
Hydrogen Sulfide-Linked Persulfidation Maintains Protein Stability of ABSCISIC ACID-INSENSITIVE 4 and Delays Seed Germination



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Inhibition of Germination and Seedling Growth in Arabidopsis by Exogenous H2S
2.2. DES1-Produced H2S Inhibits Arabidopsis Seed Germination and Post-Germination Growth
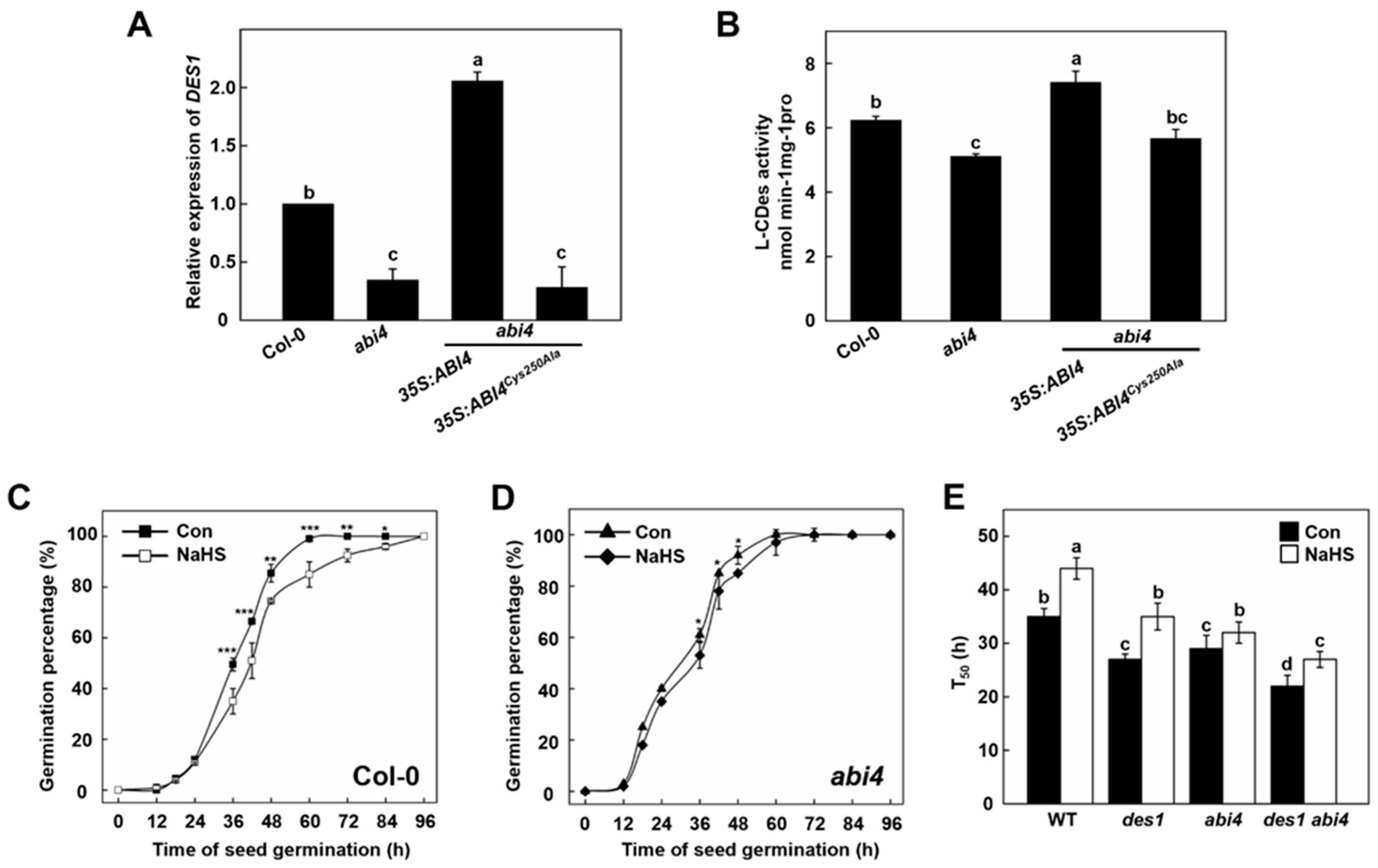
2.3. Functional of DES1 and ABI4 on Seed Germination and Post-Germination Growth
2.4. ABI4 Protein and Its Persulfidation Level Decreases during Germination and Post-Germination Stages
2.5. Persulfidation at Cys250 of ABI4 Inhibits Its Degradation
2.6. The Stability of ABI4 Is Regulated by Its Persulfidation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Molecular Cloning
4.2.1. For Expression in Escherichia coli
4.2.2. For Transient Expression in Arabidopsis Protoplasts
4.2.3. For Expression in Planta
4.3. Real-Time RT-PCR Analysis
4.4. Determination of Activity of L-Cysteine Desulfhydrase
4.5. Seed Germination and Green Open Cotyledon Assays
4.6. Expression and Purification of Recombinant Protein
4.7. SDS-PAGE and Immunoblotting
4.8. Immunochemical Detection of S-Persulfidated Proteins
4.9. Cell-Free Protein Degradation Assay
4.10. Protoplast-Based Protein Degradation Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frederick, D.D.; Nair, S.P.; Ward, P.D. Increased growth and germination success in plants following hydrogen sulfide administration. PLoS ONE 2013, 8, e62048. [Google Scholar]
- Laureano-Marín, A.M.; Aroca, Á.; Pérez-Pérez, M.E.; Yruela, I.; Jurado-Flores, A.; Moreno, I.; Gotor, C. Abscisic acid-triggered persulfidation of the cysteine protease ATG4 mediates regulation of autophagy by sulfide. Plant Cell 2020, 32, 3902–3920. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hu, S.L.; Zhang, Z.J.; Hu, L.Y.; Jiang, C.X.; Wei, Z.J.; Liu, J.; Wang, H.L.; Jiang, S.T. Hydrogen sulfide acts as a regulator of flower senescence in plants. Postharvest Biol. Technol. 2011, 60, 251–257. [Google Scholar] [CrossRef]
- Scuffi, D.; Álvarez, C.; Laspina, N.; Gotor, C.; Lamattina, L.; García-Mata, C. Hydrogen sulfide generated by L-cysteine desulfhydrase acts upstream of nitric oxide to modulate ABA-dependent stomatal closure. Plant Physiol. 2014, 166, 2065–2076. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Alfonseca, L.; Gotor, C.; Romero, L.C.; Garcıá, I. ß-cyanoalanine synthase action in root hair elongationis exerted at early steps of the root hair elongation pathway and is independent of direct cyanide inactivation of NADPH oxidase. Plant Cell Physiol. 2018, 59, 1072–1083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, M.J.; Zhou, H.; Zhao, D.D.; Gotor, C.; Romero, L.C.; Shen, J.; Ge, Z.L.; Zhang, Z.R.; Shen, W.B.; et al. Hydrogen sulfide (H2S), a signaling molecule in plant stress responses. J. Integr. Plant Biol. 2021, 63, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.; Calo, L.; Romero, L.C.; Garcia, I.; Gotor, C. An O-acetylserine(thiol)lyase homolog with L-cysteine desulfhydrase activity regulates cysteine homeostasis in Arabidopsis. Plant Physiol. 2010, 152, 656–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Z.P.; Xue, S.W.; Luo, Y.N.; Tian, B.H.; Fang, H.H.; Li, H.; Pei, Y.X. Hydrogen sulfide interacting with abscisic acid in stomatal regulation responses to drought stress in Arabidopsis. Plant Physiol. Biochem. 2013, 62, 41–46. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, M.J.; Ge, Z.L.; Shen, J.; Zhou, C.; Gotor, C.; Romero, L.C.; Duan, X.L.; Liu, X.; Wu, D.L.; et al. ABA-triggered guard cell L-cysteine desulfhydrase function and in situ H2S production contributes to heme oxygenase-modulated stomatal closure. Plant Cell Environ. 2020, 43, 624–636. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen sulfide signaling in plants: Emerging roles of protein persulfidation. Front. Plant Sci. 2018, 9, 1369. [Google Scholar] [CrossRef] [Green Version]
- Moseler, A.; Dhalleine, T.; Rouhier, N.; Couturier, J. Arabidopsis thaliana 3-mercaptopyruvate sulfurtransferases interact with and are protected by reducing systems. J. Biol. Chem. 2021, 296, 100429. [Google Scholar] [CrossRef]
- Arif, Y.; Hayat, S.; Yusuf, M.; Bajguz, A. Hydrogen sulfide: A versatile gaseous molecule in plants. Plant Physiol. Biochem. 2021, 158, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R.; Jovanović, V.M. More than just an intermediate: Hydrogen sulfide signalling in plants. J. Exp. Bot. 2017, 68, 4733–4736. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, J.; Zhou, M.J.; Zhou, H.; Cui, B.M.; Gotor, C.; Romero, L.C.; Fu, L.; Yang, J.; Foyer, C.H.; et al. Persulfidation-based modification of cysteine desulfhydrase and the NADPH oxidase RBOHD controls guard cell abscise acid signaling. Plant Cell 2020, 32, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Benito, J.M.; Gotor, C.; Romero, L.C. Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.J.; Zhang, J.; Shen, J.; Zhou, H.; Zhao, D.D.; Gotor, C.; Romero, L.C.; Fu, L.; Li, Z.M.; Yang, J.; et al. Hydrogen sulfide-linked persulfidation of ABSCISIC INSENSITIVE 4 controls Arabidopsis ABA responses through the transactivation of mitogen-activated protein kinase kinase kinase 18. Mol. Plant 2021, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Flores, A.; Romero, L.C.; Gotor, C. Label-free quantitative proteomic analysis of nitrogen starvation in Arabidopsis root reveals new aspects of H2S signaling by protein persulfidation. Antioxidants 2021, 10, 508. [Google Scholar] [CrossRef] [PubMed]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nambara, E.; Okamoto, M.; Tatematsu, K.; Yano, R.; Seo, M.; Kamiya, Y. Abscisic acid and the control of seed dormancy and germination. Seed Sci. Res. 2010, 20, 55–67. [Google Scholar] [CrossRef]
- Zhou, M.J.; Zhou, H.; Shen, J.; Zhang, Z.R.; Gotor, C.; Romero, L.C.; Yuan, X.X.; Xie, Y.J. H2S action in plant life cycle. Plant Growth Regul. 2021, 94, 1–9. [Google Scholar] [CrossRef]
- Chandrasekaran, U.; Luo, X.F.; Zhou, W.G.; Shu, K. Multifaceted signaling networks mediated by Abscisic Acid Insensitive 4. Plant Commun. 2020, 1, 10040. [Google Scholar] [CrossRef]
- Shu, K.; Zhang, H.W.; Wang, S.F.; Chen, M.L.; Wu, Y.R.; Tang, S.Y.; Liu, C.Y.; Feng, Y.Q.; Cao, X.F.; Xie, Q. ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis. PLoS Genet. 2013, 9, e1003577. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Chen, Q.; Wu, Y.R.; Liu, R.J.; Zhang, H.W.; Wang, P.F.; Li, Y.L.; Wang, S.F.; Tang, S.Y.; Liu, C.Y.; et al. ABI4 mediates antagonistic effects of abscisic acid and gibberellins at transcript and protein levels. Plant J. 2016, 85, 348–361. [Google Scholar] [CrossRef] [Green Version]
- Nott, A.; Jung, H.S.; Koussevitzky, S.; Chory, J. Plastid-to-nucleus retrograde signaling. Annu. Rev. Plant Biol. 2006, 57, 739–759. [Google Scholar] [CrossRef]
- Wind, J.J.; Peviani, A.; Snel, B.; Hanson, J.; Smeekens, S.C. ABI4: Versatile activator and repressor. Trends Plant Sci. 2012, 18, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, X.; Gong, Z.; Yang, S.; Shi, Y. ABI4 represses the expression of type-A ARRs to inhibit seed germination in Arabidopsis. Plant J. 2017, 89, 354–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroca, A.; Gotor, C.; Bassham, D.C.; Romero, L.C. Hydrogen sulfide: From a toxic molecule to a key molecule of cell life. Antioxidants 2020, 9, 621. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.F.; Dai, Y.J.; Zheng, C.; Yang, Y.Z.; Chen, W.; Wang, Q.C.; Chandrasekaran, U.; Du, J.B.; Liu, W.G.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes reactive oxygen species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef]
- Giraud, E.; Van Aken, O.; Ho, L.H.M.; Whelan, J. The transcription factor ABI4 is a regulator of mitochondrial retrograde expression of ALTERNATIVE OXIDASE1a. Plant Physiol. 2009, 150, 1286–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shkolnik-Inbar, D.; Bar-Zvi, D. Expression of ABSCISIC ACID INSENSITIVE 4 (ABI4) in developing Arabidopsis seedlings. Plant Signal. Behav. 2011, 6, 694–696. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.B.; Lin, B.Y.; Yang, X.Y.; Liu, L.J.; Xia, R.; Li, J.G.; Wu, Y.R.; Xie, Q. The UBC27-AIRP3 ubiquitination complex modulates ABA signaling by promoting the degradation of ABI1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 27694–27702. [Google Scholar] [CrossRef]
- Kong, L.Y.; Cheng, J.K.; Zhu, Y.J.; Ding, Y.L.; Meng, J.J.; Chen, Z.Z.; Xie, Q.; Guo, Y.; Li, J.G.; Yang, S.H.; et al. Degradation of the ABA co-receptor ABI1 by PUB12/13 U-box E3 ligases. Nat. Commun. 2015, 6, 8630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albertos, P.; Romero-Puertas, M.C.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Nambara, E.; Lorenzo, O. S-nitrosylation triggers ABI5 degradation to promote seed germination and seedling growth. Nat. Commun. 2015, 6, 8669. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.J.; MacLean, A.M.; Sugio, A.; Kuo, C.H.; Kuo, R.G.H.; Hogenhout, S.A. Parasitic modulation of host development by ubiquitin-independent protein degradation. Cell 2021, 184, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Söderman, E.M.; Brocard, I.M.; Lynch, T.J.; Finkelstein, R.R. Regulation and function of the Arabidopsis ABA-insensitive 4 gene in seed and abscisic acid response signaling networks. Plant Physiol. 2000, 124, 1752–1765. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.S.; Jia, H.L.; Wang, X.F.; Shi, C.; Wang, X.; Ma, P.Q.; Wang, J.; Ren, M.J.; Li, J.S. Hydrogen sulfide positively regulates abscisic acid signaling through persulfidation of SnRK2.6 in Guard Cells. Mol. Plant 2020, 13, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Wang, X.F.; Jia, H.L.; Li, F.L.; Ma, Y.; Liesche, J.; Liao, M.Z.; Ding, X.T.; Liu, C.X.; Chen, Y.; et al. Persulfidation-induced structural change in SnRK2.6 establishes intramolecular interaction between phosphorylation and persulfidation. Mol. Plant 2021, 14, 1814–1830. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Riemenschneider, A.; Nikiforova, V.; Hoefgen, R.; De Kok, L.J.; Papenbrock, J. Impact of elevated H2S on metabolite levels, activity of enzymes and expression of genes involved in cysteine metabolism. Plant Physiol. Biochem. 2005, 43, 473–483. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Zhang, J.; Zhou, H.; Zhao, D.; Duan, T.; Wang, S.; Yuan, X.; Xie, Y. Hydrogen Sulfide-Linked Persulfidation Maintains Protein Stability of ABSCISIC ACID-INSENSITIVE 4 and Delays Seed Germination. Int. J. Mol. Sci. 2022, 23, 1389. https://doi.org/10.3390/ijms23031389
Zhou M, Zhang J, Zhou H, Zhao D, Duan T, Wang S, Yuan X, Xie Y. Hydrogen Sulfide-Linked Persulfidation Maintains Protein Stability of ABSCISIC ACID-INSENSITIVE 4 and Delays Seed Germination. International Journal of Molecular Sciences. 2022; 23(3):1389. https://doi.org/10.3390/ijms23031389
Chicago/Turabian StyleZhou, Mingjian, Jing Zhang, Heng Zhou, Didi Zhao, Tianqi Duan, Shuhan Wang, Xingxing Yuan, and Yanjie Xie. 2022. "Hydrogen Sulfide-Linked Persulfidation Maintains Protein Stability of ABSCISIC ACID-INSENSITIVE 4 and Delays Seed Germination" International Journal of Molecular Sciences 23, no. 3: 1389. https://doi.org/10.3390/ijms23031389
APA StyleZhou, M., Zhang, J., Zhou, H., Zhao, D., Duan, T., Wang, S., Yuan, X., & Xie, Y. (2022). Hydrogen Sulfide-Linked Persulfidation Maintains Protein Stability of ABSCISIC ACID-INSENSITIVE 4 and Delays Seed Germination. International Journal of Molecular Sciences, 23(3), 1389. https://doi.org/10.3390/ijms23031389