
Exogenous Bioactive Peptides Have a Potential Therapeutic Role in Delaying Aging in Rodent Models


Abstract
:1. Introduction
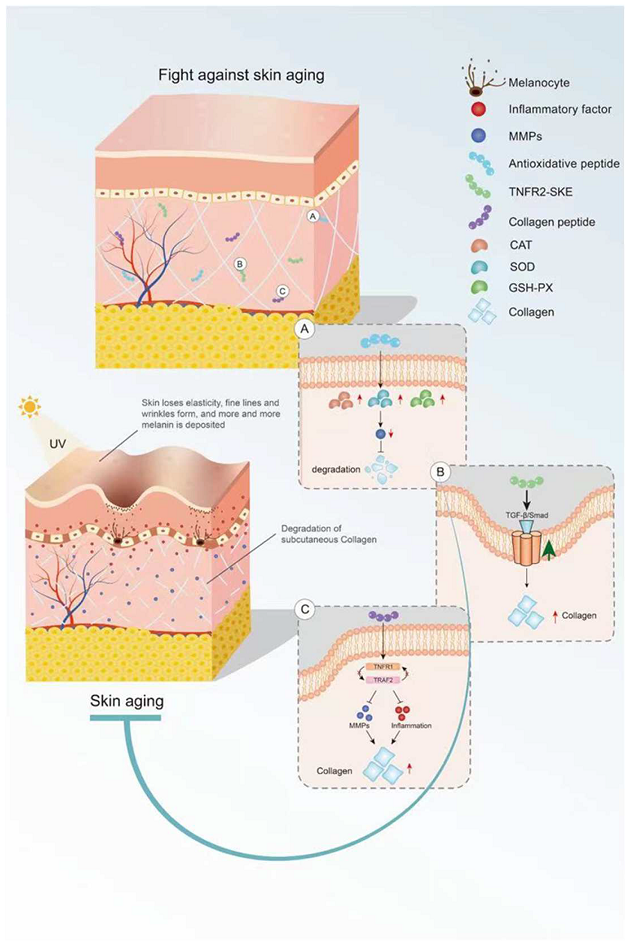
2. Bioactive Peptides Delay Skin Aging
2.1. Skin Aging
2.2. Antioxidant Peptides in Delaying Skin Aging
2.3. Anti-Inflammatory Peptides in Delaying Skin Aging
2.4. Peptides in Reducing Collagen Hydrolysis
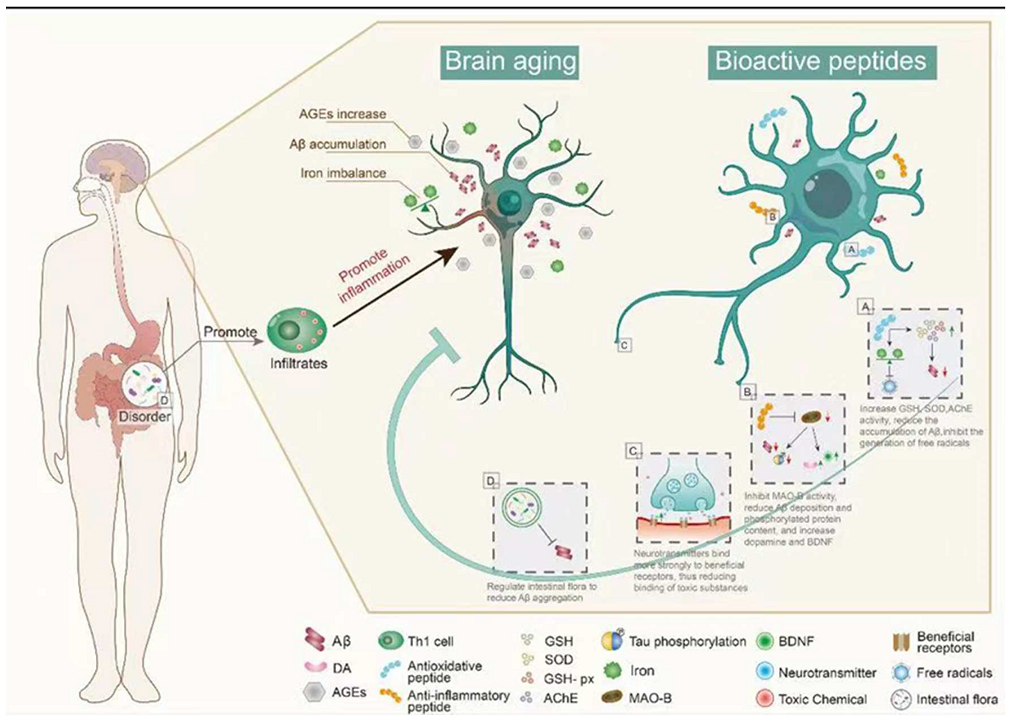
3. Bioactive Peptides and Brain Aging
3.1. Brain Aging
3.2. Antioxidant Peptides in Delaying Brain Aging
3.3. Anti-Inflammatory Peptide in Delaying Brain Aging
3.4. Regulation of Peptide Receptors in Delaying Brain Aging
3.5. Intestinal Microbiota Regulation by Peptides in Delaying Brain Aging
4. Bioactive Peptides and Aging in Other Organs
4.1. Lung Aging
4.1.1. Antioxidant Peptides in Delaying Lung Aging
4.1.2. Anti-Inflammatory Peptide in Delaying Lung Aging
4.2. Liver Aging
4.2.1. Antioxidant and Anti-Inflammatory Peptides in Delaying Liver Aging
4.2.2. Intestinal Microbiota Regulation by Peptides in Delaying Liver Aging
4.3. Kidney Aging
4.3.1. Antioxidant Peptides in Delaying Renal Aging
4.3.2. Anti-Inflammatory Peptides in Delaying Renal Aging
4.4. Aging of the Heart and Blood Vessels
4.4.1. Antioxidant Peptides in Delaying Cardiovascular Aging
4.4.2. Anti-Inflammatory Peptide in Delaying Cardiovascular Aging
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Abbreviations | Full Name | Abbreviations | Full Name |
|---|---|---|---|
| SOD | Superoxide dismutase | Samd | Drosophila mothers against decapentaplegic protein |
| GSH-Px | Glutathione peroxidase | pAMPK/AMP | AMP-activated protein kinase/AMP |
| GSH | Glutathione | pPKCζ/PKCζ | Anti-phospho-protein kinase ζ |
| CAT | Catalase | NOX4 | Antibodies against NADPH oxidase4 |
| Nrf2 | Transcription factor nuclear factor erythroid 2-related factor 2 | EPO | Erythropoietin |
| BDNF | Brain-derived neurotrophic factor | HIF-2α | Hypoxia-inducible factor |
| CREB | cAMP-response element-binding protein | NF-κB | Nuclear factor-kappa beta |
| AchE | Acetylcholinesterase | COX2 | Cyclooxygenase |
| MDA | Malondialdehyde | Ach | Acetylcholine |
| TNFα | Tumour necrosis factor-α | AchR | Cetylcholine receptor |
| IL-8 | Interleukin-8 | NPTX2 | Neuronal pentraxin 2 |
| IL-6 | Interleukin-6 | INMT | Indolethylamine N-methyltransferas |
| IL-1β | Interleukin-1β | HAL | Histamine ammonia-lyase |
| ATP | Adenosine triphosphate | MMP | Atrix metalloproteinase |
| PINK1 | Mutations in the PTEN-induced kinase 1 | ERK | Extracellular signal-regulated kinase |
| Parkin | Parkin RBR E3 ubiquitin protein ligase | HYP | Hydroxyproline |
| LC3Ⅱ/LC3Ⅰ | Microtubule-associated protein light chain 3 | p-CaMKII | Phosphorylated Ca2+/calmodulin-dependent protein kinase II |
| KEAP1 | Kelch-like ECH-associated protein 1 | PLN | Dephosphorylation of phospholamban |
| p62 | Protein sequestosome 1/p62 | ERK | Extracellular regulated protein kinases |
| pAKT | Phosphorylated protein kinase B | pGSK3β | Phosphorylated glycogen synthase kinase-3beta |
| Sirt1 | Silencing information regulator 2 related enzyme | 4-HNE | 4-hydroxynonenal |
| pAMPK | Phosphorylated AMP-activated protein kinase | FGF-2 | Fibroblast growth factor 2 |
| PGC1α | Peroxisome proliferator-activated receptor-γ co-activator-1α | SNAP-25 | Synaptosomal associated protein 25 |
| pFOXO3a | Phospho forkhead box O3a | PSD95 | Postsynaptic density proteins |
| GOT | Glutamic oxaloacetic transaminase | GluR1 | Anti-phospho-AMPARs |
| GPT | Glutamic-pyruvic transaminase | p-CREB | Phosphorylated cAMP-response element-binding protein |
| LDL | Low-density lipoprotein | GFAP | Astrocytosis |
| ANP | Atrial natriuretic peptide | p-IKKβ | Phosphorylated IKKbeta |
| BNP | Cerebral natriuretic peptide | NOS2 | Nitric oxide synthase-2 |
| pGATA4 | Phosphorylated GATA binding protein 4 | p-JNK | Hospho-c-JunN-terminal Kinase |
| p-p38/p38 | Phosphorylated p38 kinase/p38 kinase | TGFβ | Transforming growth factor-beta |
| GSN | Gelsolin |
References
- Partridge, L.; Fuentealba, M.; Kennedy, B.K. The quest to slow ageing through drug discovery. Nat. Rev. Drug Discov. 2020, 19, 513–532. [Google Scholar] [CrossRef] [PubMed]
- Kane, A.E.; Sinclair, D.A. Sirtuins and NAD+in the Development and Treatment of Metabolic and Cardiovascular Diseases. Circ. Res. 2018, 123, 868–885. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Mills, K.; Le Cessie, S.; Noordam, R.; Van Heemst, D. Ageing, age-related diseases and oxidative stress: What to do next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, J.P.; Vitorino, R.; Silva, G.M.; Vogel, C.; Duarte, A.C.; Rocha-Santos, T. A synopsis on aging-Theories, mechanisms and future prospects. Ageing Res. Rev. 2016, 29, 90–112. [Google Scholar] [CrossRef]
- Wagner, K.H.; Cameron-Smith, D.; Wessner, B.; Franzke, B. Biomarkers of Aging: From Function to Molecular Biology. Nutrients 2016, 8, 338. [Google Scholar] [CrossRef] [Green Version]
- Gurau, F.; Baldoni, S.; Prattichizzo, F.; Espinosa, E.; Amenta, F.; Procopio, A.D.; Albertini, M.C.; Bonafe, M.; Olivieri, F. Anti-senescence compounds: A potential nutraceutical approach to healthy aging. Ageing Res. Rev. 2018, 46, 14–31. [Google Scholar] [CrossRef]
- Kim, D.H.; Bang, E.; Jung, H.J.; Noh, S.G.; Yu, B.P.; Choi, Y.J.; Chung, H.Y. Anti-aging Effects of Calorie Restriction (CR) and CR Mimetics based on the Senoinflammation Concept. Nutrients 2020, 12, 422. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Gorguc, A.; Gencdag, E.; Yilmaz, F.M. Bioactive peptides derived from plant origin by-products: Biological activities and techno-functional utilizations in food developments—A review. Food Res. Int. 2020, 136, 109504. [Google Scholar] [CrossRef]
- Bechaux, J.; Gatellier, P.; Le Page, J.F.; Drillet, Y.; Sante-Lhoutellier, V. A comprehensive review of bioactive peptides obtained from animal byproducts and their applications. Food Funct. 2019, 10, 6244–6266. [Google Scholar] [CrossRef]
- Rutherfurd-Markwick, K.J. Food proteins as a source of bioactive peptides with diverse functions. Br. J. Nutr. 2012, 108 (Suppl. 2), S149–S157. [Google Scholar] [CrossRef] [Green Version]
- Azzu, V.; Valencak, T.G. Energy Metabolism and Ageing in the Mouse: A Mini-Review. Gerontology 2017, 63, 327–336. [Google Scholar] [CrossRef]
- Brunet, A. Old and new models for the study of human ageing. Nat. Rev. Mol. Cell. Biol. 2020, 21, 491–493. [Google Scholar] [CrossRef]
- Gomes, M.J.C.; Lima, S.L.S.; Alves, N.E.G.; Assis, A.; Moreira, M.E.C.; Toledo, R.C.L.; Rosa, C.O.B.; Teixeira, O.R.; Bassinello, P.Z.; De Mejia, E.G.; et al. Common bean protein hydrolysate modulates lipid metabolism and prevents endothelial dysfunction in BALB/c mice fed an atherogenic diet. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Su, G.; Zhang, X.; Song, G.; Zhang, L.; Zheng, L.; Zhao, M. Characterization and Exploration of Potential Neuroprotective Peptides in Walnut (Juglans regia) Protein Hydrolysate against Cholinergic System Damage and Oxidative Stress in Scopolamine-Induced Cognitive and Memory Impairment Mice and Zebrafish. J. Agric. Food Chem. 2021, 69, 2773–2783. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, L.; Zhao, T.; Zhang, Q.; Liu, Y.; Sun, B.; Su, G.; Zhao, M. Inhibitory Effects of Walnut (Juglans regia) Peptides on Neuroinflammation and Oxidative Stress in Lipopolysaccharide-Induced Cognitive Impairment Mice. J. Agric. Food Chem. 2020, 68, 2381–2392. [Google Scholar] [CrossRef]
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin barrier immunity and ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef]
- Lephart, E.D. Skin aging and oxidative stress: Equol’s anti-aging effects via biochemical and molecular mechanisms. Ageing Res. Rev. 2016, 31, 36–54. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Q.; Wang, L.; Xu, W.; He, Y.; Li, Y.; He, S.; Ma, H. Improvement of skin condition by oral administration of collagen hydrolysates in chronologically aged mice. J. Sci. Food Agric. 2017, 97, 2721–2726. [Google Scholar] [CrossRef]
- Aguirre-Cruz, G.; León-López, A.; Cruz-Gómez, V.; Jiménez-Alvarado, R.; Aguirre-Álvarez, G. Collagen Hydrolysates for Skin Protection: Oral Administration and Topical Formulation. Antioxidants 2020, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, X.; Bai, F.; Fang, Y.; Wang, J.; Gao, R. The anti-skin-aging effect of oral administration of gelatin from the swim bladder of Amur sturgeon (Acipenser schrenckii). Food Funct. 2019, 10, 3890–3897. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, Y.; Shi, Y.; Chen, R.; Fu, X.; Zhao, Y. Polypeptides extracted from Eupolyphaga sinensis walker via enzymic digestion alleviate UV radiation-induced skin photoaging. Biomed. Pharmacother. 2019, 112, 108636. [Google Scholar] [CrossRef]
- Lee, K.J.; Park, K.H.; Hahn, J.H. Alleviation of Ultraviolet-B Radiation-Induced Photoaging by a TNFR Antagonistic Peptide, TNFR2-SKE. Mol. Cells 2019, 42, 151–160. [Google Scholar] [CrossRef]
- Kim, K.H.; Son, J.M.; Benayoun, B.A.; Lee, C. The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress. Cell Metab. 2018, 28, 516–524.e7. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Zeng, J.; Drew, B.G.; Sallam, T.; Martin-Montalvo, A.; Wan, J.; Kim, S.J.; Mehta, H.; Hevener, A.L.; de Cabo, R.; et al. The mitochondrial-derived peptide MOTS-c promotes metabolic homeostasis and reduces obesity and insulin resistance. Cell Metab. 2015, 21, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Lu, H.; Hu, G.; Ye, Z.; Zhai, D.; Yan, Z.; Wang, L.; Xiang, A.; Lu, Z. Earlier changes in mice after D-galactose treatment were improved by mitochondria derived small peptide MOTS-c. Biochem. Biophys. Res. Commun. 2019, 513, 439–445. [Google Scholar] [CrossRef]
- Song, H.; Zhang, S.; Zhang, L.; Li, B. Effect of Orally Administered Collagen Peptides from Bovine Bone on Skin Aging in Chronologically Aged Mice. Nutrients 2017, 9, 1209. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, S.; Song, H.; Li, B. Ingestion of collagen hydrolysates alleviates skin chronological aging in an aged mouse model by increasing collagen synthesis. Food Funct. 2020, 11, 5573–5580. [Google Scholar] [CrossRef]
- Lee, H.J.; Jang, H.L.; Ahn, D.K.; Kim, H.J.; Jeon, H.Y.; Seo, D.B.; Lee, J.H.; Choi, J.K.; Kang, S.S. Orally administered collagen peptide protects against UVB-induced skin aging through the absorption of dipeptide forms, Gly-Pro and Pro-Hyp. Biosci. Biotechnol. Biochem. 2019, 83, 1146–1156. [Google Scholar] [CrossRef]
- Kang, M.C.; Yumnam, S.; Kim, S.Y. Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis. Int. J. Mol. Sci. 2018, 19, 3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Su, G.; Zhou, F.; Zhang, J.; Zheng, L.; Zhao, M. Protective Effect of Bovine Elastin Peptides against Photoaging in Mice and Identification of Novel Antiphotoaging Peptides. J. Agric. Food Chem. 2018, 66, 10760–10768. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Arumugam, T.V. Hallmarks of Brain Aging: Adaptive and Pathological Modification by Metabolic States. Cell Metab. 2018, 27, 1176–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Lalut, J.; Karila, D.; Dallemagne, P.; Rochais, C. Modulating 5-HT and 5-HT receptors in Alzheimer’s disease treatment. Future Med. Chem. 2017, 9, 781–795. [Google Scholar] [CrossRef]
- Hipkiss, A.R. Glycotoxins: Dietary and Metabolic Origins; Possible Amelioration of Neurotoxicity by Carnosine, with Special Reference to Parkinson’s Disease. Neurotox. Res. 2018, 34, 164–172. [Google Scholar] [CrossRef]
- Wang, X.; Sun, G.; Feng, T.; Zhang, J.; Huang, X.; Wang, T.; Xie, Z.; Chu, X.; Yang, J.; Wang, H.; et al. Sodium oligomannate therapeutically remodels gut microbiota and suppresses gut bacterial amino acids-shaped neuroinflammation to inhibit Alzheimer’s disease progression. Cell Res. 2019, 29, 787–803. [Google Scholar] [CrossRef]
- Banerjee, S.; Mukherjee, B.; Poddar, M.K.; Dunbar, G.L. Carnosine improves aging-induced cognitive impairment and brain regional neurodegeneration in relation to the neuropathological alterations in the secondary structure of amyloid beta (Abeta). J. Neurochem. 2021, 158, 710–723. [Google Scholar] [CrossRef]
- Aydin, A.F.; Coban, J.; Dogan-Ekici, I.; Betul-Kalaz, E.; Dogru-Abbasoglu, S.; Uysal, M. Carnosine and taurine treatments diminished brain oxidative stress and apoptosis in D-galactose aging model. Metab. Brain Dis. 2016, 31, 337–345. [Google Scholar] [CrossRef]
- Derry, P.J.; Hegde, M.L.; Jackson, G.R.; Kayed, R.; Tour, J.M.; Tsai, A.L.; Kent, T.A. Revisiting the intersection of amyloid, pathologically modified tau and iron in Alzheimer’s disease from a ferroptosis perspective. Prog. Neurobiol. 2020, 184, 101716. [Google Scholar] [CrossRef]
- Roberts, B.R.; Ryan, T.M.; Bush, A.I.; Masters, C.L.; Duce, J.A. The role of metallobiology and amyloid-beta peptides in Alzheimer’s disease. J. Neurochem. 2012, 120 (Suppl. 1), 149–166. [Google Scholar] [CrossRef]
- Zou, Z.; Cai, J.; Zhong, A.; Zhou, Y.; Wang, Z.; Wu, Z.; Yang, Y.; Li, X.; Cheng, X.; Tan, J.; et al. Using the synthesized peptide HAYED (5) to protect the brain against iron catalyzed radical attack in a naturally senescence Kunming mouse model. Free Radic. Biol. Med. 2019, 130, 458–470. [Google Scholar] [CrossRef]
- Zou, Z.; Shao, S.; Zou, R.; Qi, J.; Chen, L.; Zhang, H.; Shen, Q.; Yang, Y.; Ma, L.; Guo, R.; et al. Linking the low-density lipoprotein receptor-binding segment enables the therapeutic 5-YHEDA peptide to cross the blood-brain barrier and scavenge excess iron and radicals in the brain of senescent mice. Alzheimers Dement. 2019, 5, 717–731. [Google Scholar] [CrossRef]
- Cui, X.; Lin, Q.; Liang, Y. Plant-Derived Antioxidants Protect the Nervous System From Aging by Inhibiting Oxidative Stress. Front. Aging Neurosci. 2020, 12, 209. [Google Scholar] [CrossRef]
- Kelly, A.S.; Auerbach, P.; Barrientos-Perez, M.; Gies, I.; Hale, P.M.; Marcus, C.; Mastrandrea, L.D.; Prabhu, N.; Arslanian, S.; Investigators, N.N.T. A Randomized, Controlled Trial of Liraglutide for Adolescents with Obesity. N. Engl. J. Med. 2020, 382, 2117–2128. [Google Scholar] [CrossRef]
- Wicinski, M.; Socha, M.; Malinowski, B.; Wodkiewicz, E.; Walczak, M.; Gorski, K.; Slupski, M.; Pawlak-Osinska, K. Liraglutide and its Neuroprotective Properties-Focus on Possible Biochemical Mechanisms in Alzheimer’s Disease and Cerebral Ischemic Events. Int. J. Mol. Sci. 2019, 20, 1050. [Google Scholar] [CrossRef] [Green Version]
- Hansen, H.H.; Fabricius, K.; Barkholt, P.; Niehoff, M.L.; Morley, J.E.; Jelsing, J.; Pyke, C.; Knudsen, L.B.; Farr, S.A.; Vrang, N. The GLP-1 Receptor Agonist Liraglutide Improves Memory Function and Increases Hippocampal CA1 Neuronal Numbers in a Senescence-Accelerated Mouse Model of Alzheimer’s Disease. J. Alzheimers Dis. 2015, 46, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Pang, M.; Yu, Y.; Gou, X.; Si, P.; Zhawatibai, A.; Zhang, Y.; Zhang, M.; Guo, T.; Yi, X.; et al. The neuroprotection of liraglutide on diabetic cognitive deficits is associated with improved hippocampal synapses and inhibited neuronal apoptosis. Life Sci. 2019, 231, 116566. [Google Scholar] [CrossRef]
- Ano, Y.; Yoshino, Y.; Uchida, K.; Nakayama, H. Preventive Effects of Tryptophan-Methionine Dipeptide on Neural Inflammation and Alzheimer’s Pathology. Int. J. Mol. Sci. 2019, 20, 3206. [Google Scholar] [CrossRef] [Green Version]
- Ano, Y.; Ayabe, T.; Ohya, R.; Kondo, K.; Kitaoka, S.; Furuyashiki, T. Tryptophan-Tyrosine Dipeptide, the Core Sequence of beta-Lactolin, Improves Memory by Modulating the Dopamine System. Nutrients 2019, 11, 348. [Google Scholar] [CrossRef] [Green Version]
- Ano, Y.; Yoshino, Y.; Kutsukake, T.; Ohya, R.; Fukuda, T.; Uchida, K.; Takashima, A.; Nakayama, H. Tryptophan-related dipeptides in fermented dairy products suppress microglial activation and prevent cognitive decline. Aging 2019, 11, 2949–2967. [Google Scholar] [CrossRef]
- Ano, Y.; Ayabe, T.; Kutsukake, T.; Ohya, R.; Takaichi, Y.; Uchida, S.; Yamada, K.; Uchida, K.; Takashima, A.; Nakayama, H. Novel lactopeptides in fermented dairy products improve memory function and cognitive decline. Neurobiol. Aging 2018, 72, 23–31. [Google Scholar] [CrossRef]
- Ano, Y.; Ohya, R.; Takaichi, Y.; Washinuma, T.; Uchida, K.; Takashima, A.; Nakayama, H. beta-Lactolin, a Whey-Derived Lacto-Tetrapeptide, Prevents Alzheimer’s Disease Pathologies and Cognitive Decline. J. Alzheimers Dis. 2020, 73, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- Kowianski, P.; Lietzau, G.; Czuba, E.; Waskow, M.; Steliga, A.; Morys, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Banerjee, S.; Ghosh, T.K.; Poddar, M.K. Carnosine reverses the aging-induced down regulation of brain regional serotonergic system. Mech. Ageing Dev. 2015, 152, 5–14. [Google Scholar] [CrossRef]
- Banerjee, S.; Poddar, M.K. Aging-induced changes in brain regional serotonin receptor binding: Effect of Carnosine. Neuroscience 2016, 319, 79–91. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Reglodi, D.; Atlasz, T.; Szabo, E.; Jungling, A.; Tamas, A.; Juhasz, T.; Fulop, B.D.; Bardosi, A. PACAP deficiency as a model of aging. Geroscience 2018, 40, 437–452. [Google Scholar] [CrossRef]
- Metz, V.V.; Kojro, E.; Rat, D.; Postina, R. Induction of RAGE shedding by activation of G protein-coupled receptors. PLoS ONE 2012, 7, e41823. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Bekhit, A.E.-D.A.; Wu, Q.; Chen, M.; Liao, X.; Wang, J.; Ding, Y. Bioactive peptides and gut microbiota: Candidates for a novel strategy for reduction and control of neurodegenerative diseases. Trends Food Sci. Technol. 2021, 108, 164–176. [Google Scholar] [CrossRef]
- Wang, M.; Amakye, W.K.; Guo, L.; Gong, C.; Zhao, Y.; Yao, M.; Ren, J. Walnut-Derived Peptide PW5 Ameliorates Cognitive Impairments and Alters Gut Microbiota in APP/PS1 Transgenic Mice. Mol. Nutr. Food Res. 2019, 63, e1900326. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.J.; Stout-Delgado, H.W. Aging and Lung Disease. Annu. Rev. Physiol. 2020, 82, 433–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Zhang, Y. Targeting Autophagy in Aging and Aging-Related Cardiovascular Diseases. Trends Pharmacol. Sci. 2018, 39, 1064–1076. [Google Scholar] [CrossRef]
- Easter, M.; Bollenbecker, S.; Barnes, J.W.; Krick, S. Targeting Aging Pathways in Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2020, 21, 6924. [Google Scholar] [CrossRef]
- Barnes, P.J.; Baker, J.; Donnelly, L.E. Cellular Senescence as a Mechanism and Target in Chronic Lung Diseases. Am. J. Respir. Crit. Care Med. 2019, 200, 556–564. [Google Scholar] [CrossRef]
- Chan, S.M.H.; Selemidis, S.; Bozinovski, S.; Vlahos, R. Pathobiological mechanisms underlying metabolic syndrome (MetS) in chronic obstructive pulmonary disease (COPD): Clinical significance and therapeutic strategies. Pharmacol. Ther. 2019, 198, 160–188. [Google Scholar] [CrossRef]
- Lonardo, A.; Nascimbeni, F.; Ponz de Leon, M. Nonalcoholic fatty liver disease and COPD: Is it time to cross the diaphragm? Eur. Respir J. 2017, 49, 1700546. [Google Scholar] [CrossRef] [Green Version]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. The human tripeptide GHK-Cu in prevention of oxidative stress and degenerative conditions of aging: Implications for cognitive health. Oxid. Med. Cell Longev. 2012, 2012, 324832. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-R.; Lee, H.; Kim, S.-I.; Yang, S.-R. The tri-peptide GHK-Cu complex ameliorates lipopolysaccharide-induced acute lung injury in mice. Oncotarget 2016, 7, 58405–58417. [Google Scholar] [CrossRef]
- Meiners, S.; Eickelberg, O. Next-generation personalized drug discovery: The tripeptide GHK hits center stage in chronic obstructive pulmonary disease. Genome Med. 2012, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Yang, H.; Lin, S.D.; Zhao, L.; Jiang, C.; Chen, Z.B.; Liu, Y.Y.; Kan, Y.J.; Hu, J.; Pang, W.S. Cyclic Peptide Extracts Derived From Pseudostellaria heterophylla Ameliorates COPD via Regulation of the TLR4/MyD88 Pathway Proteins. Front. Pharmacol. 2020, 11, 850. [Google Scholar] [CrossRef]
- Estes, C.; Razavi, H.; Loomba, R.; Younossi, Z.; Sanyal, A.J. Modeling the epidemic of nonalcoholic fatty liver disease demonstrates an exponential increase in burden of disease. Hepatology 2018, 67, 123–133. [Google Scholar] [CrossRef]
- Niemann, J.; Johne, C.; Schroder, S.; Koch, F.; Ibrahim, S.M.; Schultz, J.; Tiedge, M.; Baltrusch, S. An mtDNA mutation accelerates liver aging by interfering with the ROS response and mitochondrial life cycle. Free Radic. Biol. Med. 2017, 102, 174–187. [Google Scholar] [CrossRef]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef]
- Tilg, H.; Zmora, N.; Adolph, T.E.; Elinav, E. The intestinal microbiota fuelling metabolic inflammation. Nat. Rev. Immunol. 2020, 20, 40–54. [Google Scholar] [CrossRef]
- Lonardo, A.; Nascimbeni, F.; Mantovani, A.; Targher, G. Hypertension, diabetes, atherosclerosis and NASH: Cause or consequence? J. Hepatol. 2018, 68, 335–352. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Tabibian, J.H.; Ekstedt, M.; Kechagias, S.; Hamaguchi, M.; Hultcrantz, R.; Hagstrom, H.; Yoon, S.K.; Charatcharoenwitthaya, P.; et al. Association of non-alcoholic fatty liver disease with chronic kidney disease: A systematic review and meta-analysis. PLoS Med. 2014, 11, e1001680. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xue, H.; Fang, W.; Chen, K.; Chen, S.; Yang, W.; Shen, T.; Chen, X.; Zhang, P.; Ling, W. Adropin protects against liver injury in nonalcoholic steatohepatitis via the Nrf2 mediated antioxidant capacity. Redox. Biol. 2019, 21, 101068. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, X.; Lin, Q.; Cai, J.; Tang, L.; Liang, Y. Active Peptide KF-8 from Rice Bran Attenuates Oxidative Stress in a Mouse Model of Aging Induced by d-Galactose. J. Agric. Food Chem. 2020, 68, 12271–12283. [Google Scholar] [CrossRef]
- Moreira, G.V.; Azevedo, F.F.; Ribeiro, L.M.; Santos, A.; Guadagnini, D.; Gama, P.; Liberti, E.A.; Saad, M.; Carvalho, C. Liraglutide modulates gut microbiota and reduces NAFLD in obese mice. J. Nutr. Biochem. 2018, 62, 143–154. [Google Scholar] [CrossRef]
- Choudhury, D.; Levi, M. Kidney aging—Inevitable or preventable? Nat. Rev. Nephrol. 2011, 7, 706–717. [Google Scholar] [CrossRef]
- Kimura, T.; Isaka, Y.; Yoshimori, T. Autophagy and kidney inflammation. Autophagy 2017, 13, 997–1003. [Google Scholar] [CrossRef]
- Gansevoort, R.T.; Correa-Rotter, R.; Hemmelgarn, B.R.; Jafar, T.H.; Heerspink, H.J.L.; Mann, J.F.; Matsushita, K.; Wen, C.P. Chronic kidney disease and cardiovascular risk: Epidemiology, mechanisms, and prevention. Lancet 2013, 382, 339–352. [Google Scholar] [CrossRef]
- Sun, L.; Xu, H.; Wang, Y.; Ma, X.; Xu, Y.; Sun, F. The mitochondrial-targeted peptide SBT-20 ameliorates inflammation and oxidative stress in chronic renal failure. Aging 2020, 12, 18238–18250. [Google Scholar] [CrossRef]
- Yang, H.-Y.; Chen, J.-R.; Chang, L.-S. Effects of soy protein hydrolysate on blood pressure and angiotensin-converting enzyme activity in rats with chronic renal failure. Hypertens. Res. 2008, 31, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Paneni, F.; Diaz Canestro, C.; Libby, P.; Luscher, T.F.; Camici, G.G. The Aging Cardiovascular System: Understanding It at the Cellular and Clinical Levels. J. Am. Coll. Cardiol. 2017, 69, 1952–1967. [Google Scholar] [CrossRef] [PubMed]
- Donato, A.J.; Machin, D.R.; Lesniewski, L.A. Mechanisms of Dysfunction in the Aging Vasculature and Role in Age-Related Disease. Circ. Res. 2018, 123, 825–848. [Google Scholar] [CrossRef]
- Camare, C.; Pucelle, M.; Negre-Salvayre, A.; Salvayre, R. Angiogenesis in the atherosclerotic plaque. Redox. Biol. 2017, 12, 18–34. [Google Scholar] [CrossRef]
- Gong, X.; An, Q.; Le, L.; Geng, F.; Jiang, L.; Yan, J.; Xiang, D.; Peng, L.; Zou, L.; Zhao, G.; et al. Prospects of cereal protein-derived bioactive peptides: Sources, bioactivities diversity, and production. Crit. Rev. Food Sci. Nutr. 2020, 1–17. [Google Scholar] [CrossRef]
- Tong, L.-T.; Ju, Z.; Wang, L.; Qiu, J.; Liu, L.; Zhou, X.; Liang, T.; Geng, D.; Zhou, S. Peptides derived from rice α-globulin reduce atherosclerosis in apolipoprotein E-deficient mice by inhibiting TNF-α-induced vascular endothelial cells injury. J. Funct. Foods 2019, 63, 103582. [Google Scholar] [CrossRef]
- Ktari, N.; Bkhairia, I.; Nasri, R.; Ben Abdallah Kolsi, R.; Ben Slama-Ben Salem, R.; Ben Amara, I.; Zeghal, N.; Ben Salah, B.; Ben Salah, R.; Nasri, M. Zebra blenny protein hydrolysates as a source of bioactive peptides with prevention effect against oxidative dysfunctions and DNA damage in heart tissues of rats fed a cholesterol-rich diet. Food Res. Int. 2017, 100, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Volpe, M. Natriuretic Peptides in the Cardiovascular System: Multifaceted Roles in Physiology, Pathology and Therapeutics. Int. J. Mol. Sci. 2019, 20, 3991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Rignault-Clerc, S.; Bielmann, C.; Bon-Mathier, A.C.; Deglise, T.; Carboni, A.; Ducrest, M.; Rosenblatt-Velin, N. Increasing heart vascularisation after myocardial infarction using brain natriuretic peptide stimulation of endothelial and WT1(+) epicardial cells. Elife 2020, 9, e61050. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yamashita, T.; Shirai, R.; Shibata, K.; Okano, T.; Yamaguchi, M.; Mori, Y.; Hirano, T.; Watanabe, T. Adropin Contributes to Anti-Atherosclerosis by Suppressing Monocyte-Endothelial Cell Adhesion and Smooth Muscle Cell Proliferation. Int. J. Mol. Sci. 2018, 19, 1293. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Liu, C.; Fang, L.; Lu, H.; Wang, J.; Gao, Y.; Gabbianelli, R.; Min, W. Walnut-Derived Peptide Activates PINK1 via the NRF2/KEAP1/HO-1 Pathway, Promotes Mitophagy, and Alleviates Learning and Memory Impairments in a Mice Model. J. Agric. Food Chem. 2021, 69, 2758–2772. [Google Scholar] [CrossRef]
- Asokan, S.M.; Wang, T.; Wang, M.F.; Lin, W.T. A novel dipeptide from potato protein hydrolysate augments the effects of exercise training against high-fat diet-induced damages in senescence-accelerated mouse-prone 8 by boosting pAMPK/SIRT1/PGC-1α/pFOXO3 pathway. Aging 2020, 12, 7334–7349. [Google Scholar] [CrossRef]
- Hu, W.S.; Ting, W.J.; Tamilselvi, S.; Day, C.H.; Wang, T.; Chiang, W.D.; Viswanadha, V.P.; Yeh, Y.L.; Lin, W.T.; Huang, C.Y. Oral administration of alcalase potato protein hydrolysate-APPH attenuates high fat diet-induced cardiac complications via TGF-beta/GSN axis in aging rats. Environ. Toxicol. 2019, 34, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zheng, L.; Su, G.; Zeng, X.A.; Sun, B.; Zhao, M. Evaluation and Exploration of Potentially Bioactive Peptides in Casein Hydrolysates against Liver Oxidative Damage in STZ/HFD-Induced Diabetic Rats. J. Agric. Food Chem. 2020, 68, 2393–2405. [Google Scholar] [CrossRef]
- Wang, F.; Weng, Z.; Lyu, Y.; Bao, Y.; Liu, J.; Zhang, Y.; Sui, X.; Fang, Y.; Tang, X.; Shen, X. Wheat germ-derived peptide ADWGGPLPH abolishes high glucose-induced oxidative stress via modulation of the PKCzeta/AMPK/NOX4 pathway. Food Funct. 2020, 11, 6843–6854. [Google Scholar] [CrossRef]
- Zhu, S.; Wu, L.; Zhang, J.; Miao, Y.; Zhao, Y.; Zeng, M.; Li, D.; Wu, H. Collagen Hydrolysate Corrects Anemia in Chronic Kidney Disease via Anti-Inflammatory Renoprotection and HIF-2alpha-Dependent Erythropoietin and Hepcidin Regulation. J. Agric. Food Chem. 2020, 68, 11726–11734. [Google Scholar] [CrossRef]
- Zhao, T.; Zheng, L.; Zhang, Q.; Wang, S.; Zhao, Q.; Su, G.; Zhao, M. Stability towards the gastrointestinal simulated digestion and bioactivity of PAYCS and its digestive product PAY with cognitive improving properties. Food Funct. 2019, 10, 2439–2449. [Google Scholar] [CrossRef]
- Kozaczek, M.; Bottje, W.; Greene, E.; Lassiter, K.; Kong, B.; Dridi, S.; Korourian, S.; Hakkak, R. Comparison of liver gene expression by RNAseq and PCR analysis after 8 weeks of feeding soy protein isolate- or casein-based diets in an obese liver steatosis rat model. Food Funct. 2019, 10, 8218–8229. [Google Scholar] [CrossRef]
- Xu, D.; Li, D.; Zhao, Z.; Wu, J.; Zhao, M. Regulation by walnut protein hydrolysate on the components and structural degradation of photoaged skin in SD rats. Food Funct. 2019, 2019, 6792–6802. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Z.; Huang, H.; Wang, Y.; Lin, B.; Wu, S.; Ma, J.; Chen, B.; He, Z.; Wu, J.; et al. Inhibition of bleomycin-induced pulmonary fibrosis in mice by the novel peptide EZY-1 purified from Eucheuma. Food Funct. 2019, 10, 3198–3208. [Google Scholar] [CrossRef]
- Ge, H.; Cai, Z.; Chai, J.; Liu, J.; Liu, B.; Yu, Y.; Liu, J.; Zhang, T. Egg white peptides ameliorate dextran sulfate sodium-induced acute colitis symptoms by inhibiting the production of pro-inflammatory cytokines and modulation of gut microbiota composition. Food Chem. 2021, 360, 129981. [Google Scholar] [CrossRef]
- Yu, X.C.; Li, Z.; Liu, X.R.; Hu, J.N.; Liu, R.; Zhu, N.; Li, Y. The Antioxidant Effects of Whey Protein Peptide on Learning and Memory Improvement in Aging Mice Models. Nutrients 2021, 13, 2100. [Google Scholar] [CrossRef]
- Gomez-Mendoza, D.P.; Lemos, R.P.; Jesus, I.C.G.; Gorshkov, V.; McKinnie, S.M.K.; Vederas, J.C.; Kjeldsen, F.; Guatimosim, S.; Santos, R.A.; Pimenta, A.M.C.; et al. Moving Pieces in a Cellular Puzzle: A Cryptic Peptide from the Scorpion Toxin Ts14 Activates AKT and ERK Signaling and Decreases Cardiac Myocyte Contractility via Dephosphorylation of Phospholamban. J. Proteome Res. 2020, 19, 3467–3477. [Google Scholar] [CrossRef]
- Qin, Q.; Mehta, H.; Yen, K.; Navarrete, G.; Brandhorst, S.; Wan, J.; Delrio, S.; Zhang, X.; Lerman, L.O.; Cohen, P.; et al. Chronic treatment with the mitochondrial peptide humanin prevents age-related myocardial fibrosis in mice. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1127–H1136. [Google Scholar] [CrossRef] [Green Version]
- Ali, T.; Badshah, H.; Kim, T.H.; Kim, M.O. Melatonin attenuates D-galactose-induced memory impairment, neuroinflammation and neurodegeneration via RAGE/NF-K B/JNK signaling pathway in aging mouse model. J. Pineal Res. 2015, 58, 71–85. [Google Scholar] [CrossRef]


| Classification | Name and Delivery Way | Source | Rodent Model | Target Organ | Mechanism |
|---|---|---|---|---|---|
| Food-derived active peptide | Walnut protein hydrolysates(WPH) Oral gavage for 21 days Low: 333 mg/kg High: 666 mg/kg | Walnut | Alzheimer’s disease model mice aged 6–8 weeks scopolamine solution (1.0 mg/kg) | Brain | SOD↑ GSH-Px↑ CAT↑ Nrf2↑ BDNF↑ CREB↑ MDA↓ TNFα↓ AchE↓ Trp-, Tyr-, or Phe-containing peptide has high affinity to Keap1 and Ache, so it can increase the activity of NRF2 and reduce the activity of Ache, which ultimately increases antioxidant capacity and anti-inflammatory ability and leads to increased BDNF, CREB transcription [15] |
| Walnut protein hydrolysate and its low-molecular-weight fraction (WPH/WPHL) Oral gavage for 21 daysWPH: 666 mg/kgWPHL: 666 mg/kg | Walnut | Alzheimer’s disease model mice aged 6–8 weeks LPS (300 μg/kg bw) | Brain | SOD↑ GSH-Px↑ CAT↑ MDA↓ TNFα↓ TNFα↓ IL-6↓ IL-1β↓ Trp, Gly, Leu residues, hydrophobic amino acids, and aromatic amino acids in polypeptides can inhibit the expression of pro-inflammatory factors TNF-α, IL-1β, and IL-6 and reduce inflammation [16] | |
| Tyr-Val-Leu-Leu-Pro-Ser-Pro-Ly (walnut protein hydrolysates) Continuous injection for 4 weeks 60 mg/kg bw | Walnut | Alzheimer’s disease model mice (C57BL/6) 5–6 week oldscopolamine solution (1 mg/kg bw) | Brain | ATP↑ PINK1↑ Parkin↑ NRF2↑ LC3 II/LC3 I↑ Beclin↑ KEAP1↓ p62↓ It can increase antioxidant capacity through Nrf2 signaling pathway and increase the expression of Beclin-1, Parkin, and PINK1 to enhance mitochondrial autophagy capacity [96] | |
| Alcalase potato-protein hydrolysates (IF) Oral administration 3 weeks 1 mg/kg bw | Potato | Senescence-Accelerated mice (SAMP8) 6 months high-fat diet | Liver/heart | pAKT↑ Sirt1↑ pAMPK↑ PGC1α↑ pFOXO3a↑ Bax↓ GOT↓ GPT↓ LDL↓ ANP↓ BNP↓ pGATA4↓ It can down-regulate cardiac hypertrophy markers ANP and BNP, reduce inflammation in the heart and liver, and reduce apoptosis by stimulating the activity of Sirt1 [97] | |
| Alcalase potato protein hydrolysate (APPH) Oral administration 4 weeks Low: 15 mg/kg/day Middle: 45 mg/kg/day High: 75 mg/kg/day | Potato | Sprague-Dawley (SD) rat 23 months old high-fat diet | Heart | p-p38/p38↓ GSN↓ p-Gata4↓ TGFβ↓ APPH has good lipid solubility and can reduce myocardial hypertrophy and fibrosis in aging rats through TGF-β/GSN pathway [98] | |
| Casein hydrolysates Continuous injection 10 weeks 200 mg/kg | Casein | Diabetic rat high-fat diet | Liver | NRF2↑ HO-1↑ SOD↑ GSH↑ MDA↓ By enhancing Nrf2 translation, the activity of antioxidant enzymes was enhanced, and the activities of DPP-IV and ACE were inhibited, among which dipeptide WM could inhibit Keap1/Nrf2 interaction [99] | |
| Wheat germ albumin hydrolysates ((Ala-Asp-Trp-Gly-Gly-Pro-Leu-Pro-His)) Continuous injection 1 week 4 mg/kg | Wheat | Diabetic mice 6 weeks old | Vascular | pAMPK/AMP↑ pPKCζ/PKCζ↓ NOX4↓ ROS↓ pAKT/AKT↓ Inhibition of NOX4 expression through the PKCζ/AMPK signaling pathway reduced oxidative stress levels and the release of inflammatory factors [100] | |
| Collagen hydrolysate Pro-Hyp Oral administration 4 weeks 210 mg/kg | Porcine skin | Chronic kidney disease mice 6 weeks old | Kidney | Liver iron content↑ EPO↑ HIF-2α↑ Hepcidin↓ TNF-α↓ IL-1β↓ IL-6↓ NF-κB↓ COX2↓ It reduces inflammation by regulating inflammatory pathways and plays a protective role in regulating HIF-2α, EPO, and Hepcidin [101] | |
| Anchovy hydrolysates Pro-Ala-Tyr-Cys-Ser (PAYCS) 20 days 0.2 mM/kg/day | Anchovy | Alzheimer’s disease model mice 6 weeks old Scopolamine solution (1 mg/kg bw) | Brain | Ach↑ AchR↑ Nrf2↑ BDNF↑ SOD↑ The antioxidative effects of PAYCS and PAY may be related to the Try active phenolic structure in the sequence and the hydrogen donor of the sulfhydryl group in Cys. Both active peptides have the ability to promote the binding of Ach and AchR [102] | |
| Soy protein isolate (SPI) Oral administration 8 weeks | Soy | Obese rat 6 weeks old | Liver | NPTX2↑ GPT↑ INMT↑ HAL↑ The increased expression of NPTX2 reduced the inflammation of the rat liver, the increased expression of GPT may be related to mitochondrial energy metabolism, the increased expression of INMT may be related to the relief of NAFLD symptoms, and the increased expression of HLT can consume excess protein in the liver [103] | |
| Walnut protein hydrolysate Oral administration Low: 0.32 g/L Middle: 0.96 g/L High: 2.88 g/L | Walnut | Skin-aging model rat Exposed to UV-R | Skin | Elastin↑ Fibrillin-1↑ MMP-1↓ Increasing the expression of Col I, Col III, HYP, and HA and significantly attenuated the activity of MMP-1 [104] | |
| Eucheuma hydrolysate (EZY-1) 28 day 0.25 mg/kg 0.5 mg/kg 1 mg/kg 50 mg/kg | Eucheuma | Pulmonary fibrosis mice (C57BL/6J) 8 weeks old injected with 3.5 mg/kg of bleomycin | Lung | T-SOD↑ GSH-Px↑ HYP↓ MDA↓ pSmad3↓ EZY- 1 is easily absorbed in the intestinal tract, and its hydrophobic point facilitates the entry of EZY-1 into cells, while EZY-1 can reduce pulmonary fibrosis through TGF-β/Samd signaling pathway. [105] | |
| Egg white protein hydrolysate (EWPs) Gavage 14 days Low: 50 mg/kg Middle: 100 mg/kg High: 200 mg/kg | Egg | Colitis model mice (BALB/c) administered 3% (w/v) DSS | Gut | Candidatus_Sacchar-imonas↑ norank_f_Ruminococcaceae↓ Ruminiclostridium↓ TNF-α↓ IL-6↓ IL-8↓ EWPs contain Trp, Try, His, and Met, which make it have good antioxidant activity and can reduce the release of inflammatory factors by increasing the content of Lactobacillus and Candidatus-Saccharimonas in the gut [106] | |
| Whey protein hydrolysate (WHP) Gavage 30 days Low: 0.3 g/kg Middle: 1.5 g/kg High: 3.0 g/kg | Egg | D-galactose-treated mice (C57BL/6N) 6 months 100 mg/kg | Brain | SOD↑ GSH-Px↑ AChE↑ p-CaMKII↑ MDA↓ TNF-α↓ IL-1β↓ TNF-α↓ WHP can reduce the release of inflammatory factors, increase the activity of antioxidant enzymes, and enhance the activities of AchE and P-CamKII, which play an important role in maintaining synaptic plasticity [107] | |
| A peptide encrypted from the venom of Tityus serrulatus scorpion | Lys-Pro-Pro (KPP) | Scorpion | Mice 10 weeks old | Heart | pPLN/PLN↓ pERK/ERK↓ KPP regulates cellular stress-related proteins and exerts cardioprotective effects through PLN dephosphorylation [108] |
| Secretory bioactive peptide | Humanin (HNG) Injections 14 months 4 mg/kg | Mitochondria | Aging mice (C57BL/6N) 18 months | Heart | pAKT↑ pGSK3β↓ 4-HNE↓ TGF-β1↓ FGF-2↓ MMP-2↓ HNG down-regulated the expression of GSK-3β through Akt pathway, reduced myocardial apoptosis, down-regulated FGF-2 and MMP-2 expression, and inhibited cardiac fibrosis [109] |
| Peptide hormone | Melatonin Injections 30 days 10 mg/kg | Pineal gland | Aging mice 8 weeks old D-galactose 100 mg/kg | Brain | SNAP-25↑ PSD95↑ GluR1↑ p-CREB↑ ROS↓ GFAP↓ p-IKKβ↓ NF-κB↓ COX-2↓ NOS2↓ IL-1β↓ TNFα↓ p-JNK↓ Melatonin can reduce synaptic damage caused by oxidative stress and neuroinflammation through RAGE/NFκB/JNK pathway and has a good therapeutic effect on neurodegeneration [110] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Wu, Y.; Chen, Z.; Chen, Y.; Lin, Q.; Liang, Y. Exogenous Bioactive Peptides Have a Potential Therapeutic Role in Delaying Aging in Rodent Models. Int. J. Mol. Sci. 2022, 23, 1421. https://doi.org/10.3390/ijms23031421
Wang J, Wu Y, Chen Z, Chen Y, Lin Q, Liang Y. Exogenous Bioactive Peptides Have a Potential Therapeutic Role in Delaying Aging in Rodent Models. International Journal of Molecular Sciences. 2022; 23(3):1421. https://doi.org/10.3390/ijms23031421
Chicago/Turabian StyleWang, Jianqiang, Yixin Wu, Zhongxu Chen, Yajuan Chen, Qinlu Lin, and Ying Liang. 2022. "Exogenous Bioactive Peptides Have a Potential Therapeutic Role in Delaying Aging in Rodent Models" International Journal of Molecular Sciences 23, no. 3: 1421. https://doi.org/10.3390/ijms23031421