
Anti-Thrombotic Effects of Artesunate through Regulation of cAMP and PI3K/MAPK Pathway on Human Platelets


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
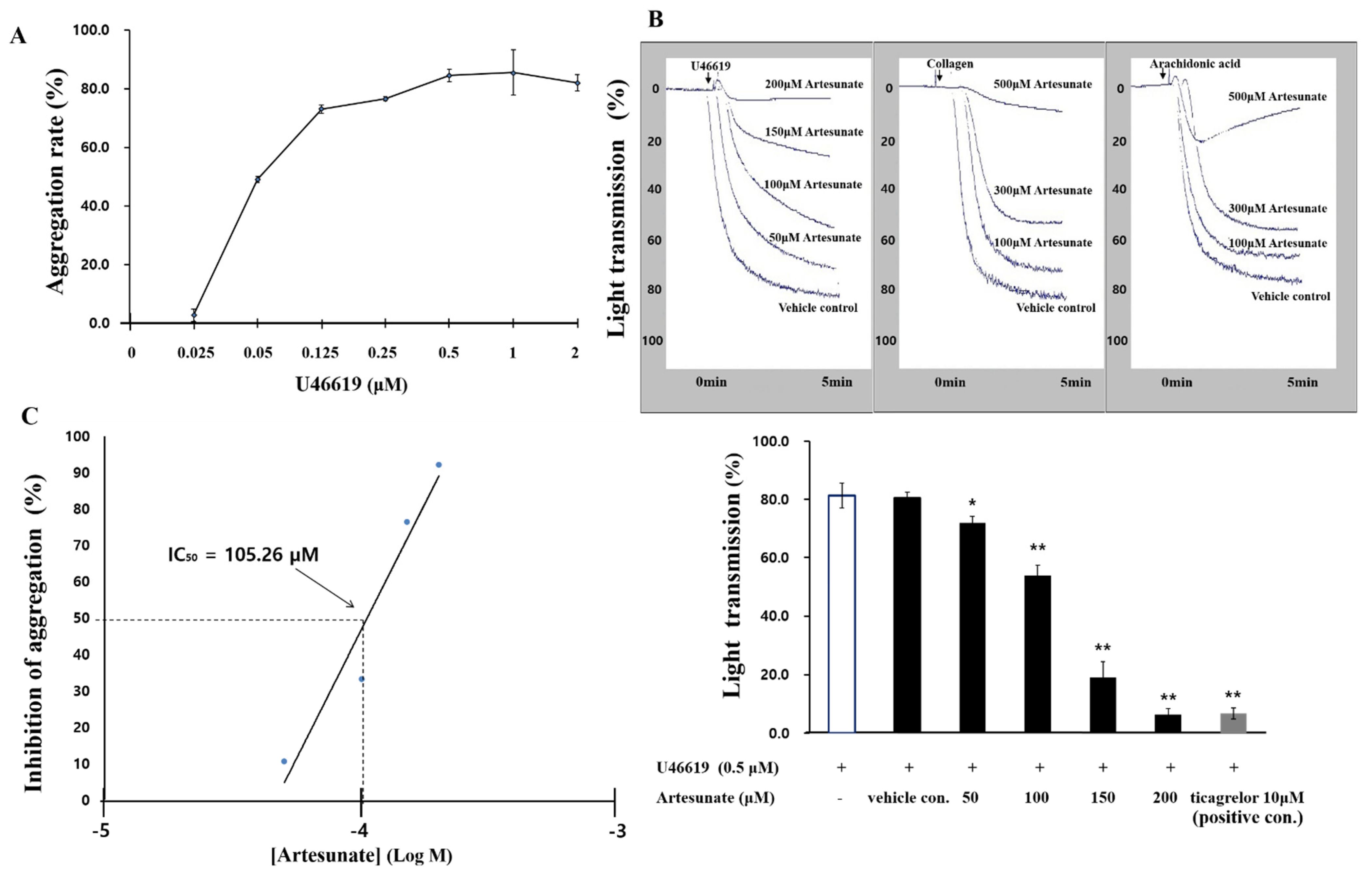
2.1. Effects of Artesunate on U46619-Induced Platelet Aggregation
2.2. Effects of Artesunate on the Cyclic Nucleotides Production
2.3. Effects of Artesunate on Intracellular Ca2+ Mobilization and IP3R Phosphorylation
2.4. Effects of Artesunate on VASP Phosphorylation and Fibrinogen Binding
2.5. Effects of Artesunate on TXA2 Production and Granule Secretion
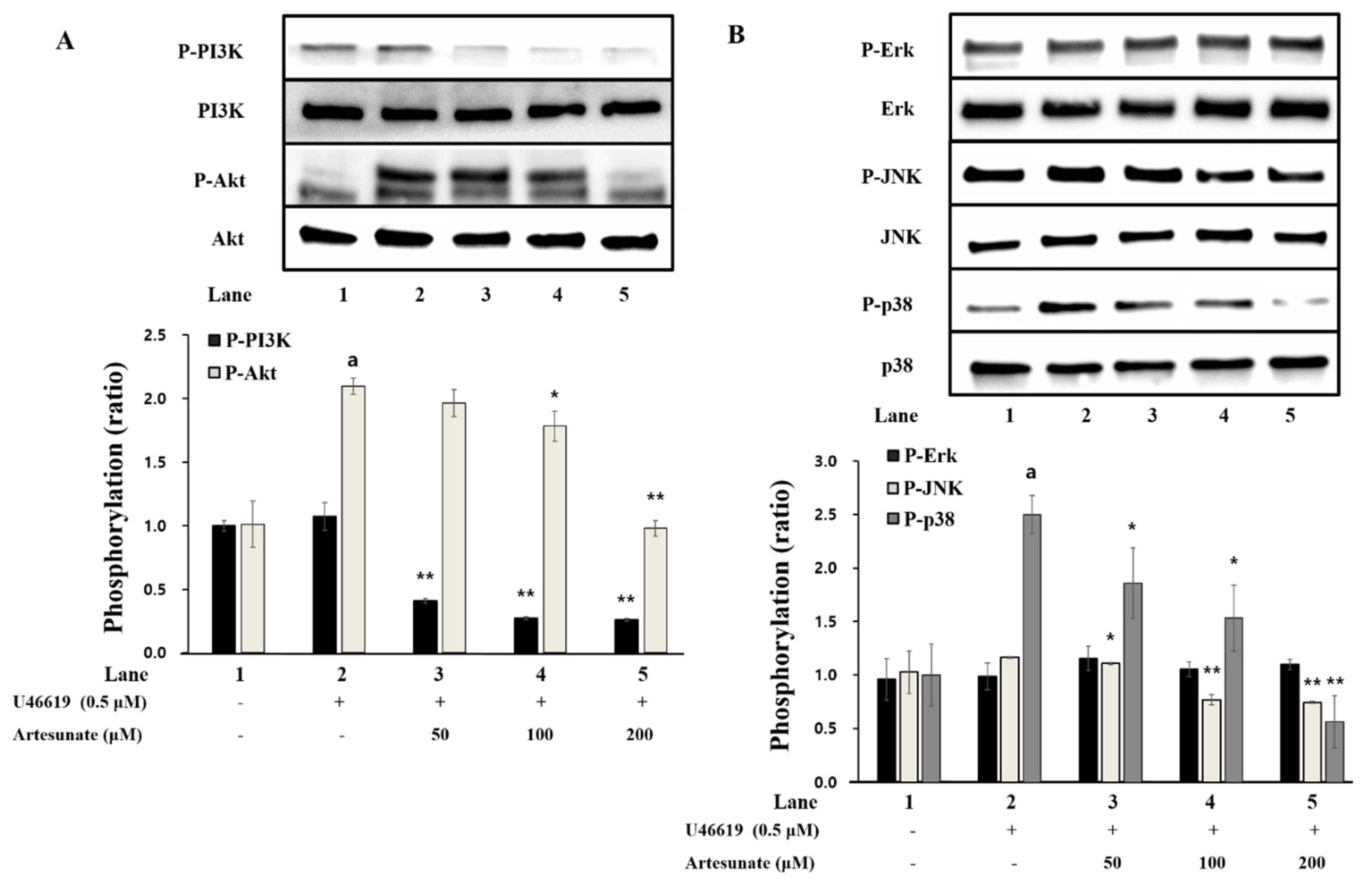
2.6. Effects of Artesunate on Phosphorylation of PI3K/Akt and MAPK
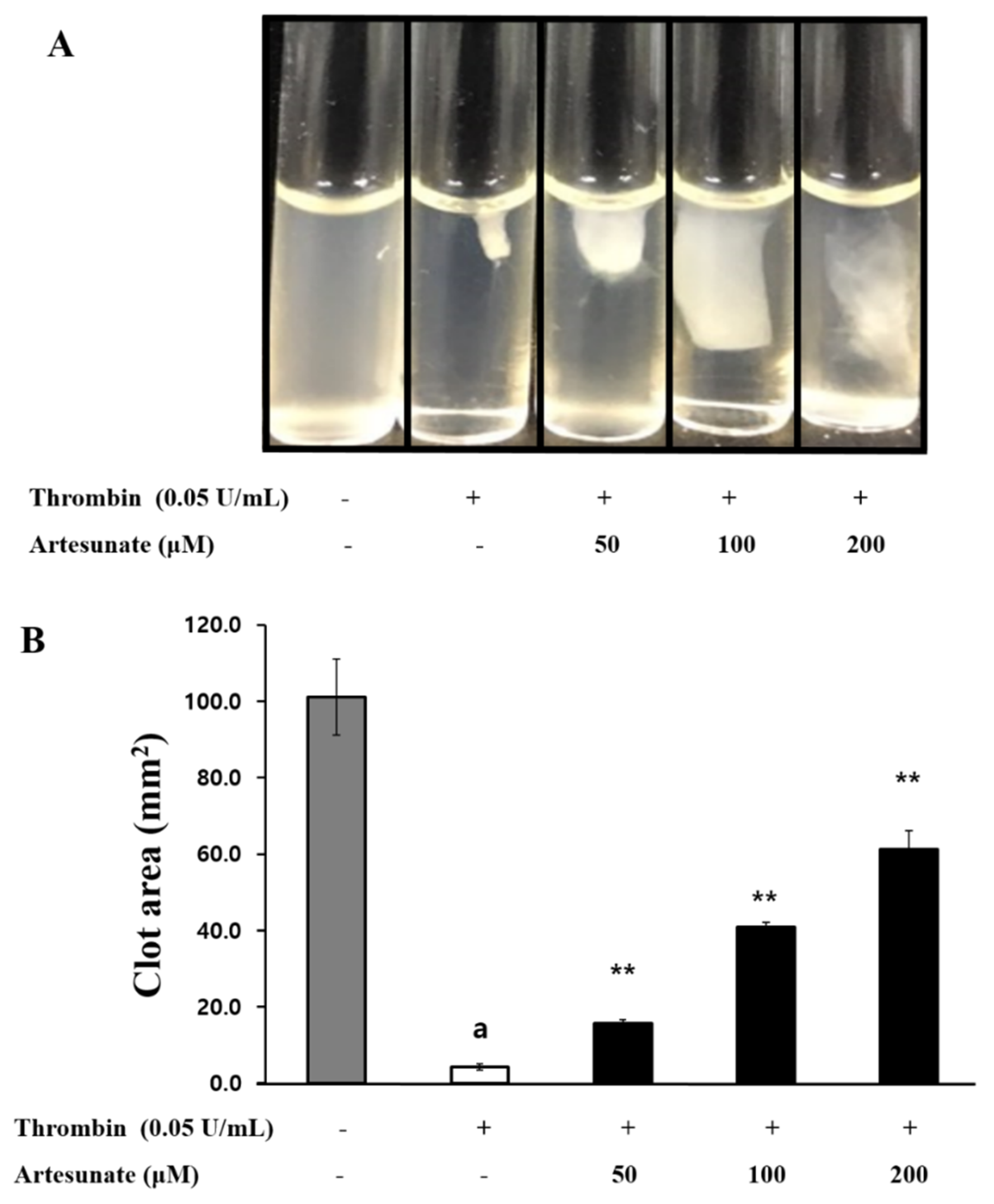
2.7. Effects of Artesunate on Platelet-Mediated Fibrin Clot Retraction
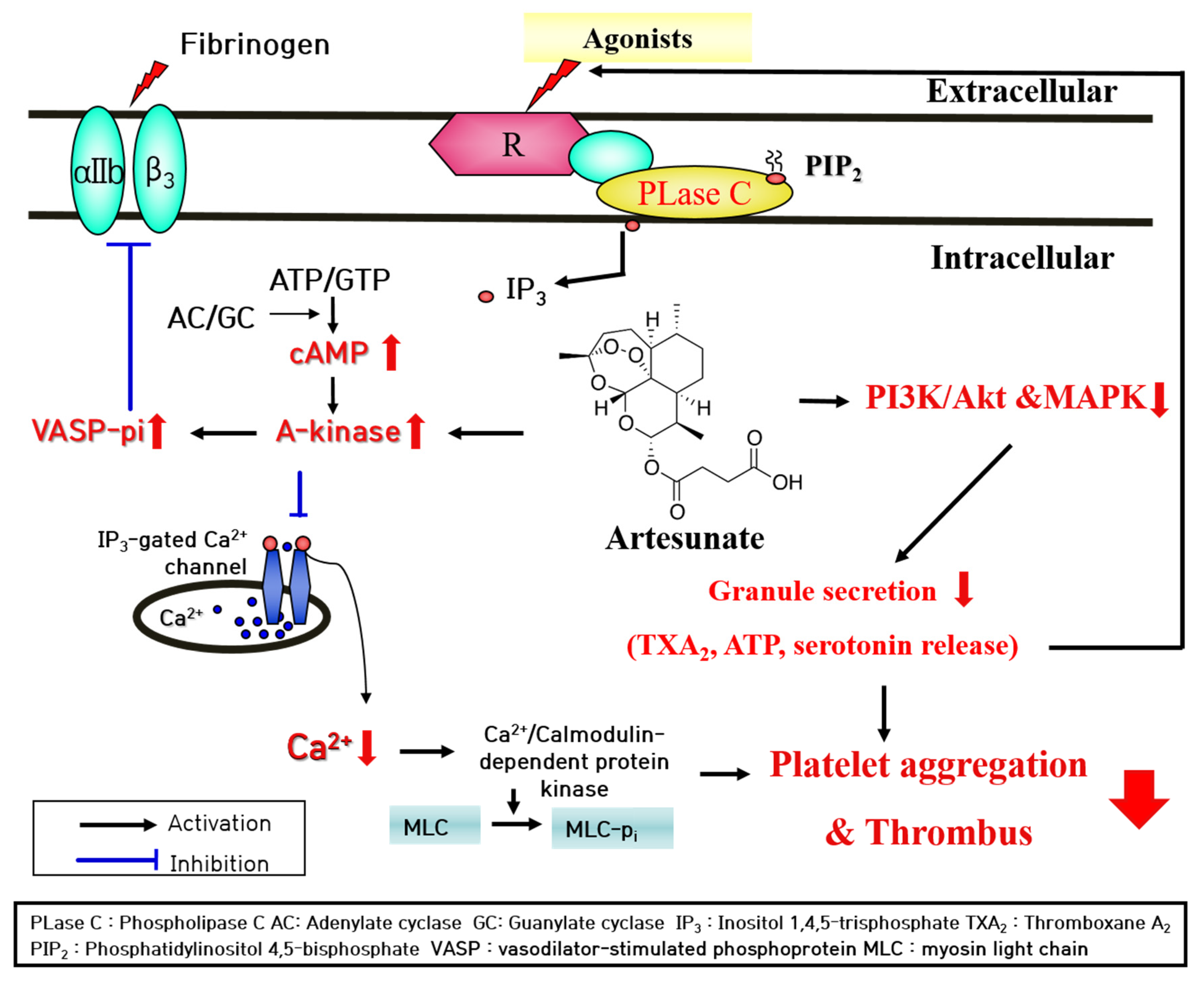
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Human Washed Platelets
4.3. Platelet Aggregation Measurement
4.4. Cytotoxicity Measurement
4.5. Cyclic Nucleotides (cAMP and cGMP) Production Measurement
4.6. Intracellular Ca2+ Mobilization Measurement
4.7. Fibrinogen Binding Measurement
4.8. TXB2 Production Measurement
4.9. ATP and Serotonin Release Measurement
4.10. Western Immunoblotting Measurement
4.11. Platelet-Mediated Fibrin Clot Retraction Measurement
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; de Ferranti, S.; Després, J.P.; Fullerton, H.J.; et al. Heart disease and stroke statistics-2016 update a report from the American Heart Association. Circulation 2016, 133, e38–e48. [Google Scholar] [PubMed]
- Andrews, R.K.; Berndt, M.C. Platelet physiology and thrombosis. Thromb Res. 2004, 114, 447–453. [Google Scholar] [CrossRef]
- Barrett, N.E.; Holbrook, L.; Jones, S.; Kaiser, W.J.; Moraes, L.A.; Rana, R.; Sage, T.; Stanley, R.G.; Tucker, K.L.; Wright, B.; et al. Future innovations in anti-platelet therapies. Br. J. Pharmacol. 2008, 154, 918–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badimon, L.; Vilahur, G.; Padro, T. Nutraceuticals and atherosclerosis: Human trials. Cardiovasc. Ther. 2010, 28, 202–215. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary prevention of cardiovascular disease with a Mediterranean diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irfan, M.; Kwon, T.H.; Yun, B.S.; Park, N.H.; Rhee, M.H. Eisenia bicyclis (brown alga) modulates platelet function and inhibits thrombus formation via impaired P2Y12 receptor signaling pathway. Phytomedicine 2018, 40, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Pandey, M.M.; Rawat, A. Traditional herbs: A remedy for cardiovascular disorders. Phytomedicine 2016, 23, 1082–1089. [Google Scholar] [CrossRef]
- Quinton, T.M.; Dean, W.L. Cyclic AMP-dependent phosphorylation of the inositol-1,4,5-trisphosphate receptor inhibits Ca2+ release from platelet membranes. Biochem. Biophys Res. Commun. 1992, 184, 893–899. [Google Scholar] [CrossRef]
- Menshikov, M.Y.U.; Ivanova, K.; Schaefer, M.; Drummer, C.; Gerzer, R. Influence of the cGMP analog 8-PCPT-cGMP on agonist-induced increases in cytosolic ionized Ca2+ and on aggregation of human platelets. Eur. J. Pharmacol. 1993, 245, 281–284. [Google Scholar] [CrossRef]
- Schwarz, U.R.; Walter, U.; Eigenthaler, M. Taming platelets with cyclic nucleotides. Biochem. Pharmacol. 2001, 62, 1153–1161. [Google Scholar] [CrossRef]
- Cavallini, L.; Coassin, M.; Borean, A.; Alexandre, A. Prostacyclin and sodium nitroprusside inhibit the activity of the platelet inositol 1,4,5-trisphosphate receptor and promote its phosphorylation. J. Biol. Chem. 1996, 271, 5545–5551. [Google Scholar] [CrossRef] [Green Version]
- Laurent, V.; Loisel, T.P.; Harbeck, B.; Wehman, A.; Gröbe, L.; Jockusch, B.M.; Wehland, J.; Gertler, F.B.; Carlier, M.F. Role of proteins of the Ena/VASP family in actin-based motility of Listeria monocytogenes. J. Cell Biol. 1999, 144, 1245–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudo, T.; Ito, H.; Kimura, Y. Phosphorylation of the vasodilator-stimulated phosphoprotein (VASP) by the anti-platelet drug, cilostazol, in platelets. Platelets 2003, 14, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Irfan, M.; Jeong, D.; Kwon, H.W.; Shin, J.H.; Park, S.J.; Kwak, D.; Kim, T.H.; Lee, D.H.; Park, H.J.; Rhee, M.H. Ginsenoside-Rp3 inhibits platelet activation and thrombus formation by regulating MAPK and cyclic nucleotide signaling. Vasc. Pharmacol. 2018, 109, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Adam, F.; Kauskot, A.; Rosa, J.P.; Bryckaert, M. Mitogen activated protein kinases in hemostasis and thrombosis. J. Thromb. Haemost. 2008, 6, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Bugaud, F.; Nadal-Wollbold, F.; Lévy-Toledano, S.; Rosa, J.P.; Bryckaert, M. Regulation of c-jun-NH2 terminal kinase and extracellular-signal regulated kinase in human platelets. Blood 1990, 94, 3800–3805. [Google Scholar] [CrossRef]
- Kramer, R.M.; Roberts, E.F.; Strifler, B.A.; Johnstone, E.M. Thrombin induces activation of p38 MAP kinase in human platelets. J. Biol. Chem. 1995, 270, 27395–27398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadal-Wollbold, F.; Pawlowski, M.; Lévy-Toledano, S.; Berrou, E.; Rosa, J.P.; Bryckaert, M. Platelet ERK2 activation by thrombin is dependent on calcium and conventional protein kinases C but not Raf-1 or B-Raf. FEBS Lett. 2002, 531, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Michelson, A.D. Antiplatelet therapies for the treatment of cardiovascular disease. Nat. Rev. Drug Discov. 2010, 9, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Flevaris, P.; Li, Z.; Zhang, G.; Zheng, Y.; Liu, J.; Du, X. Two distinct roles of mitogen-activated protein kinases in platelets and a novel Rac1-MAPK-dependent integrin outside-in retractile signaling pathway. Blood 2009, 113, 893–901. [Google Scholar] [CrossRef] [Green Version]
- Kramer, R.M.; Roberts, E.F.; Um, S.L.; Börsch-Haubold, A.G.; Watson, S.P.; Fisher, M.J.; Jakubowski, J.A. p38 mitogen activated protein kinase phosphorylates cytosolic phospholipase A2 (cPLA2) in thrombin-stimulated platelets. Evidence that proline-directed phosphorylation is not required for mobilization of arachidonic acid by cPLA2. J. Biol Chem. 1996, 271, 27723–27729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNicol, A.; Shibou, T.S. Translocation and phosphorylation of cytosolic phospholipase A2 in activated platelets. Thromb. Res. 1998, 92, 19–26. [Google Scholar] [CrossRef]
- Chuang, W.Y.; Kung, P.H.; Kuo, C.Y.; Wu, C.C. Sulforaphane prevents human platelet aggregation through inhibiting the phosphatidylinositol 3-kinase/Akt pathway. Thromb. Haemost. 2013, 109, 1120–1130. [Google Scholar] [CrossRef]
- Thanaketpaisarn, O.; Waiwut, P.; Sakurai, H.; Saiki, I. Artesunate enhances TRAIL-induced apoptosis in human cervical carcinoma cells through inhibition of the NF-κB and PI3K/Akt signaling pathways. Int. J. Oncol. 2011, 39, 279–285. [Google Scholar]
- Cao, T.H.; Jin, S.G.; Fei, D.S.; Kang, K.; Jiang, L.; Lian, Z.Y.; Pan, S.H.; Zhao, M.R.; Zhao, M.Y. Artesunate protects against sepsis-induced lung injury via heme oxygenase-1 modulation. Inflammation 2016, 39, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Ng, D.S.; Liao, W.; Tan, W.D.; Chan, T.K.; Loh, X.Y.; Wong, W.F. Anti-malarial drug artesunate protects against cigarette smoke-induced lung injury in mice. Phytomedicine 2014, 21, 1638–1644. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Wang, B.; Cui, N.; Zhang, Y. Artesunate suppresses oxidative and inflammatory processes by activating Nrf2 and ROS-dependent p38 MAPK and protects against cerebral ischemia-reperfusion injury. Mol. Med. Rep. 2018, 17, 6639–6646. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Luo, H.S.; Li, M.; Tan, S.Y. Artesunate inhibits the growth and induces apoptosis of human gastric cancer cells by downregulating COX-2. Onco Targets Ther. 2015, 8, 845–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.S.; Li, J.; Wang, Z.; Mi, C.; Ma, J.; Piao, L.X.; Xu, G.H.; Li, X.; Jin, X. Artemisinin inhibits inflammatory response via regulating NF-κB and MAPK signaling pathways. Immunopharmacol. Immunotoxicol. 2016, 21, 1–9. [Google Scholar] [CrossRef]
- Kamruzzaman, S.M.; Endale, M.; Oh, W.J.; Park, S.C.; Kim, T.H.; Lee, I.K.; Cho, J.Y.; Park, H.J.; Kim, S.K.; Yun, B.S.; et al. Antiplatelet activity of Phellinus baummii methanol extract is mediated by cyclic AMP elevation and inhibition of collagen-activated integrin-α(IIb) β3 and MAP kinase. Phytother. Res. 2011, 25, 1596–1603. [Google Scholar] [CrossRef]
- Berridge, M.J.; Irvine, R.F. Inositol phosphates and cell signalling. Nature 1989, 341, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Tanaka, T.; Hidaka, H. Ca2+-calmodulin-dependent phosphorylation and platelet secretion. Nature 1980, 287, 863–865. [Google Scholar] [CrossRef] [PubMed]
- Kuo, J.F.; Andersson, R.G.; Wise, B.C.; Mackerlova, L.; Salomonsson, I.; Brackett, N.L.; Katoh, N.; Shoji, M.; Wrenn, R.W. Calciumdependent protein kinase: Widespread occurrence in various tissues and phyla of the animal kingdom and comparison of effects of phospholipid, calmodulin, and trifluoperazine. Proc. Natl. Acad. Sci. USA 1980, 77, 7039–7043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wentworth, J.K.; Pula, G.; Poole, A.W. Vasodilator-stimulated phosphoprotein (VASP) is phosphorylated on Ser157 by protein kinase C-dependent and-independent mechanisms in thrombin-stimulated human platelets. Biochem. J. 2006, 393, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Napeñas, J.J.; Oost, F.C.; DeGroot, A.; Loven, B.; Hong, C.H.; Brennan, M.T.; Lockhart, P.B.; van Diermen, D.E. Review of postoperative bleeding risk in dental patients on antiplatelet therapy. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2013, 115, 491–499. [Google Scholar] [CrossRef]
- Gao, J.; Tao, J.; Liang, W.; Zhao, M.; Du, X.; Cui, S.; Duan, H.; Kan, B.; Su, X.; Jiang, Z. Identification and characterization of phosphodiesterases that specifically degrade 3′3′-cyclic GMP-AMP. Cell Res. 2015, 25, 539–550. [Google Scholar] [CrossRef]
- Haslam, R.J.; Dickinson, N.T.; Jang, E.K. Cyclic nucleotides and phos- phodiesterases in platelets. Thromb. Haemost. 1999, 82, 412–423. [Google Scholar]
- Chang, M.C.; Wang, T.M.; Yeung, S.Y.; Jeng, P.Y.; Liao, C.H.; Lin, T.Y.; Lin, C.C.; Lin, B.R.; Jeng, J.H. Antiplatelet effect by p-cresol, a uremic and environmental toxicant, is related to inhibition of reactive oxygen species, ERK/p38 signaling and thromboxane A2 production. Atherosclerosis 2011, 219, 559–565. [Google Scholar] [CrossRef]
- Marijon, A.; Bonnot, G.; Fourier, A.; Bringer, C.; Lavoignat, A.; Gagnieu, M.C.; Bienvenu, A.L.; Picot, S. Efficacy of intranasal administration of artesunate in experimental cerebral malaria. Malar. J. 2014, 13, 501. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, D.A. Integrin activation. J. Cell Sci. 2004, 117, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Jung-Hae Shin, J.H.; Kwon, H.W.; Irfan, M.; Rhee, M.H.; Lee, D.H. Ginsenoside Rk1 suppresses platelet mediated thrombus formation by downregulation of granule release and αIIbβ3 activation. J. Ginseng. Res. 2021, 45, 490–497. [Google Scholar]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, S.-S.; Kwon, H.-W.; Shin, J.-H.; Rhee, M.H.; Park, C.-E.; Lee, D.-H. Anti-Thrombotic Effects of Artesunate through Regulation of cAMP and PI3K/MAPK Pathway on Human Platelets. Int. J. Mol. Sci. 2022, 23, 1586. https://doi.org/10.3390/ijms23031586
Yoon S-S, Kwon H-W, Shin J-H, Rhee MH, Park C-E, Lee D-H. Anti-Thrombotic Effects of Artesunate through Regulation of cAMP and PI3K/MAPK Pathway on Human Platelets. International Journal of Molecular Sciences. 2022; 23(3):1586. https://doi.org/10.3390/ijms23031586
Chicago/Turabian StyleYoon, Shin-Sook, Hyuk-Woo Kwon, Jung-Hae Shin, Man Hee Rhee, Chang-Eun Park, and Dong-Ha Lee. 2022. "Anti-Thrombotic Effects of Artesunate through Regulation of cAMP and PI3K/MAPK Pathway on Human Platelets" International Journal of Molecular Sciences 23, no. 3: 1586. https://doi.org/10.3390/ijms23031586
APA StyleYoon, S.-S., Kwon, H.-W., Shin, J.-H., Rhee, M. H., Park, C.-E., & Lee, D.-H. (2022). Anti-Thrombotic Effects of Artesunate through Regulation of cAMP and PI3K/MAPK Pathway on Human Platelets. International Journal of Molecular Sciences, 23(3), 1586. https://doi.org/10.3390/ijms23031586