
An Efficient Marker Gene Excision Strategy Based on CRISPR/Cas9-Mediated Homology-Directed Repair in Rice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
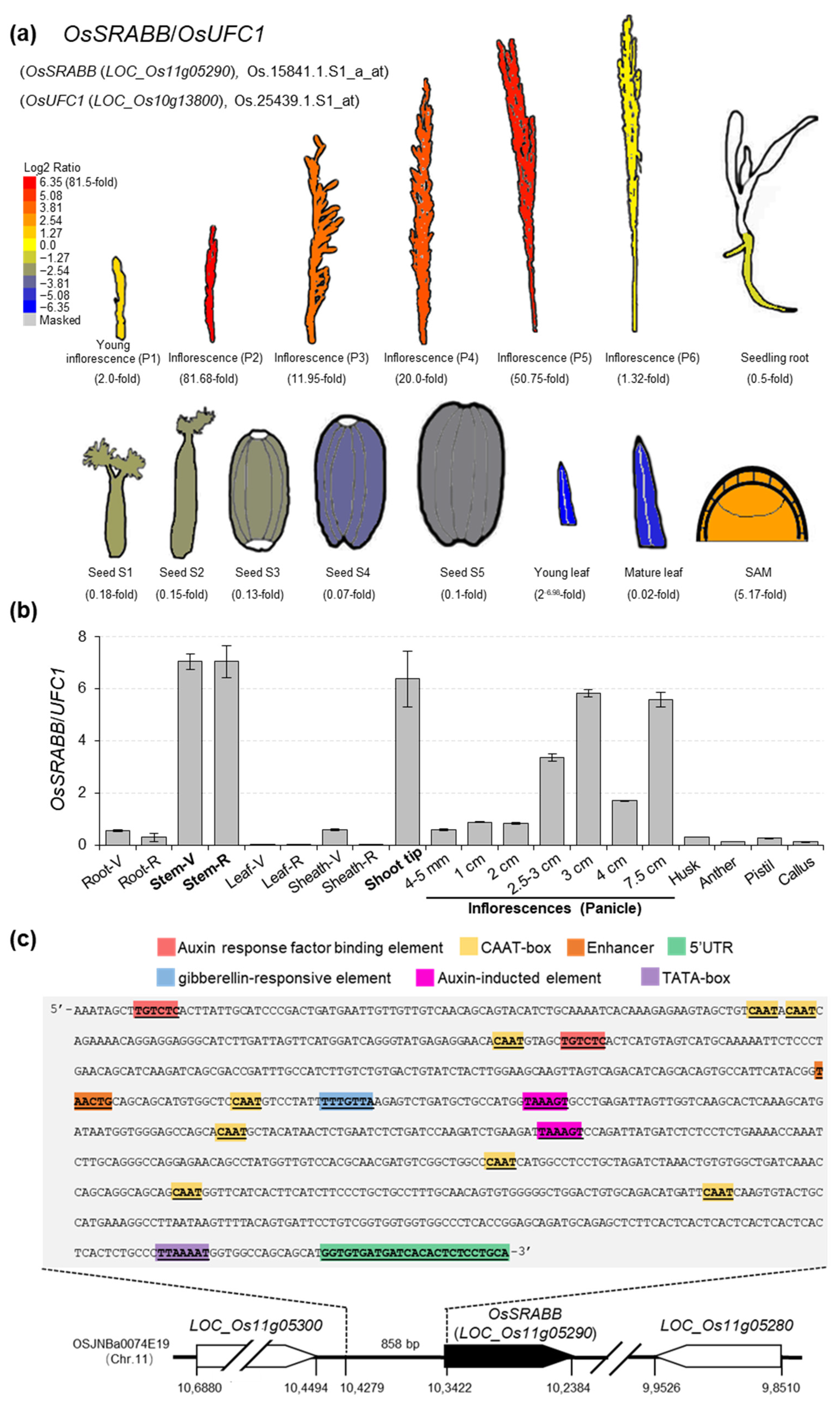
2.1. Identification of a Stem-, Shoot Tip- and Inflorescence-Strong Promoter
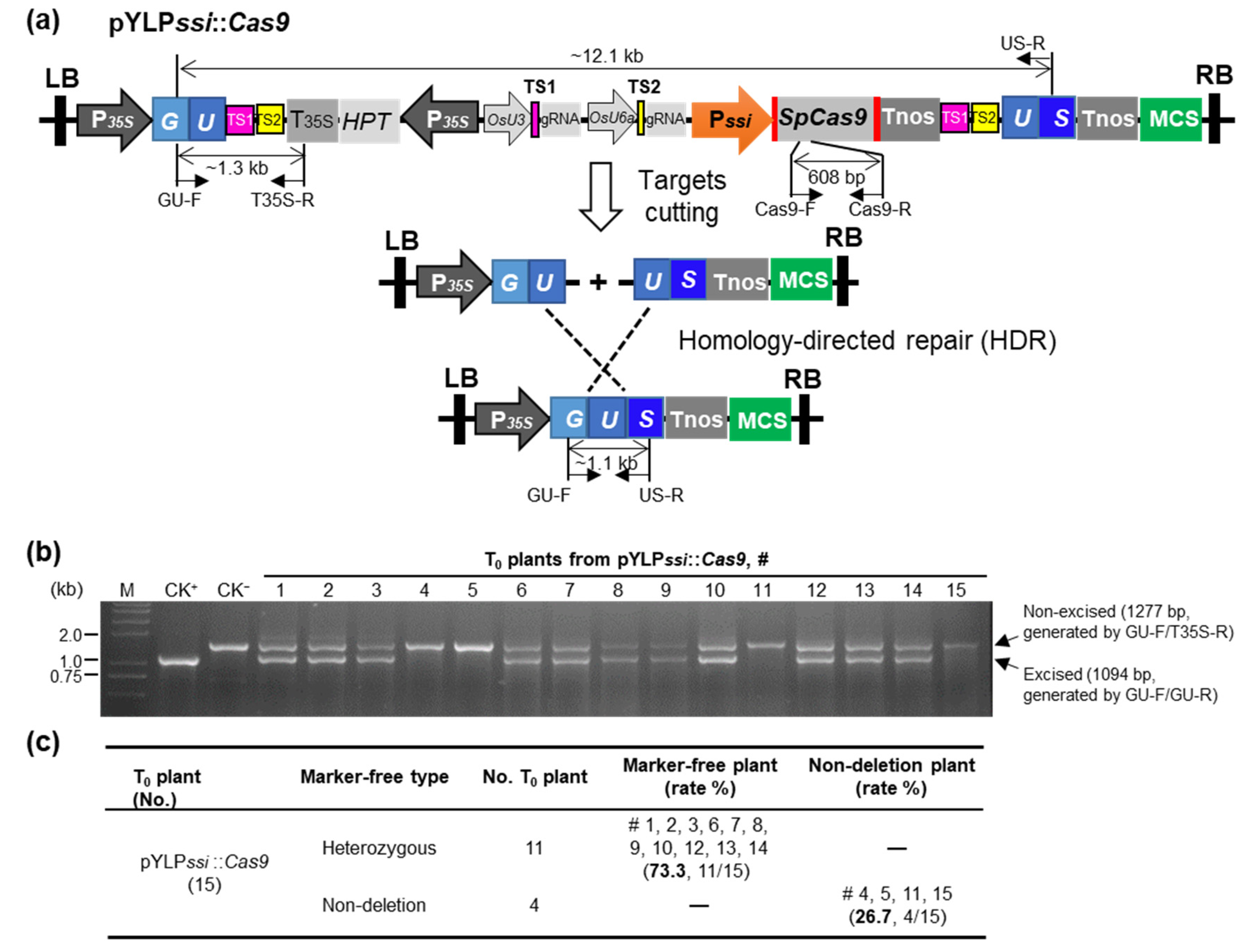
2.2. Development of a Marker-Free Transformation Tool Based on the Pssi Driving CRISPR/Cas9-Mediated HDR
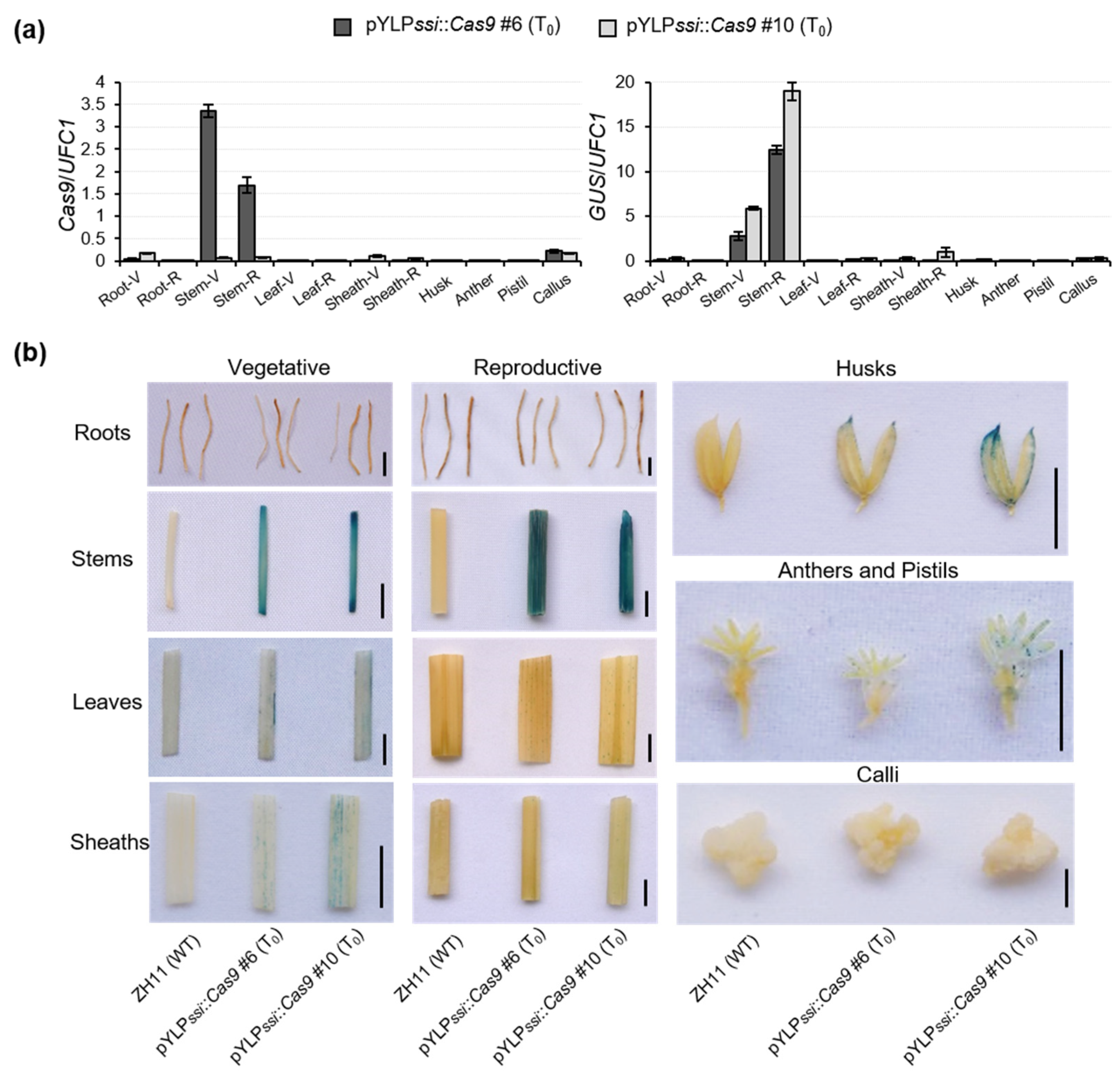
2.3. The PssiCHMF Tool Enables Efficient Excision of Marker Gene
2.4. PssiCHMF Is a Highly-Efficient and Time-Saving Marker Excision System
3. Discussion
4. Materials and Methods
4.1. Comprehensive Evaluation of Gene Expression, Promoter Activity, and Cis-Elements
4.2. Vector Construction
4.3. Plant Materials and Transformation
4.4. qRT-PCR Analysis
4.5. GUS Reporter-Aided Analysis
4.6. Molecular Characterization of Marker-Free Transgenic Rice
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chong-Pérez, B.; Angenon, G. Strategies for Generating Marker-Free Transgenic Plants. Genet. Eng. 2013, 10, 5772–55573. [Google Scholar]
- Bai, X.; Wang, Q.; Chu, C. Excision of a selective marker in transgenic rice using a novel Cre/loxP system controlled by a floral specific promoter. Transgenic Res. 2008, 17, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Darwish, N.A.; Khan, R.S.; Ntui, V.O.; Nakamura, I.; Mii, M. Generation of selectable marker-free transgenic eggplant resistant to Alternaria solani using the R/RS site-specific recombination system. Plant Cell Rep. 2014, 33, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pan, L.; Bi, D.; Tian, X.; Li, L.; Xu, Z.; Wang, L.; Zou, X.; Gao, X.; Yang, H.; et al. Generation of Marker-Free Transgenic Rice Resistant to Rice Blast Disease Using Ac/Ds Transposon-Mediated Transgene Reintegration System. Front. Plant Sci. 2021, 12, 644437. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.-Y.; Li, B.; Zhang, J.-R. Production of Marker-free Transgenic Tobacco Plants by FLP/frt Recombination System. Chin. J. Biotechnol. 2006, 22, 744–749. [Google Scholar] [CrossRef]
- Wang, K.; Liu, H.; Du, L.; Ye, X. Generation of marker-free transgenic hexaploid wheat via an Agrobacterium-mediated co-transformation strategy in commercial Chinese wheat varieties. Plant Biotechnol. J. 2017, 15, 614–623. [Google Scholar] [CrossRef]
- Zubko, E.; Scutt, C.; Meyer, P. Intrachromosomal recombination between attP regions as a tool to remove selectable marker genes from tobacco transgenes. Nat. Biotechnol. 2000, 18, 442–445. [Google Scholar] [CrossRef]
- Khattri, A.; Nandy, S.; Srivastava, V. Heat-inducible Cre-lox system for marker excision in transgenic rice. J. Biosci. 2011, 36, 37–42. [Google Scholar] [CrossRef]
- Sreekala, C.; Wu, L.; Gu, K.; Wang, D.; Tian, D.; Yin, Z. Excision of a selectable marker in transgenic rice (Oryza sativa L.) using a chemically regulated Cre/loxP system. Plant Cell Rep. 2005, 24, 86–94. [Google Scholar] [CrossRef]
- Zhu, Q.; Yu, S.; Zeng, D.; Liu, H.; Wang, H.; Yang, Z.; Xie, X.; Shen, R.; Tan, J.; Li, H.; et al. Development of “Purple Endosperm Rice” by Engineering Anthocyanin Biosynthesis in the Endosperm with a High-Efficiency Transgene Stacking System. Mol. Plant 2017, 10, 918–929. [Google Scholar] [CrossRef] [Green Version]
- Kurachi, M.; Ngiow, S.; Kurachi, J.; Chen, Z.; Wherry, E.J. Hidden Caveat of Inducible Cre Recombinase. Immunity 2019, 51, 591–592. [Google Scholar] [CrossRef] [PubMed]
- Thakore, P.I.; Gersbach, C.A. Chapter 3—Genome Engineering for Therapeutic Applications. In Translating Gene Therapy to the Clinic; Academic Press: Cambridge, MA, USA, 2015; pp. 27–43. [Google Scholar]
- Kaplan, A.R.; Glazer, P.M. Pharmacological methods to transcriptionally modulate double-strand break DNA repair. Int. Rev. Cell Mol. Biol. 2020, 354, 187–213. [Google Scholar] [PubMed]
- Davies, J.P.; Kumar, S.; Sastry-Dent, L. Chapter Three—Use of Zinc-Finger Nucleases for Crop Improvement. Prog. Mol. Biol. Transl. Sci. 2017, 149, 47–63. [Google Scholar] [PubMed]
- El Khoury, L.Y.; Campbell, J.M.; Clark, K.J. Chapter 17—The Transition of Zebrafish Functional Genetics from Random Mutagenesis to Targeted Integration. In Molecular-Genetic and Statistical Techniques for Behavioral and Neural Research; Elsevier: Amsterdam, The Netherlands, 2018; pp. 401–416. [Google Scholar]
- Boel, A.; De Saffel, H.; Steyaert, W.; Callewaert, B.; De Paepe, A.; Coucke, P.J.; Willaert, A. CRISPR/Cas9-mediated homology-directed repair by ssODNs in zebrafish induces complex mutational patterns resulting from genomic integration of repair-template fragments. Dis. Models Mech. 2018, 11, dmm035352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, V.T.; Weber, T.; Wefers, B.; Wurst, W.; Sander, S.; Rajewsky, K.; Kühn, R. Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells. Nat. Biotechnol. 2015, 33, 543–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Lu, Y.; Botella, J.R.; Mao, Y.; Hua, K.; Zhu, J.K. Gene Targeting by Homology-Directed Repair in Rice Using a Geminivirus-Based CRISPR/Cas9 System. Mol. Plant 2017, 10, 1007–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.-h.; Wang, B.; Li, N.; Tang, Y.; Shengbao, Y.; Yang, T.; Xu, J.; Guo, C.; Yan, P.; Wang, Q.; et al. CRISPR/Cas9-induced Targeted Mutagenesis and Gene Replacement to Generate Long-shelf Life Tomato Lines. Sci. Rep. 2017, 7, 11874. [Google Scholar] [CrossRef] [PubMed]
- Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef]
- Ali, S.; Khan, N.; Xie, L. Molecular and Hormonal Regulation of Leaf Morphogenesis in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 5132. [Google Scholar] [CrossRef]
- Reinhardt, D.; Mandel, T.; Kuhlemeier, C. Auxin regulates the initiation and radial position of plant lateral organs. Plant Cell 2000, 12, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Guan, C.; Jiao, Y. Interplay between the shoot apical meristem and lateral organs. aBIOTECH 2020, 1, 178–184. [Google Scholar] [CrossRef]
- Yao, W.; Li, G.; Yu, Y.; Ouyang, Y. funRiceGenes dataset for comprehensive understanding and application of rice functional genes. GigaScience 2018, 7, gix119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Tang, Y.; Ruan, N.; Dang, Z.; Huang, Y.; Miao, W.; Xu, Z.; Li, F. The Rice BZ1 Locus Is Required for Glycosylation of Arabinogalactan Proteins and Galactolipid and Plays a Role in both Mechanical Strength and Leaf Color. Rice 2020, 13, 41. [Google Scholar] [CrossRef]
- Moon, S.; Jung, K.H.; Lee, D.E.; Lee, D.Y.; Lee, J.; An, K.; Kang, H.G.; An, G. The rice FON1 gene controls vegetative and reproductive development by regulating shoot apical meristem size. Mol. Cells 2006, 21, 147–152. [Google Scholar] [PubMed]
- Wang, Y.; Wang, D.; Gan, T.; Liu, L.; Long, W.; Wang, Y.; Niu, M.; Li, X.; Zheng, M.; Jiang, L.; et al. CRL6, a member of the CHD protein family, is required for crown root development in rice. Plant Physiol. Biochem. 2016, 105, 185–194. [Google Scholar] [CrossRef]
- Li, S.; Xia, L. Precise gene replacement in plants through CRISPR/Cas genome editing technology: Current status and future perspectives. aBIOTECH 2020, 1, 58–73. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Rehman, S.; Tang, X.; Gu, K.; Fan, Q.; Chen, D.; Ma, W. Methodologies for Improving HDR Efficiency. Front. Genet. 2018, 9, 691. [Google Scholar] [CrossRef]
- Mlynárová, L.; Conner, A.; Nap, J.-P. Directed microspore-specific recombination of transgenic alleles to prevent pollen-mediated transmission of transgenes. Plant Biotechnol. J. 2006, 4, 445–452. [Google Scholar] [CrossRef]
- Tan, J.; Zhao, Y.; Wang, B.; Hao, Y.; Wang, Y.; Li, Y.; Luo, W.; Zong, W.; Li, G.; Chen, S.; et al. Efficient CRISPR/Cas9-based plant genomic fragment deletions by microhomology-mediated end joining. Plant Biotechnol. J. 2020, 18, 2161–2163. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Nijhawan, A.; Arora, R.; Agarwal, P.; Ray, S.; Sharma, P.; Kapoor, S.; Tyagi, A.K.; Khurana, J.P. F-box proteins in rice. Genome-wide analysis, classification, temporal and spatial gene expression during panicle and seed development, and regulation by light and abiotic stress. Plant Physiol. 2007, 143, 1467–1483. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Takehisa, H.; Kamatsuki, K.; Minami, H.; Namiki, N.; Ikawa, H.; Ohyanagi, H.; Sugimoto, K.; Antonio, B.; Nagamura, Y. RiceXPro Version 3.0: Expanding the informatics resource for rice transcriptome. Nucleic Acids Res. 2013, 41, 1206–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Xie, W.; Chen, Y.; Tang, W.; Yang, J.; Ye, R.; Liu, L.; Lin, Y.; Xu, C.; Xiao, J.; et al. A dynamic gene expression atlas covering the entire life cycle of rice. Plant J. 2010, 61, 752–766. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A Robust CRISPR/Cas9 System for Convenient, High-Efficiency Multiplex Genome Editing in Monocot and Dicot Plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zeng, D.; Shen, R.; Ma, X.; Zhang, Q.; Chen, L.; Liu, Y.-G.; Zhu, Q. Rapid in vitro splicing of coding sequences from genomic DNA by isothermal recombination reaction-based PCR. Biotechnol. Biotechnol. Equip. 2016, 30, 864–868. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Ma, X.; Zhu, Q.; Zeng, D.; Li, G.; Liu, Y.-G. CRISPR-GE: A Convenient Software Toolkit for CRISPR-Based Genome Editing. Mol. Plant 2017, 10, 1246–1249. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Yang, Z.; Zhang, Q.; Chen, L.; Liu, Y.-G. Robust multi-type plasmid modification based on isothermal in vitro recombination. Gene 2014, 548, 39–42. [Google Scholar] [CrossRef]
- Nishimura, A.; Aichi, I.; Matsuoka, M. A protocol for Agrobacterium-mediated transformation in rice. Nat. Protoc. 2006, 1, 2796–2802. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Z.; Ding, Z.; Meng, H.; Shen, R.; Tang, H.; Liu, Y.G.; Chen, L. Public-transcriptome-database-assisted selection and validation of reliable reference genes for qRT-PCR in rice. Sci. China Life Sci. 2020, 63, 92–101. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Doyle, J.J.T.; Doyle, J.L. Isolation of Plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, J.; Wang, Y.; Chen, S.; Lin, Z.; Zhao, Y.; Xue, Y.; Luo, Y.; Liu, Y.-G.; Zhu, Q. An Efficient Marker Gene Excision Strategy Based on CRISPR/Cas9-Mediated Homology-Directed Repair in Rice. Int. J. Mol. Sci. 2022, 23, 1588. https://doi.org/10.3390/ijms23031588
Tan J, Wang Y, Chen S, Lin Z, Zhao Y, Xue Y, Luo Y, Liu Y-G, Zhu Q. An Efficient Marker Gene Excision Strategy Based on CRISPR/Cas9-Mediated Homology-Directed Repair in Rice. International Journal of Molecular Sciences. 2022; 23(3):1588. https://doi.org/10.3390/ijms23031588
Chicago/Turabian StyleTan, Jiantao, Yaxi Wang, Shuifu Chen, Zhansheng Lin, Yanchang Zhao, Yang Xue, Yuyu Luo, Yao-Guang Liu, and Qinlong Zhu. 2022. "An Efficient Marker Gene Excision Strategy Based on CRISPR/Cas9-Mediated Homology-Directed Repair in Rice" International Journal of Molecular Sciences 23, no. 3: 1588. https://doi.org/10.3390/ijms23031588
APA StyleTan, J., Wang, Y., Chen, S., Lin, Z., Zhao, Y., Xue, Y., Luo, Y., Liu, Y.-G., & Zhu, Q. (2022). An Efficient Marker Gene Excision Strategy Based on CRISPR/Cas9-Mediated Homology-Directed Repair in Rice. International Journal of Molecular Sciences, 23(3), 1588. https://doi.org/10.3390/ijms23031588