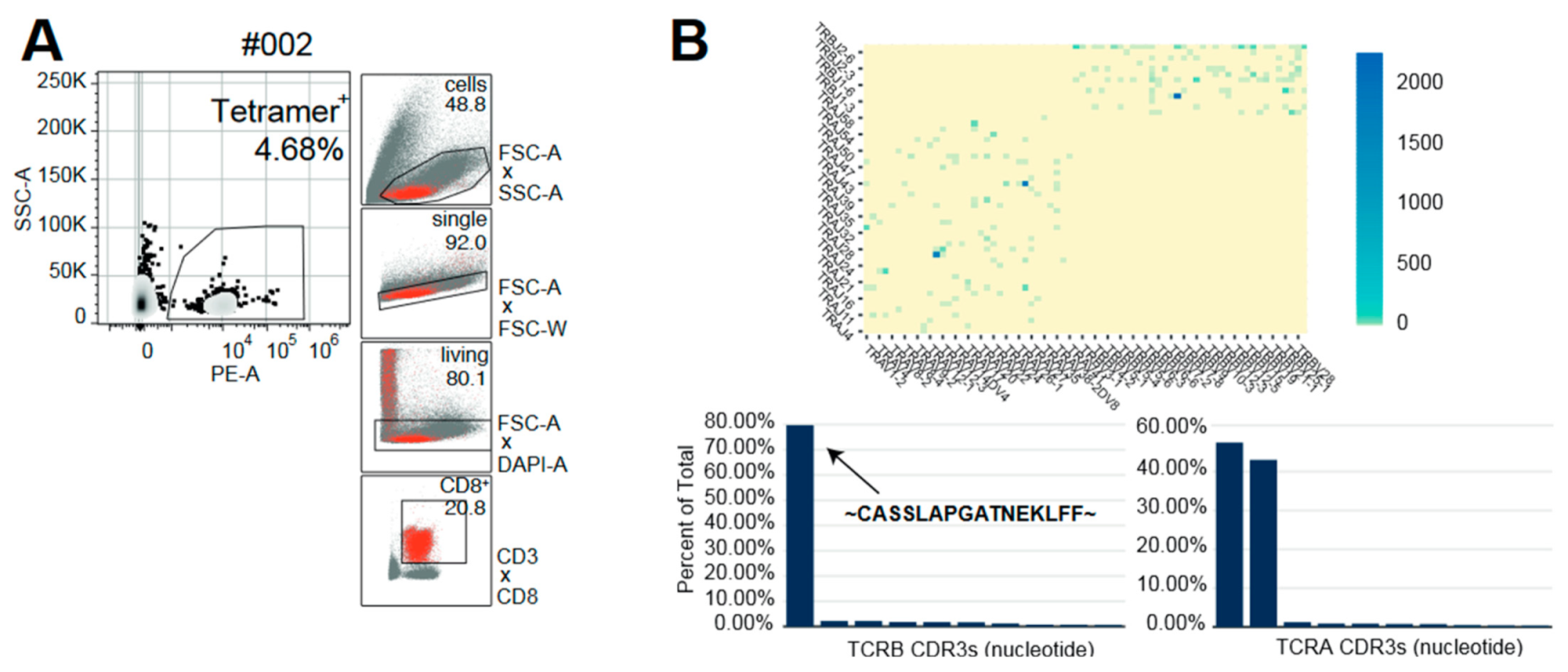
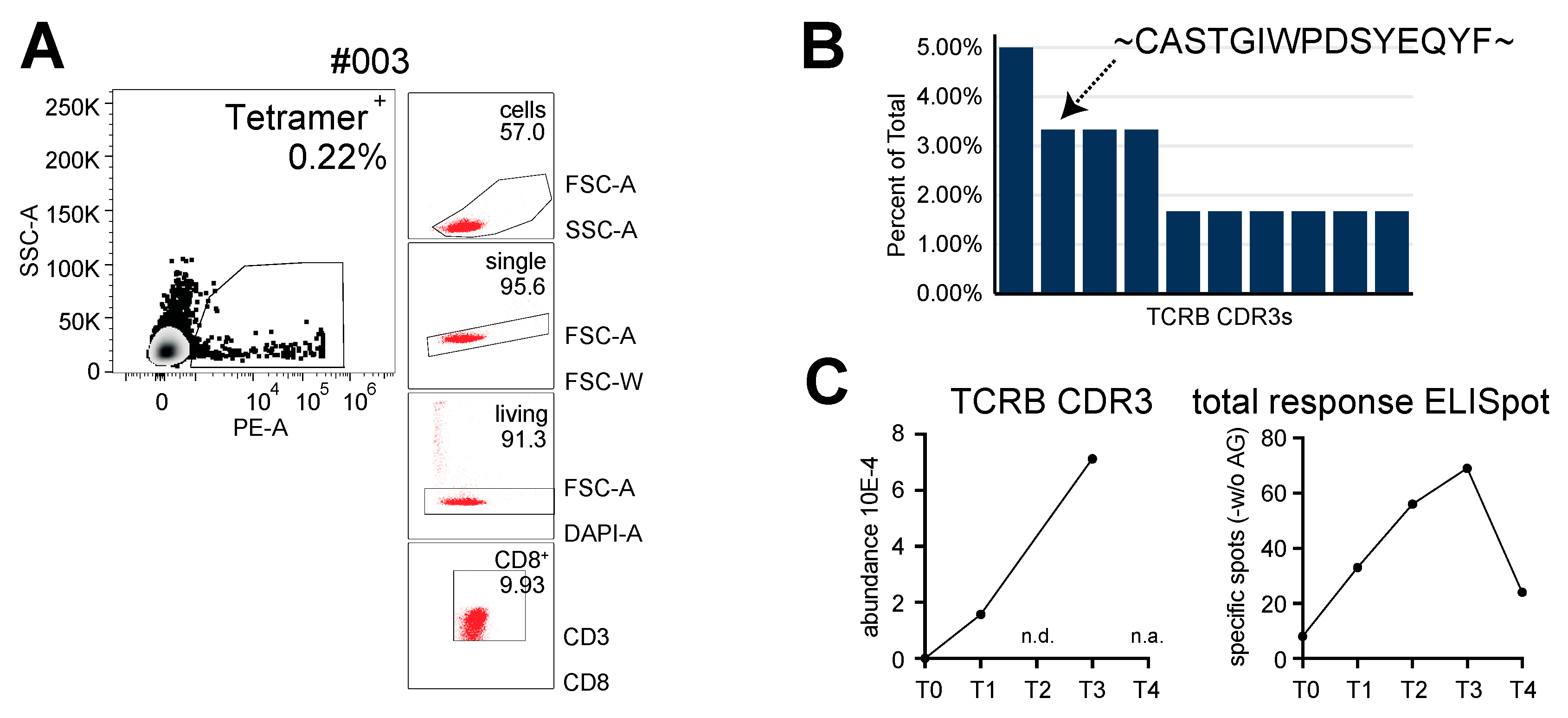
4.2. Tetramer Staining for CMV-Specific CD8+ T Cells
4.3. Bulk T-Cell-Receptor Repertoire Sequencing and Analysis
4.4. Single-Cell RNA and T-Cell-Receptor Library Construction and Sequencing
4.5. Single-Cell Data Analysis
4.6. IFN-γ ELISpot
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Keyzer, K.; Van Laecke, S.; Peeters, P.; Vanholder, R. Human Cytomegalovirus and Kidney Transplantation: A Clinician’s Update. Am. J. Kidney Dis. 2011, 58, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Sagedal, S.; Hartmann, A.; Nordal, K.P.; Osnes, K.; Leivestad, T.; Foss, A.; Degré, M.; Fauchald, P.; Rollag, H. Impact of early cytomegalovirus infection and disease on long-term recipient and kidney graft survival. Kidney Int. 2004, 66, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, E.M.; A Jones, C.; Webster, A.C.; Strippoli, G.F.; Barclay, P.G.; Kable, K.; Vimalachandra, D.; Craig, J.C. Antiviral medications to prevent cytomegalovirus disease and early death in recipients of solid-organ transplants: A systematic review of randomised controlled trials. Lancet 2005, 365, 2105–2115. [Google Scholar] [CrossRef]
- Helanterä, I.; Kyllönen, L.; Lautenschlager, I.; Salmela, K.; Koskinen, P. Primary CMV Infections Are Common in Kidney Transplant Recipients After 6 Months Valganciclovir Prophylaxis. Arab. Archaeol. Epigr. 2010, 10, 2026–2032. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Boppana, S.B. Vaccination against the human cytomegalovirus. Vaccine 2018, 37, 7437–7442. [Google Scholar] [CrossRef]
- Hill, A.B. The immune response to CMV infection and vaccination in mice, monkeys and humans: Recent developments. Curr. Opin. Virol. 2018, 28, 161–166. [Google Scholar] [CrossRef]
- Nakamura, R.; La Rosa, C.; Longmate, J.; Drake, J.; Slape, C.; Zhou, Q.; Lampa, M.G.; O’Donnell, M.; Cai, J.-L.; Farol, L.; et al. Viraemia, immunogenicity, and survival outcomes of cytomegalovirus chimeric epitope vaccine supplemented with PF03512676 (CMVPepVax) in allogeneic haemopoietic stem-cell transplantation: Randomised phase 1b trial. Lancet Haematol. 2015, 3, e87–e98. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, D.I.; Munoz, F.M.; Callahan, S.T.; Rupp, R.; Wootton, S.H.; Edwards, K.M.; Turley, C.B.; Stanberry, L.R.; Patel, S.M.; Mcneal, M.M.; et al. Safety and efficacy of a cytomegalovirus glycoprotein B (gB) vaccine in adolescent girls: A randomized clinical trial. Vaccine 2015, 34, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pass, R.F.; Zhang, C.; Evans, A.; Simpson, T.; Andrews, W.; Huang, M.-L.; Corey, L.; Hill, J.; Davis, E.; Flanigan, C.; et al. Vaccine Prevention of Maternal Cytomegalovirus Infection. N. Engl. J. Med. 2009, 360, 1191–1199. [Google Scholar] [CrossRef]
- Wloch, M.K.; Smith, L.R.; Boutsaboualoy, S.; Reyes, L.; Han, C.; Kehler, J.; Smith, H.D.; Selk, L.; Nakamura, R.; Brown, J.M.; et al. Safety and Immunogenicity of a Bivalent Cytomegalovirus DNA Vaccine in Healthy Adult Subjects. J. Infect. Dis. 2008, 197, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- A Kharfan-Dabaja, M.; Boeckh, M.; Wilck, M.B.; A Langston, A.; Chu, A.H.; Wloch, M.K.; Guterwill, D.F.; Smith, L.R.; Rolland, A.P.; Kenney, R.T. A novel therapeutic cytomegalovirus DNA vaccine in allogeneic haemopoietic stem-cell transplantation: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Infect. Dis. 2012, 12, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Reusser, P.; Riddell, S.R.; Meyers, J.D.; Greenberg, P.D. Cytotoxic T-lymphocyte response to cytomegalovirus after human allogeneic bone marrow transplantation: Pattern of recovery and correlation with cytomegalovirus infection and disease. Blood 1991, 78, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Sommerer, C.; Schmitt, A.; Hückelhoven-Krauss, A.; Giese, T.; Bruckner, T.; Wang, L.; Schnitzler, P.; Meuer, S.; Zeier, M.; Schmitt, M. Peptide Vaccination Against Cytomegalovirus Induces Specific T Cell Response in Responses in CMV Seronegative End-Stage Renal Disease Patients. Vaccines 2021, 9, 133. [Google Scholar] [CrossRef]
- Sidhom, J.-W.; Larman, H.B.; Pardoll, D.M.; Baras, A.S. DeepTCR: A deep learning framework for revealing structural concepts within TCR Repertoire. bioRxiv 2018, 464107. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, C.; Moseman, E.A.; Loughhead, S.M.; Alvarez, D.; Zwijnenburg, A.J.; Waanders, L.; Garg, R.; de la Torre, J.C.; von Andrian, U.H. The Chemokine Receptor CX3CR1 Defines Three Antigen-Experienced CD8 T Cell Subsets with Distinct Roles in Immune Surveillance and Homeostasis. Immunity 2016, 45, 1270–1284. [Google Scholar] [CrossRef] [Green Version]
- Ng, S.S.; Rivera, F.D.L.; Yan, J.; Corvino, D.; Das, I.; Zhang, P.; Kuns, R.; Chauhan, S.B.; Hou, J.; Li, X.-Y.; et al. The NK cell granule protein NKG7 regulates cytotoxic granule exocytosis and inflammation. Nat. Immunol. 2020, 21, 1205–1218. [Google Scholar] [CrossRef]
- Pogorelyy, M.V.; Minervina, A.A.; Shugay, M.; Chudakov, D.M.; Lebedev, Y.B.; Mora, T.; Walczak, A.M. Detecting T cell receptors involved in immune responses from single repertoire snapshots. PLoS Biol. 2019, 17, e3000314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethna, Z.; Elhanati, Y.; Callan, C.G.; Walczak, A.M.; Mora, T. OLGA: Fast computation of generation probabilities of B- and T-cell receptor amino acid sequences and motifs. Bioinformatics 2019, 35, 2974–2981. [Google Scholar] [CrossRef] [Green Version]
- Torre-Cisneros, J.; Aguado, J.; Caston, J.; Almenar, L.; Alonso, A.; Cantisán, S.; Carratala, J.; Cervera, C.; Cordero, E.; Fariñas, M.; et al. Management of cytomegalovirus infection in solid organ transplant recipients: SET/GESITRA-SEIMC/REIPI recommendations. Transplant. Rev. 2016, 30, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Lúcia, M.; Crespo, E.; Cruzado, J.M.; Grinyó, J.M.; Bestard, O. Human CMV-specific T-cell responses in kidney transplantation; toward changing current risk-stratification paradigm. Transpl. Int. 2014, 27, 643–656. [Google Scholar] [CrossRef]
- Huth, A.; Liang, X.; Krebs, S.; Blum, H.; Moosmann, A. Antigen-Specific TCR Signatures of Cytomegalovirus Infection. J. Immunol. 2018, 202, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Yang, X.; Ko, A.; Sun, X.; Gao, M.; Zhang, Y.; Shi, A.; Mariuzza, R.A.; Weng, N.-P. Sequence and Structural Analyses Reveal Distinct and Highly Diverse Human CD8 + TCR Repertoires to Immunodominant Viral Antigens. Cell Rep. 2017, 19, 569–583. [Google Scholar] [CrossRef]
- Emerson, R.; DeWitt, W.S.; Vignali, M.; Gravley, J.; Hu, J.K.; Osborne, E.J.; Desmarais, C.; Klinger, M.; Carlson, C.S.; A Hansen, J.; et al. Immunosequencing identifies signatures of cytomegalovirus exposure history and HLA-mediated effects on the T cell repertoire. Nat. Genet. 2017, 49, 659–665. [Google Scholar] [CrossRef]
- Banas, B.; Steubl, D.; Renders, L.; Chittka, D.; Banas, M.C.; Wekerle, T.; Koch, M.; Witzke, O.; Mühlfeld, A.; Sommerer, C.; et al. Clinical validation of a novel enzyme-linked immunosorbent spot assay-basedin vitrodiagnostic assay to monitor cytomegalovirus-specific cell-mediated immunity in kidney transplant recipients: A multicenter, longitudinal, prospective, observational study. Transpl. Int. 2018, 31, 436–450. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, P.; Jose, M.; Atkins, R.C.; Holdsworth, S.R. Ex vivo lymphocyte proliferative function is severely inhibited in renal transplant patients on mycophenolate mofetil treatment. Transpl. Immunol. 2004, 13, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Rentenaar, R.J.; Van Diepen, F.N.; Meijer, R.T.; Surachno, S.; Wilmink, J.M.; Schellekens, P.T.A.; Pals, S.T.; van Lier, R.; Berge, I.J.T. Immune responsiveness in renal transplant recipients: Mycophenolic acid severely depresses humoral immunity in vivo. Kidney Int. 2002, 62, 319–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulley, W.R.; Visvanathan, K.; Hurt, A.C.; Brown, F.G.; Polkinghorne, K.R.; Mastorakos, T.; Lewicki, M.C.; Stuart, R.L.; Tan, S.-J.; Chean, R.; et al. Mycophenolate and lower graft function reduce the seroresponse of kidney transplant recipients to pandemic H1N1 vaccination. Kidney Int. 2012, 82, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Joshi, M.D.; Singhania, S.; Ramsey, K.H.; Murthy, A.K. Peptide Vaccine: Progress and Challenges. Vaccines 2014, 2, 515–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platten, M.; Bunse, L.; Wick, A.; Bunse, T.; Le Cornet, L.; Harting, I.; Sahm, F.; Sanghvi, K.; Tan, C.L.; Poschke, I.; et al. A vaccine targeting mutant IDH1 in newly diagnosed glioma. Nature 2021, 592, 463–468. [Google Scholar] [CrossRef]
- Schmitt, A.; Bechter, C.; Yao, J.; Goetz, M.; Maccari, B.; Schauwecker, P.; Wiesneth, M.; Schmitt, M. Cytomegalovirus vaccination of leukemia and lymphoma patients after allogeneic stem cell transplantation—Validation of a peptide vaccine. J. Immunol. Methods 2009, 343, 140–147. [Google Scholar] [CrossRef]
- Shugay, M.; Bagaev, D.V.; Zvyagin, I.V.; Vroomans, R.M.; Crawford, J.C.; Dolton, G.; A Komech, E.; Sycheva, A.L.; E Koneva, A.; Egorov, E.S.; et al. VDJdb: A curated database of T-cell receptor sequences with known antigen specificity. Nucleic Acids Res. 2017, 46, D419–D427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bunse, L.; Sommerer, C.; Tan, C.L.; Korell, F.; Schmitt, A.; Hückelhoven-Krauss, A.; Neuber, B.; Mertens, T.; Platten, M.; Green, E.W.; et al. Common T-Cell-Receptor Motifs and Features in Patients with Cytomegalovirus (CMV)-Seronegative End-Stage Renal Disease Receiving a Peptide Vaccination against CMV. Int. J. Mol. Sci. 2022, 23, 1029. https://doi.org/10.3390/ijms23031029
Bunse L, Sommerer C, Tan CL, Korell F, Schmitt A, Hückelhoven-Krauss A, Neuber B, Mertens T, Platten M, Green EW, et al. Common T-Cell-Receptor Motifs and Features in Patients with Cytomegalovirus (CMV)-Seronegative End-Stage Renal Disease Receiving a Peptide Vaccination against CMV. International Journal of Molecular Sciences. 2022; 23(3):1029. https://doi.org/10.3390/ijms23031029
Chicago/Turabian StyleBunse, Lukas, Claudia Sommerer, Chin Leng Tan, Felix Korell, Anita Schmitt, Angela Hückelhoven-Krauss, Brigitte Neuber, Thomas Mertens, Michael Platten, Edward W. Green, and et al. 2022. "Common T-Cell-Receptor Motifs and Features in Patients with Cytomegalovirus (CMV)-Seronegative End-Stage Renal Disease Receiving a Peptide Vaccination against CMV" International Journal of Molecular Sciences 23, no. 3: 1029. https://doi.org/10.3390/ijms23031029