
Nutraceutical and Dietary Strategies for Up-Regulating Macroautophagy

Catalytic Longevity Foundation, San Diego, CA 92109, USA
Int. J. Mol. Sci. 2022, 23(4), 2054; https://doi.org/10.3390/ijms23042054
Submission received: 29 December 2021
/
Revised: 7 February 2022
/
Accepted: 8 February 2022
/
Published: 12 February 2022
(This article belongs to the Special Issue Molecular and Biological Mechanisms of Longevity)
Abstract
:Macroautophagy is a “cell cleansing” process that rids cells of protein aggregates and damaged organelles that may contribute to disease pathogenesis and the dysfunctions associated with aging. Measures which boost longevity and health span in rodents typically up-regulate macroautophagy, and it has often been suggested that safe strategies which can promote this process in humans may contribute to healthful aging. The kinase ULK1 serves as a trigger for autophagy initiation, and the transcription factors TFEB, FOXO1, ATF4 and CHOP promote expression of a number of proteins which mediate macroautophagy. Nutraceutical or dietary measures which stimulate AMPK, SIRT1, eIF5A, and that diminish the activities of AKT and mTORC1, can be expected to boost the activities of these pro-autophagic factors. The activity of AMPK can be stimulated with the phytochemical berberine. SIRT1 activation may be achieved with a range of agents, including ferulic acid, melatonin, urolithin A, N1-methylnicotinamide, nicotinamide riboside, and glucosamine; correction of ubiquinone deficiency may also be useful in this regard, as may dietary strategies such as time-restricted feeding or intermittent fasting. In the context of an age-related decrease in cellular polyamine levels, provision of exogenous spermidine can boost the hypusination reaction required for the appropriate post-translational modification of eIF5A. Low-protein plant-based diets could be expected to increase ATF4 and CHOP expression, while diminishing IGF-I-mediated activation of AKT and mTORC1. Hence, practical strategies for protecting health by up-regulating macroautophagy may be feasible.
1. Boosting Autophagy for Health Protection
Macroautophagy is the process whereby the contents of a cell, including whole organelles, are gradually taken up into forming autophagosomes for degradation in lysosomes, with preference given to the disposal of proteins and organelles that are damaged or marked via ubiquitination. Effective macroautophagy is often referred to as “cell cleansing”, and, when of moderate intensity, and appropriately balanced by synthesis of new proteins and organelles—particularly mitochondria, produced in the complex process of “mitochondrial biogenesis”—it is generally thought to enhance health span [1,2,3,4]. (Henceforth, macroautophagy will be referred to simply as “autophagy”—albeit chaperone-mediated autophagy and microautophagy are distinct processes that also assist in protein disposal within autophagosomes [5,6].) Measures which are known to increase median and maximal lifespan in rodents typically are associated with increased autophagy. Autophagy may be particularly protective with respect to neurodegenerative disorders in which intracellular aggregates of damaged proteins can lead to neuron dysfunction and death [7,8,9]. An age-related decrease in autophagy in lymphocytes appears to be a mediator of the immune senescence that puts the elderly at greater risk for infections and cancer [10]. For these reasons, it is rational to seek safe and practical nutraceutical and dietary measures which can induce a moderate up-regulation of autophagy. An understanding of the molecular biology regulating autophagy is a prerequisite to this search.
2. Key Drivers of Autophagy
Activation of the serine/threonine kinase ULK1 is a trigger for initiating the formation of phagophores, which are then elongated into the autophagosomes that engulf cell contents during their formation [11,12]. AMP-activated kinases (AMPK) and mammalian target of rapamycin complex 1 (mTORC1) act as key functional antagonists modulating ULK1 activity [13]. AMPK, which is activated by elevation of AMP+ADP, and hence serves as a monitor for the availability of the biochemically useful free energy provided by ATP, confers an activating phosphorylation on ULK1 [14,15,16,17]. mTORC1, whose activity is enhanced by growth factor activity via AKT and decreased by a cellular deficit of certain amino acids, phosphorylates ULK1 in a way that inhibits its activity [13,18]. The ability of AMPK to suppress mTORC1 activity by complementary phosphorylations of Raptor and TSC2 hence amplifies its positive impact on ULK1 activation [19,20,21].
Like AMPK, the deacetylase SIRT1 functions as a detector of cellular energy deficit; its obligate substrate is NAD+, and a relative paucity of oxidizable substrate enhances its activity by boosting the cytosolic NAD+/NADH ratio. SIRT1 activity can stimulate ULK1 indirectly, by increasing AMPK activation via LKB1. The serine/threonine kinase LKB1 activates AMPK via phosphorylation of Thr472 [22]. (Binding of AMP to AMPK aids this phosphorylation, and binding of AMP or ADP impedes its dephosphorylation [14]. Deacetylation of LKB1 by SIRT1 enhances its cytoplasmic localization and its interaction with its activator STRAD1, thereby enhancing its effective activity [23]. Hence, SIRT1 promotes LKB1-mediated activation of AMPK, leading to increased ULK1 activity.
The biosynthesis of autophagosomes and the lysosomes with which they ultimately fuse requires the synthesis of a vast array of proteins. However, a relatively small number of transcription factors can drive the synthesis of most or all of these proteins. In particular, transcription factors EB (TFEB), forkhead box O1 (FOXO1), activating transcription factor 4 (ATF4), and C/EBP homologous protein (CHOP) play a prominent role in this regard, and measures which boost their synthesis and/or activity can thereby enhance autophagy.
3. Modulation of TFEB Activity
TFEB is considered the master transcriptional regulator of the synthesis of a great number of proteins required for both lysosome and autophagosome formation [24]. It promotes the transcription of genes carrying a CLEAR regulatory element (coordinated lysosomal expression and regulation) in their promoters, to which TFEB binds; at least 471 such genes have been identified [25]. mTORC1 and ERK2 can phosphorylate TFEB on Ser142 and Ser211; by enabling binding of TFEB to the chaperone 14-3-3, these phosphorylations tend to promote the nuclear export of TFEB, such that it cannot drive transcription [26,27]. Within the cytoplasm, TFEB tends to associate with mTORC1 on the membrane of lysosomes; hence, mTORC1-mediated phosphorylation appears to be the chief mechanism keeping TFEB out of the nucleus [24].
In contrast, AMPK boosts TFEB’s transcriptional activity through several complementary mechanisms. As noted, AMPK functions to decrease mTORC1 activity. Furthermore, AMPK activates the phosphatase calcineurin in the microenvironment of the lysosomal surface, which can reverse the phosphorylations conferred on TFEB by mTORC1 [28,29]. AMPK achieves this indirectly by phosphorylating and activating the kinase PIKFYVE in the lysosomal membrane; this converts phosphatidylinositol-3-phosphate to phosphatidylinositol-3,5-diphosphate [30,31]. The latter can then interact with TRPML1 (also known as MCOLN1) calcium channels to increase their permeability to calcium [32,33]. The resulting increase in free calcium in the lysosomal membrane microenvironment activates calcineurin, enabling it to remove the Ser142 and Ser21 phosphorylations of TFEB [28,29]. Finally, AMPK can itself phosphorylate TFEB; whereas this does not influence TFEB’s subcellular location, it amplifies its transactivational activity [34].
Modulation of TFEB activity by acetylation appears to be complex. On the one hand, up-regulation of this acetylation by treatment with SAHA, an inhibitor of type I, II, and IV histone deacetylases, has been found to enhance TFEB-mediated transcription [35]. On the other hand, SIRT1-mediated deacetylation of K116 in TFEB has been reported to enhance its transcriptional activity by promoting its nuclear import and enhancing its binding to CLEAR elements [36,37]. Moreover, SIRT1 can boost TFEB activity less directly by enhancing AMPK activation. Hence, SIRT1 and AMPK collaborate to induce the nuclear import of TFEB while boosting its transactivational activity.
The synthesis of TFEB is modulated at the translational level by a rather remarkable mechanism. This protein contains two triproline motifs. The efficient translation of such proteins requires optimal activity of eukaryotic initiation factor 5A (eIF5A). This activity, in turn, requires enzymatic activity that converts a specific lysine in eIF5A to the unusual amino acid hypusine; eIF5A is the only mammalian protein known to contain a hypusine residue [38]. This conversion of lysine to hypusine requires the polyamine spermidine as a substrate [39]. Tissue levels of spermidine tend to decline with age, and recent studies show that these levels are sufficiently low in lymphocytes from elderly donors that provision of exogenous spermidine amplifies their hypusinated eIF5A level and thereby boosts the subnormally active autophagy in these lymphocytes [39,40,41].
4. Roles of ATF4, CHOP, and FOXO1 in Autophagy
Another transcription factor which promotes synthesis of a number of proteins required for autophagy and lysosomal activity is ATF4. The synthesis of this is enhanced in certain stress situations (the integrated stress response) promoting phosphorylation of eIF2α—a modification which selectively enhances translation of ATF4 mRNA [42]. ATF4, in turn, promotes transcription of the gene encoding the transcription factor CHOP [43]. ATF4 and CHOP collaborate in boosting the transcription of a number of genes coding for autophagosome/lysosomal proteins, whereas the transcription of other sets of genes important for autophagy can be driven by either ATF4 or CHOP acting alone [44]. Genes whose transcription ATF4 and/or CHOP promote include many of those involved in phagophore formation, such as ATGs 3, 5, 7, 10, 12 and 16, as well as Beclin1 [44].
The activity of the transcription factor FOXO1 is modulated by growth factor activity and by SIRT1-reversible acetylation. Growth factor activity, via AKT activation, phosphorylates FOXO1 in such a way that 14-3-3 promotes its nuclear export, abolishing its transcriptional activity [45]. Conversely, a deacetylation of FOXO1 promotes its nuclear import, and also enables it to drive the transcription of the gene coding for the G protein Rab7 [46,47]. The latter plays an essential role in enabling the fusion of late autophagosomes with lysosomes, a crucial step in autosomal flux [48,49].
5. Additional Impacts of AMPK and SIRT1 on Autophagy
In addition to up-regulation of ULK1 and TFEB activities, AMPK works in several other ways to promote autophagy [50,51,52]. It confers an activating phosphorylation on the class III phosphatidylinositol-3-kinase PIKC3/VPS34, which plays a crucial role in autophagosome formation [53,54]. Via phosphorylation of FOXO3a on Ser413, it increase the expression and activates the transcriptional activity of this factor, which promotes increased expression of autophagy mediators including LC3B-II, Gabarapl1, and Beclin1 [55,56]. AMPK activity is also required for SIRT1-mediated deacetylation and inactivation of BRD4, a repressor of the transcription of autophagic and lysosomal genes [57].
With respect to SIRT1, its activity also supports autophagy by deacetylating a number of different proteins mediating autophagy, including LC3, ATG5, AT7 and Beclin1 [58,59]. Protein acetylation might be construed as a signal of energy abundance, as the acetyl-CoA employed for protein acetylation derives from oxidation of substrate.
This overview predicts that measures which activate AMPK, SIRT1, and eIF5A, and promote phosphorylation of eIF2α, while down-regulating growth factor activities boosting AKT and mTORC1 activities, could be expected to up-regulate autophagy.
6. Nutraceutical Activation of AMPK with Berberine
With respect to AMPK, the nutraceutical berberine, a compound found in a range of herbs used in traditional Chinese medicine, has shown clinically useful activity for control of type 2 diabetes and hyperlipidemia, usually at intakes of 1–2 g daily in divided doses, and these benefits are at least partially attributable to activation of AMPK [60,61,62,63,64]. Indeed, berberine appears to activate AMPK in a manner analogous to the drug metformin, producing a partial inhibition of site I of the mitochondrial electron transport chain; the resulting rise in AMP and ADP enables AMPK to receive an activating phosphorylation from LKB1 [65]. Berberine’s ability to promote autophagy has been demonstrated in cell culture and rodent models [66,67,68].
It should be noted that a number of additional phytochemicals found in medicinal herbs have the potential to activate AMPK. These include compounds found in Chinese medicinal herbs such as mangiferin, astragaloside IV, and triterpenic acids of Cyclocarya paliurus [69]. The traditional anti-diabetic agent bitter melon (Momordica charantia) contains triterpenoids which can activate AMKP via activation of its upstream kinase Ca+2/calmodulin-dependent kinase kinase-β (CaMKKβ). Nor should it be forgotten that the prototypical pharmaceutical activator of AMPK, metformin, was developed by chemical modification of guanidine found in Galega officinalis (goat’s rue) [70]. The advantage of berberine is that it is a well-defined chemical compound available as an inexpensive nutraceutical that has an extensive track record of safe and effective clinical use.
7. Manifold Nutraceutical and Dietary Options for Activation of SIRT1
The natural lignin resveratrol has been shown to boost Sirt1 activity in rodent and cell culture studies; it is reported to increase Sirt1′s affinity for both NAD+ and acetylated substrate [71]. The basis of its effect in this regard is still unclear; some evidence suggests that it may directly activate Sirt1 via allosteric interaction, whereas other data suggest that its effect is indirect [72,73,74]. In any case, owing to inefficient oral absorption and rapid conjugation (sulfation and glucuronidation) in intestinal mucosa and liver, most efforts to demonstrate that oral resveratrol can activate Sirt1 in human tissues have failed to observe an effect in this regard [75,76]. Efforts to develop resveratrol preparations with higher absorption, and the possibility that resveratrol might exert some Sirt1-independent health benefits, cannot be ruled out [75].
Nonetheless, a number of other nutraceuticals have emerged as potentially useful for SIRT1 activation in recent studies. Berberine can be expected to boost SIRT1 activity, as AMPK, via induction of nicotinamide phosphoribosyltransferase (NAMPT), can increase the availability of SIRT1’s obligate substrate NAD+; NAMPT catalyzes the initial and rate-limiting step in the conversion of free nicotinamide (a product of SIRT1 activity which causes feedback inhibition) to NAD+ [77,78,79]. An alternative way to increase cellular levels of NAD+ is to administer the natural metabolite nutraceutical nicotinamide riboside (NR), which after phosphorylation enters the pathway of NAD+ synthesis. NR has been shown to enhance SIRT1 in a number of cellular and pre-clinical studies, although the dose schedule that might achieve this effect clinically is still being assessed [80,81,82]. When mitochondria are functionally deficient in their obligate cofactor ubiquinone (coenzyme Q10 in humans), the backup of electrons in the electron transport chain which this induces could be expected to decrease the NAD+/NADH ratio, compromising SIRT1 activity; hence, correction of suboptimal ubiquinone status could be expected to aid effective SIRT1 activity [83]. For optimized absorption, ubiquinone is most effectively administered in its reduced form ubiquinol, in nanoliposomes; doses of 200 mg daily have shown clinical efficacy [84,85,86,87,88,89].
Ferulic acid has shown remarkable anti-inflammatory and antioxidant activity in pre-clinical studies and, as sodium ferulate, is widely used in cardiovascular medicine in China [90,91]. Recent studies have found that this may reflect, in large part, the ability of ferulic acid to up-regulate SIRT1 expression at the mRNA and protein level [92,93,94,95,96,97]. This effect has been demonstrated in a number of cell types, and hence appears to be a general effect; its mechanistic basis remains unclear. Dietary anthocyanins, while very poorly absorbed intact, are converted to ferulic acid by gut bacteria, which is then absorbed [98]. It has been postulated that the health benefits attributed to anthocyanin-rich diets may be traceable to ferulic acid [90]. This compound also occurs in conjugated form in a range of whole grains, fruits, vegetables, and legumes; it is partially available from these sources owing to gut esterase activities. In whole grains, ferulic acid serves as a cross-linking agent for arabinoxylan insoluble fiber found in bran; it has been suggested that, in conjunction with phytate, magnesium, and zinc, ferulic acid may be largely responsible for the health benefits associated with regular consumption of whole grains [99]. In a recent controlled clinical study, 1 g ferulic acid daily (500 mg twice daily) was found to decrease C-reactive protein by about one-third in hyperlipidemic patients [100].
Another phytochemical derived from gut bacterial activity is urolithin A, an absorbable metabolite produced from the ellagitannins found in pomegranate and a range of other plant foods; it and its fellow metabolite urolithin B seem likely to mediate the health benefits linked to dietary consumption of pomegranates and pomegranate juice [101,102,103,104]. Several recent rodent and cell culture studies report that urolithin A can modestly increase both the protein expression of SIRT1 and the NAD+/NADH ratio; the basis of these effects remains obscure [105,106]. Urolithin A has only recently become available as a nutraceutical, and further clinical study is needed to establish worthwhile dose levels. Administration of 500–1000 mg daily to elderly humans was found to be safe and associated with increased skeletal muscle markers of mitochondrial biogenesis [107].
The anti-inflammatory and antioxidant effects of melatonin supplementation may be largely attributable to its ability to boost SIRT1 expression at the mRNA level, possibly through activation of the Bmal1 transcription factor, which drives the transcription of genes coding for both SIRT1 and Nrf2 [108,109,110,111,112,113]. Melatonin, usually in doses of 3 mg or more, is commonly administered prior to bedtime, so as not to impede the natural diurnal rhythm induced by this neurohormone.
Whereas nicotinamide can be employed for re-synthesis of NAD+, its catabolism begins with a methylation of its ring nitrogen, yielding the compound N1-methylnicotinamide (MNA) [114]. While initially assumed to be an inactive catabolite, studies nearly two decades ago found that (MNA) has anti-inflammatory activity [115]. Subsequent research has shown that MNA can be protective in rodent models of atherosclerosis, thrombosis, diabetes, metabolic syndrome, polycystic ovary syndrome, Alzheimer’s disease, depression, and age-related hearing loss [116,117,118,119,120,121,122,123,124]. These versatile benefits may be attributable, at least in part, to the ability of MNA to slow proteasomal catabolism of SIRT1, increasing its protein half-life [125]. This effect is associated with decreased phosphorylation of Ser47; phosphorylation at this site by JNK1 prepares SIRT1 for ubiquitination and subsequent proteasomal degradation [126]. How MNA achieves this effect is not yet clear. Evidently, this effect of NMA should be complementary to that of nutraceuticals that boost SIRT1 synthesis and/or enhance the NAD+/NADH ratio. Although MNA has achieved EU approval for use as a nutraceutical, this compound is not yet commercially available as a nutraceutical.
O-GlcNAcylation of SIRT1 has been reported to enhance its deacetylase activity; hence, elevating cellular pools of UDP-N-acetylglucosamine via high-dose glucosamine supplementation may have potential for boosting SIRT1 activity [127,128]. Arguably, this might be pertinent to the modest but significant impact of supplemental glucosamine on longevity in mice, and on the observation that prolonged glucosamine supplementation is associated with decreased global mortality in humans [129,130,131]. The usual dose of glucosamine in management of osteoarthritis is 1.5 g daily, but it has been argued that somewhat higher doses might be required for optimal clinical efficacy; 3 g daily is reported to amplify flow-mediated vasodilation in healthy volunteers—consistent with a role for Sirt1 in boosting endothelial nitric oxide synthase activity [128,132,133].
Calorie restriction and episodic fasting have the potential to increase SIRT1 activity by reducing the availability of oxidizable substrate to cells, and thereby lowering the NAD+/NADH ratio [134,135,136]. These strategies require dedication that may make them impractical for most people, but time-restricted feeding—in which calorie intake is restricted to only one or two meals daily—may be more feasible for many. Eating only in the morning has been shown to boost autophagy prior to resumption of food in the hours prior to food consumption [137]. The senior author has long practiced an alternative strategy wherein aerobic exercise is performed in morning fasting metabolism, and food consumption is postponed to the evening hours; carnitine supplementation might aid this protocol by optimizing the liver’s ability to oxidize fat as a satiety signal [138].
The possibility that ketogenic diets—high-fat diets low in both carbohydrate and protein, such that hepatic ketogenesis is maximized—might influence autophagy and/or Sirt1 activity merits more study. Whereas glucose metabolism via glycolysis reduces cytosolic NAD+ to NADH—an effect expected to diminish Sirt1 activity—ketones are metabolized entirely within mitochondria; moreover, the ATP that this metabolism generates could be expected to suppress glycolysis [139]. Increased markers of autophagy have been observed in the brains of type 1 diabetics who have died of ketoacidotic coma, and increased autophagy has also been observed in rodents fed ketogenic diets or infused with ketones [140,141,142].
Aerobic exercise can enhance AMPK and SIRT1 activity in cardiac muscle and the exercised skeletal muscle by temporarily boosting AMP levels and the cytosolic NAD+/NADH ratio [143,144,145]. Although this activation is transitory, it can exert a longer-term effect owing to activation of transcription factors driving the synthesis of long-lived proteins; hence, both autophagy and mitochondrial biogenesis are promoted.
8. Plant-Based Diets May Support Autophagy
A practical way to achieve a modest enhancement in ATF4 production (and hence CHOP as well), while also down-regulating the systemic growth factor activity of IGF-I, may be consumption of a plant-based diet of modest protein content [146]. The enzyme GCN2 is a kinase for eIF2α that is activated by a relative paucity of one or more essential amino acids [147,148]. The ability of GCN2 to activate autophagy hence makes homeostatic sense, as autophagy can increase the availability of essential amino acids for synthesis of essential proteins by degrading existing proteins. Activation of GCN2 in the liver by protein restriction increases hepatic synthesis of fibroblast growth factor-21 (FGF21); ATF4 drives transcription of the gene coding for this hormone [149]. This FGF21 can then act in an autocrine manner on hepatocytes to decrease their responsiveness to growth hormone signaling—an effect which suppresses hepatic secretion of IGF-I [150]. Consumption of vegan or low-protein diets has been associated with decreased plasma IGF-1 and increased plasma FGF21 in epidemiological and clinical studies [146,151,152]. It is notable that genetic overexpression of FGF21 has been found to markedly enhance median and maximal lifespan in mice, and mice with lower IGF-I levels typically experience greater longevity [153,154]. This effect might also help to explain why smaller dog breeds have notably longer lifespans than larger breeds [155,156]. In the middle years of the twentieth century, the traditional Okinawan diet was quite low in protein (9%), and especially animal protein (from just a small amount of fish—4% of calories); the Okinawans were subsequently characterized by having the highest proportion of centenarians in the world [157]. The Okinawans also practiced a modest degree of calorie restriction—an ethos that one should never completely fill one’s stomach, to ensure that enough food is available to all—which also could have enhanced their SIRT1 activity.
9. Spermidine for Hypusination of eIF5A
Spermidine, although produced in the body, can also be absorbed from the diet; the richest dietary source appears to be wheat germ. The extent to which post-translational modification of eIF5A to incorporate hypusine (hypusination) can be enhanced by supplemental spermidine needs further study; presumably, this will depend on baseline spermidine status, and might be of greater net benefit in the elderly who may have lesser spermidine tissue stores. Curiously, dietary spermidine content has been shown to correlate inversely with global mortality in a remarkable prospective epidemiological study [158]. Dietary spermidine supplementation conferred cardioprotection in mice subjected to salt-induced hypertension, an effect dependent on autophagy induction [159,160]. Administered in drinking water (5 mM per liter), spermidine has been shown to boost the functional capacity of T and B lymphocytes in aging mice by remedying an age-related reduction in autophagy; if translatable to humans, this effect might improve the natural resistance of the elderly to infections and cancer [39,40,41,161,162]. Spermidine has just recently become available as a nutraceutical (10 mg capsules; typical diets provide about 20 mg daily); the richest common food source appears to be wheat germ [163].
An independent mechanism whereby spermidine may support autophagy is by inhibition of the acetyltransferase EP300, which reduces the activity of certain proteins mediating autophagy via their acetylation [164]. Hence, spermidine complements the activity of SIRT1 in that regard.
10. Summary
To summarize, autophagy can be amplified by measures which enhance the activity of the kinase ULK1, and the transcription factors TFEB, FOXO1, ATF4 and CHOP. Measures which stimulate AMPK, SIRT1, and eIF5A, and down-regulate the activity of AKT and mTORC1 (and the growth factors which drive these activities) can be expected to have this effect. Berberine is a clinically useful AMPK activator. A number of nutraceuticals are emerging as of potential utility for boosting SIRT1 activity; these include ferulic acid, melatonin, MNA, urolithin A, NR, and glucosamine (Table 1). Ubiquinol administration may be helpful for this purpose when tissue ubiquinone levels are sub-optimal. Time-restricted feeding regimens may also promote episodic activation of SIRT1. Finally, plant-based diets of modest protein content may promote increased expression of ATF4 and CHOP, while blunting growth factor activity driven by IGF-I. Figure 1 depicts these relationships in graphic form.
It should be noted that activation of AMPK and SIRT1 may exert a number of additional health-protective effects, independent of their impact on autophagy. For one thing, acting primarily through support of PPARγ coactivator-1α activity (PGC-1α), they should promote mitochondrial biogenesis—an effect that is a necessary corrective when mitochondria are degraded by mitophagy [165]. SIRT1 also amplifies the activity of Nrf2 while suppressing the activity of NF-kappaB—accounting for the antioxidant and anti-inflammatory effects of SIRT1 activators [166,167,168]. SIRT1-mediated enhancement of the activities of KLF2 and of endothelial nitric oxide synthase in vascular endothelium can be expected to promote vascular health [169,170,171,172]. In the liver, SIRT1 activity decreases de novo lipogenesis by suppressing the activity of the transcription factor SREBP-1c, while AMPK up-regulates free fatty acid oxidation via inhibition of acetyl-CoA carboxylase—effects that are useful for management of non-alcoholic fatty liver disease [173,174]. Additionally, SIRT1 can lower LDL cholesterol by deacetylating and thereby suppressing hepatic secretion of PCSK9—a protease that binds to the external surface of hepatic LDL receptors and routes them to lysosomal degradation [175,176]. Although SIRT1’s ability to lessen p53 activation prompted by DNA damage has the potential to aid cancer promotion, this is compensated by SIRT’s ability to promote DNA repair, so that the net impact of SIRT1 activation on cancer risk remains equivocal [177].
With respect to the ability of low-protein plant-based diets to activate the GCN2-eIF2a-ATF4 pathway, this pathway, which enhances expression of FGF21 and adiponectin, may help to rationalize the lower risks for obesity, diabetes, cardiovascular disorders, autoimmunity and some “Western” types of cancer that have characterized some quasi-vegan societies [146,178].
Finally, it should be noted that, whereas moderate up-regulation of autophagy is often health protective, excessive autophagy, when not matched by compensatory anabolism, can lead to autophagic cell death [179,180,181]. Whereas up-regulation of autophagy appears to be beneficial in the early stages of various neurodegenerative diseases entailing accumulation of pathogenic protein complexes, it may be counterproductive in later-stage disease [182]. Additionally, some cancers employ elevated autophagy to enable them to survive in microenvironments low in oxygen and nutrients—pancreatic cancer is notable in this regard [183]. Hence, whether boosting autophagy is clinically worthwhile may hinge on the clinical context.
11. Dedication
This essay is dedicated to the memory of Jeremy Stone, a close friend of mine who was fascinated by the potential of up-regulated autophagy for promoting healthful longevity, and who encouraged me to pursue this topic. Jeremy, the eldest son of the great progressive journalist Izzy Stone, was President of the Federation of American Scientists for 30 years, under whose aegis he undertook private diplomacy throughout the world, aimed at preventing war and nuclear catastrophe. Perhaps the most celebrated of his many achievements was to conceive and enable the implementation of the Anti-Ballistic Missile Treaty. Jeremy, this one is for you!
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martinez-Lopez, N.; Athonvarangkul, D.; Singh, R. Autophagy and aging. Adv. Exp. Med. Biol. 2015, 847, 73–87. [Google Scholar] [PubMed] [Green Version]
- Ren, J.; Zhang, Y. Targeting Autophagy in Aging and Aging-Related Cardiovascular Diseases. Trends Pharmacol. Sci. 2018, 39, 1064–1076. [Google Scholar] [CrossRef] [PubMed]
- Bareja, A.; Lee, D.E.; White, J.P. Maximizing Longevity and Healthspan: Multiple Approaches All Converging on Autophagy. Front. Cell Dev. Biol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.; Tasset, I.; Arias, E.; Pampliega, O.; Wong, E.; Martinez-Vicente, M.; Cuervo, A.M. Autophagy and the hallmarks of aging. Ageing Res. Rev. 2021, 72, 101468. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Sintes, R.; Arias, E. Chaperone-mediated autophagy and disease: Implications for cancer and neurodegeneration. Mol. Asp. Med. 2021, 7, 101025. [Google Scholar] [CrossRef]
- Krause, G.J.; Cuervo, A.M. Assessment of mammalian endosomal microautophagy. Methods Cell Biol. 2021, 164, 167–185. [Google Scholar] [CrossRef]
- Ajoolabady, A.; Aslkhodapasandhokmabad, H.; Henninger, N.; Demillard, L.J.; Nikanfar, M.; Nourazarian, A.; Ren, J. Targeting autophagy in neurodegenerative diseases: From molecular mechanisms to clinical therapeutics. Clin. Exp. Pharmacol. Physiol. 2021, 48, 943–953. [Google Scholar] [CrossRef]
- Rana, T.; Behl, T.; Sehgal, A.; Mehta, V.; Singh, S.; Bhatia, S.; Al-Harrasi, A.; Bungau, S. Exploring the Role of Autophagy Dysfunction in Neurodegenerative Disorders. Mol. Neurobiol. 2021, 58, 4886–4905. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, X.; Song, Y.-Q.; Tu, J. Autophagy in Alzheimer’s disease pathogenesis: Therapeutic potential and future perspectives. Ageing Res. Rev. 2021, 72, 101464. [Google Scholar] [CrossRef]
- Zhang, H.; Puleston, D.J.; Simon, A.K. Autophagy and Immune Senescence. Trends Mol. Med. 2016, 22, 671–686. [Google Scholar] [CrossRef]
- Tanida, I. Autophagosome Formation and Molecular Mechanism of Autophagy. Antioxid. Redox Signal. 2011, 14, 2201–2214. [Google Scholar] [CrossRef] [PubMed]
- Zachari, M.; Ganley, I.G. The mammalian ULK1 complex and autophagy initiation. Essays Biochem. 2017, 61, 585–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, D.F.; Kim, J.; Shaw, R.J.; Guan, K.-L. The autophagy initiating kinase ULK1 is regulated via opposing phosphorylation by AMPK and mTOR. Autophagy 2011, 7, 643–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowans, G.J.; Hawley, S.A.; Ross, F.A.; Hardie, D.G. AMP Is a True Physiological Regulator of AMP-Activated Protein Kinase by Both Allosteric Activation and Enhancing Net Phosphorylation. Cell Metab. 2013, 18, 556–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Klionsky, D.J. AMPK-Dependent Phosphorylation of ULK1 Induces Autophagy. Cell Metab. 2011, 13, 119–120. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMPK and autophagy get connected. EMBO J. 2011, 30, 634–635. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Park, S.; Takahashi, Y.; Wang, H.-G. The Association of AMPK with ULK1 Regulates Autophagy. PLoS ONE 2010, 5, e15394. [Google Scholar] [CrossRef] [Green Version]
- Chan, E.Y. mTORC1 Phosphorylates the ULK1-mAtg13-FIP200 Autophagy Regulatory Complex. Sci. Signal. 2009, 2, pe51. [Google Scholar] [CrossRef]
- Inoki, K.; Zhu, T.; Guan, K.-L. TSC2 Mediates Cellular Energy Response to Control Cell Growth and Survival. Cell 2003, 115, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Gwinn, D.M.; Shackelford, D.B.; Egan, D.F.; Mihaylova, M.M.; Mery, A.; Vasquez, D.S.; Turk, B.E.; Shaw, R.J. AMPK Phosphorylation of Raptor Mediates a Metabolic Checkpoint. Mol. Cell 2008, 30, 214–226. [Google Scholar] [CrossRef] [Green Version]
- Van Nostrand, J.L.; Hellberg, K.; Luo, E.C.; Van Nostrand, E.L.; Dayn, A.; Yu, J.; Shokhirev, M.N.; Dayn, Y.; Yeo, G.W.; Shaw, R.J. AMPK regulation of Raptor and TSC2 mediate metformin effects on transcriptional control of anabolism and inflammation. Genes Dev. 2020, 34, 1330–1344. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.; Johnstone, S.R.; Dickerson, K.; Leiper, F.C.; Neumann, D.; Schlattner, U.; Wallimann, T.; Carlson, M. LKB1 is the upstream kinase in the AMP-activated protein kinase cascade. Curr. Biol. 2003, 13, 2004–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, F.; Cacicedo, J.M.; Ruderman, N.; Ido, Y. SIRT1 modulation of the acetylation status, cytosolic localization, and activity of LKB1. Possible role in AMP-activated protein kinase activation. J. Biol. Chem. 2008, 283, 27628–27635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napolitano, G.; Ballabio, A. TFEB at a glance. J. Cell Sci. 2016, 129, 2475–2481. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, M.; Impey, S.; Kang, H.; di Ronza, A.; Pelz, C.; Sardiello, M.; Ballabio, A. Characterization of the CLEAR network reveals an integrated control of cellular clearance pathways. Hum. Mol. Genet. 2011, 20, 3852–3866. [Google Scholar] [CrossRef] [Green Version]
- Martina, J.A.; Chen, Y.; Gucek, M.; Puertollano, R. MTORC1 functions as a transcriptional regulator of autophagy by preventing nuclear transport of TFEB. Autophagy 2012, 8, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Roczniak-Ferguson, A.; Petit, C.S.; Froehlich, F.; Qian, S.; Ky, J.; Angarola, B.; Walther, T.C.; Ferguson, S.M. The Transcription Factor TFEB Links mTORC1 Signaling to Transcriptional Control of Lysosome Homeostasis. Sci. Signal. 2012, 5, ra42. [Google Scholar] [CrossRef] [Green Version]
- Diego L Medina, D.L.; Di Paola, S.; Peluso, I.; Armani, A.; De Stefani, D. Venditti, R.; Montefusco, S.; Scotto-Rosato A., Prezioso, C.; Forrester, A. Lysosomal calcium signalling regulates autophagy through calcineurin and ​TFEB. Nat. Cell Biol. 2015, 17, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Mosquera, L.; Yambire, K.F.; Couto, R.; Pereyra, L.; Pabis, K.; Ponsford, A.H.; Diogo, C.V.; Stagi, M.; Milosevic, I.; Raimundo, N. Mitochondrial respiratory chain deficiency inhibits lysosomal hydrolysis. Autophagy 2019, 15, 1572–1591. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lai, Y.-C.; Hill, E.V.; Tyteca, D.; Carpentier, S.; Ingvaldsen, A.; Vertommen, D.; Lantier, L.; Foretz, M.; Dequiedt, F.; et al. Phosphatidylinositol 3-phosphate 5-kinase (PIKfyve) is an AMPK target participating in contraction-stimulated glucose uptake in skeletal muscle. Biochem. J. 2013, 455, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Zolov, S.N.; Bridges, D.; Zhang, Y.; Lee, W.-W.; Riehle, E.; Verma, R.; Lenk, G.M.; Converso-Baran, K.; Weide, T.; Albin, R.L.; et al. In vivo, Pikfyve generates PI(3,5)P2, which serves as both a signaling lipid and the major precursor for PI5P. Proc. Natl. Acad. Sci. USA 2012, 109, 17472–17477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, X.; Xu, H. Phosphoinositide isoforms determine compartment-specific ion channel activity. Proc. Natl. Acad. Sci. USA 2012, 109, 11384–11389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fine, M.; Schmiege, P.; Li, X. Structural basis for PtdInsP2-mediated human TRPML1 regulation. Nat. Commun. 2018, 9, 4192. [Google Scholar] [CrossRef] [PubMed]
- Paquette, M.; El-Houjeiri, L.; Zirden, L.C.; Puustinen, P.; Blanchette, P.; Jeong, H.; Dejgaard, K.; Siegel, P.M.; Pause, A. AMPK-dependent phosphorylation is required for transcriptional activation of TFEB and TFE3. Autophagy 2021, 17, 3957–3975. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Zhou, Z.; Park, J.-E.; Wang, L.; Wu, S.; Sun, X.; Lu, L.; Wang, T.; Lin, Q.; et al. Importance of TFEB acetylation in control of its transcriptional activity and lysosomal function in response to histone deacetylase inhibitors. Autophagy 2018, 14, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Bao, J.; Zheng, L.; Zhang, Q.; Li, X.; Zhang, X.; Li, Z.; Bai, X.; Zhang, Z.; Huo, W.; Zhao, X.; et al. Deacetylation of TFEB promotes fibrillar Aβ degradation by upregulating lysosomal biogenesis in microglia. Protein Cell 2016, 7, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Kou, J.; Wang, P.; Ye, T.; Wang, Z.; Gao, Z.; Cong, L.; Li, M.; Dong, B.; Yang, W.; et al. Berberine-induced TFEB deacetylation by SIRT1 promotes autophagy in peritoneal macrophages. Aging 2021, 13, 7096–7119. [Google Scholar] [CrossRef]
- Wolff, E.C.; Kang, K.R.; Kim, Y.S.; Park, M.H. Posttranslational synthesis of hypusine: Evolutionary progression and specificity of the hypusine modification. Amino Acids 2007, 33, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Alsaleh, G.; Feltham, J.; Sun, Y.; Napolitano, G.; Riffelmacher, T.; Charles, P.; Frau, L.; Hublitz, P.; Yu, Z.; et al. Polyamines Control eIF5A Hypusination, TFEB Translation, and Autophagy to Reverse B Cell Senescence. Mol. Cell 2019, 76, 110–125.e9. [Google Scholar] [CrossRef] [Green Version]
- Metur, S.P.; Klionsky, D.J. The curious case of polyamines: Spermidine drives reversal of B cell senescence. Autophagy 2020, 16, 389–390. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.; Ruhnau, J.; Schulze, J.; Obst, D.; Flöel, A.; Vogelgesang, A. Spermine and spermidine modulate T-cell function in older adults with and without cognitive decline ex vivo. Aging 2020, 12, 13716–13739. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, D.T.; Kaufman, R.J. All roads lead to ATF4. Dev Cell 2003, 4, 442–444. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Brewer, J.W.; Diehl, J.A.; Hendershot, L.M. Two Distinct Stress Signaling Pathways Converge Upon the CHOP Promoter During the Mammalian Unfolded Protein Response. J. Mol. Biol. 2002, 318, 1351–1365. [Google Scholar] [CrossRef]
- B'chir, W.; Maurin, A.C.; Carraro, V.; Averous, J.; Jousse, C.; Muranishi, Y.; Parry, L.; Stepien, G.; Fafournoux, P.; Bruhat, A. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res 2013, 41, 7683–7699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzivion, G.; Dobson, M.; Ramakrishnan, G. FoxO transcription factors; Regulation by AKT and 14-3-3 proteins. Biochim. Biophys. Acta 2011, 1813, 1938–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hariharan, N.; Maejima, Y.; Nakae, J.; Paik, J.; DePinho, R.; Sadoshima, J. Deacetylation of FoxO by Sirt1 Plays an Essential Role in Mediating Starvation-Induced Autophagy in Cardiac Myocytes. Circ. Res. 2010, 107, 1470–1482. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yang, Q.; Sun, Y.; Xing, Y.; Wang, Y.; Lu, X.; Bai, W.; Liu, X.; Zhao, Y. Resveratrol-enhanced autophagic flux ameliorates myocardial oxidative stress injury in diabetic mice. J. Cell. Mol. Med. 2014, 18, 1599–1611. [Google Scholar] [CrossRef]
- Gutierrez, M.G.; Munafo, D.B.; Beron, W.; Colombo, M.I. Rab7 is required for the normal progression of the autophagic pathway in mammalian cells. J. Cell Sci. 2004, 117, 2687–2697. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Wang, W.; Lu, Z.; Liu, Z.; Zhou, W.; Zhong, Z.; Ye, Q. Mild hypothermia pretreatment extenuates liver ischemia-reperfusion injury through Rab7-mediated autophagosomes-lysosomes fusion. Biochem. Biophys. Res. Commun. 2021, 550, 15–21. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y. AMPK and Autophagy. Adv. Exp. Med. Biol. 2019, 1206, 85–108. [Google Scholar]
- Tamargo-Go³mez, I.; Marino, G. AMPK: Regulation of Metabolic Dynamics in the Context of Autophagy. Int. J. Mol. Sci. 2018, 19, 3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Guan, K.L. AMPK connects energy stress to PIK3C3/VPS34 regulation. Autophagy 2013, 9, 1110–1111. [Google Scholar] [CrossRef] [Green Version]
- Jaber, N.; Dou, Z.; Chen, J.-S.; Catanzaro, J.; Jiang, Y.-P.; Ballou, L.M.; Selinger, E.; Ouyang, X.; Lin, R.Z.; Zhang, J.; et al. Class III PI3K Vps34 plays an essential role in autophagy and in heart and liver function. Proc. Natl. Acad. Sci. USA 2012, 109, 2003–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, A.M.; Csibi, A.; Raibon, A.; Cornille, K.; Gay, S.; Bernardi, H.; Candau, R. AMPK promotes skeletal muscle autophagy through activation of forkhead FoxO3a and interaction with Ulk1. J. Cell. Biochem. 2011, 113, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Lv, J.; Zeng, Q.L.; Shen, S.; Xing, J.Y.; Zhang, Y.Y.; Zhang, Z.H.; Yu, Z.J. AMPK activation ameliorates D-GalN/LPS-induced acute liver failure by upregulating Foxo3A to induce autophagy. Exp. Cell Res. 2017, 358, 335–342. [Google Scholar] [CrossRef]
- Sakamaki, J.-I.; Ryan, K.M. Transcriptional regulation of autophagy and lysosomal function by bromodomain protein BRD4. Autophagy 2017, 13, 2006–2007. [Google Scholar] [CrossRef] [Green Version]
- Ng, F.; Tang, B.L. Sirtuins’ modulation of autophagy. J. Cell. Physiol. 2013, 228, 2262–2270. [Google Scholar] [CrossRef]
- Sacitharan, P.K.; Bou-Gharios, G.; Edwards, J.R. SIRT1 directly activates autophagy in human chondrocytes. Cell Death Discov. 2020, 6, 41. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, W.S.; Kim, K.H.; Yoon, M.J.; Cho, H.J.; Shen, Y.; Ye, J.-M.; Lee, C.H.; Oh, W.K.; Kim, C.T.; et al. Berberine, a Natural Plant Product, Activates AMP-Activated Protein Kinase With Beneficial Metabolic Effects in Diabetic and Insulin-Resistant States. Diabetes 2006, 55, 2256–2264. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.S.; Lee, Y.S.; Cha, S.H.; Jeong, H.W.; Choe, S.S.; Lee, M.-R.; Oh, G.T.; Park, H.-S.; Lee, K.-U.; Lane, M.D.; et al. Berberine improves lipid dysregulation in obesity by controlling central and peripheral AMPK activity. Am. J. Physiol. Metab. 2009, 296, E812–E819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, N.; Li, J.L.; Gosby, A.; To, S.W.; Cheng, Z.; Miyoshi, H.; Taketo, M.M.; Cooney, G.J.; Kraegen, E.W.; James, D.E.; et al. Berberine and its more biologically available derivative, dihydroberberine, inhibit mitochondrial respiratory complex I: A mechanism for the action of berberine to activate AMP-activated protein kinase and improve insulin action. Diabetes 2008, 57, 1414–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Wang, N.; Zhao, L.; Lu, F. Berberine in the Treatment of Type 2 Diabetes Mellitus: A Systemic Review and Meta-Analysis. Evid.-Based Complement. Altern. Med. 2012, 2012, 591654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Xu, X.; Yin, M.; Zhang, Y.; Huang, L.; Chen, R.; Ni, J. Effects of berberine on blood glucose in patients with type 2 diabetes mellitus: A systematic literature review and a meta-analysis. Endocr. J. 2019, 66, 51–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, S.A.; Ross, F.A.; Chevtzoff, C.; Green, K.A.; Evans, A.; Fogarty, S.; Towler, M.C.; Brown, L.J.; Ogunbayo, O.A.; Evans, A.M.; et al. Use of cells expressing gamma subunit variants to identify diverse mechanisms of AMPK activation. Cell Metab. 2010, 11, 554–565. [Google Scholar] [CrossRef] [Green Version]
- Li, M.-H.; Zhang, Y.-J.; Yu, Y.-H.; Yang, S.-H.; Iqbal, J.; Mi, Q.-Y.; Li, B.; Wang, Z.-M.; Mao, W.-X.; Xie, H.-G.; et al. Berberine improves pressure overload-induced cardiac hypertrophy and dysfunction through enhanced autophagy. Eur. J. Pharmacol. 2014, 728, 67–76. [Google Scholar] [CrossRef]
- Wang, J.; Qi, Q.; Feng, Z.; Zhang, X.; Huang, B.; Chen, A.; Prestegarden, L.; Li, X.; Wang, J. Berberine induces autophagy in glioblastoma by targeting the AMPK/mTOR/ULK1-pathway. Oncotarget 2016, 7, 66944–66958. [Google Scholar] [CrossRef] [Green Version]
- Mohammadinejad, R.; Ahmadi, Z.; Tavakol, S.; Ashrafizadeh, M. Berberine as a potential autophagy modulator. J. Cell. Physiol. 2019, 234, 14914–14926. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Wang, Q.; Zhou, X.; Lu, X.; Liu, T.; Zhan, Y.; Li, P. Chinese Herbal Medicine in Ameliorating Diabetic Kidney Disease via Activating Autophagy. J. Diabetes Res. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Bailey, C.J. Metformin: Historical overview. Diabetology 2017, 60, 1566–1576. [Google Scholar] [CrossRef] [Green Version]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, B.P.; Gomes, A.P.; Dai, H.; Li, J.; Case, A.W.; Considine, T.; Riera, T.V.; Lee, J.E.; Yen, E.S.; Lamming, D.W.; et al. Evidence for a Common Mechanism of SIRT1 Regulation by Allosteric Activators. Science 2013, 339, 1216–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeberlein, M.; McDonagh, T.; Heltweg, B.; Hixon, J.; Westman, E.A.; Caldwell, S.D.; Napper, A.; Curtis, R.; DiStefano, P.S.; Fields, S.; et al. Substrate-specific Activation of Sirtuins by Resveratrol. J. Biol. Chem. 2005, 280, 17038–17045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacholec, M.; Bleasdale, J.E.; Chrunyk, B.; Cunningham, D.; Flynn, D.; Garofalo, R.S.; Griffith, D.; Griffor, M.; Loulakis, P.; Pabst, B.; et al. SRT1720, SRT2183, SRT1460, and Resveratrol Are Not Direct Activators of SIRT1. J. Biol. Chem. 2010, 285, 8340–8351. [Google Scholar] [CrossRef] [Green Version]
- Chimento, A.; De Amicis, F.; Sirianni, R.; Sinicropi, M.S.; Puoci, F.; Casaburi, I.; Saturnino, C.; Vincenzo Pezzi, V. Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 1381. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.B.; Hughes, M.C.; Edgett, B.A.; Scribbans, T.D.; Simpson, C.A.; Perry, C.G.; Gurd, B.J. An examination of resveratrol’s mechanisms of action in human tissue: Impact of a single dose in vivo and dose responses in skeletal muscle ex vivo. PLoS ONE 2014, 9, e102406. [Google Scholar] [CrossRef]
- Fulco, M.; Cen, Y.; Zhao, P.; Hoffman, E.P.; McBurney, M.W.; Sauve, A.A.; Sartorelli, V. Glucose Restriction Inhibits Skeletal Myoblast Differentiation by Activating SIRT1 through AMPK-Mediated Regulation of Nampt. Dev. Cell 2008, 14, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Costford, S.R.; Bajpeyi, S.; Pasarica, M.; Albarado, D.C.; Thomas, S.C.; Xie, H.; Church, T.S.; Jubrias, S.A.; Conley, K.E.; Smith, S.R. Skeletal muscle NAMPT is induced by exercise in humans. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E117–E126. [Google Scholar] [CrossRef] [Green Version]
- Revollo, J.R.; Grimm, A.A.; Imai, S. The NAD biosynthesis pathway mediated by nicotinamide phosphoribosyltransferase regulates Sir2 activity in mammalian cells. J. Biol. Chem. 2004, 279, 50754–50763. [Google Scholar] [CrossRef] [Green Version]
- Cantó, C.; Houtkooper, R.H.; Pirinen, E.; Youn, D.Y.; Oosterveer, M.H.; Cen, Y.; Fernandez-Marcos, P.J.; Yamamoto, H.; Andreux, P.A.; Cettour-Rose, P. The NAD(+) precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab. 2012, 15, 838–847. [Google Scholar] [CrossRef] [Green Version]
- Leduc-Gaudet, J.P.; Dulac, M.; Reynaud, O.; Ayoub, M.B.; Gouspillou, G. Nicotinamide riboside supplementation to improve skeletal muscle mitochondrial health and whole-body glucose homeostasis: Does it actually work in humans? J. Physiol. 2020, 598, 619–620. [Google Scholar] [CrossRef] [PubMed]
- Martens, C.R.; Denman, B.A.; Mazzo, M.R.; Armstrong, M.L.; Reisdorph, N.; McQueen, M.B.; Chonchol, M.; Seals, D.R. Chronic nicotinamide riboside supplementation is well-tolerated and elevates NAD(+) in healthy middle-aged and older adults. Nat. Commun. 2018, 9, 1286. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; DiNicolantonio, J.J. Coenzyme Q deficiency can be expected to compromise Sirt1 activity. Open Heart 2021, in press. [Google Scholar]
- Langsjoen, P.H.; Langsjoen, A.M. Supplemental ubiquinol in patients with advanced congestive heart failure. BioFactors 2008, 32, 119–128. [Google Scholar] [CrossRef]
- Xia, S.; Xu, S.; Zhang, X.; Zhong, F.; Wang, Z. Nanoliposomes Mediate Coenzyme Q10Transport and Accumulation across Human Intestinal Caco-2 Cell Monolayer. J. Agric. Food Chem. 2009, 57, 7989–7996. [Google Scholar] [CrossRef]
- García-Corzo, L.; Luna-Sánchez, M.; Doerrier, C.; Ortiz, F.; Escames, G.; Acuña-Castroviejo, D.; López, L.C. Ubiquinol-10 ameliorates mitochondrial encephalopathy associated with CoQ deficiency. Biochim. Biophys. Acta 2014, 1842, 893–901. [Google Scholar] [CrossRef] [Green Version]
- Langsjoen, P.H.; Langsjoen, A.M. Comparison study of plasma coenzyme Q10levels in healthy subjects supplemented with ubiquinol versus ubiquinone. Clin. Pharmacol. Drug Dev. 2014, 3, 13–17. [Google Scholar] [CrossRef]
- Mezawa, M.; Takemoto, M.; Onishi, S.; Ishibashi, R.; Ishikawa, T.; Yamaga, M.; Fujimoto, M.; Okabe, E.; He, P.; Kobayashi, K.; et al. The reduced form of coenzyme Q10 improves glycemic control in patients with type 2 diabetes: An open label pilot study. BioFactors 2012, 38, 416–421. [Google Scholar] [CrossRef]
- Sabbatinelli, J.; Orlando, P.; Galeazzi, R.; Silvestri, S.; Cirilli, I.; Marcheggiani, F.; Dludla, P.V.; Giuliani, A.; Bonfigli, A.R.; Mazzanti, L.; et al. Ubiquinol Ameliorates Endothelial Dysfunction in Subjects with Mild-to-Moderate Dyslipidemia: A Randomized Clinical Trial. Nutrients 2020, 12, 1098. [Google Scholar] [CrossRef] [Green Version]
- McCarty, M.F.; Assanga, S.B.I. Ferulic acid may target MyD88-mediated pro-inflammatory signaling—Implications for the health protection afforded by whole grains, anthocyanins, and coffee. Med. Hypotheses 2018, 118, 114–120. [Google Scholar] [CrossRef]
- Zhang, X.X.; Zhao, D.S.; Wang, J.; Zhou, H.; Wang, L.; Mao, J.L.; He, J.X. The treatment of cardiovascular diseases: A review of ferulic acid and its derivatives. Pharmazie 2021, 76, 55–60. [Google Scholar] [PubMed]
- El-Mesallamy, H.O.; Gawish, R.; Sallam, A.-A.M.; Fahmy, H.A.; Nada, A.S. Ferulic acid protects against radiation-induced testicular damage in male rats: Impact on SIRT1 and PARP1. Environ. Sci. Pollut. Res. 2017, 25, 6218–6227. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, F.H.; Mesbah-Ardakani, M.; Nasr-Esfahani, M.-H. Ferulic Acid exerts concentration-dependent anti-apoptotic and neuronal differentiation-inducing effects in PC12 and mouse neural stem cells. Eur. J. Pharmacol. 2018, 841, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Zhang, L.; Yang, X. Ferulic acid, a natural polyphenol, protects against osteoporosis by activating SIRT1 and NF-kappaB in neonatal rats with glucocorticoid-induced osteoporosis. Biomed. Pharm. 2019, 120, 109205. [Google Scholar] [CrossRef]
- Xu, T.; Song, Q.; Zhou, L.; Yang, W.; Wu, X.; Qian, Q.; Chai, H.; Han, Q.; Pan, H.; Dou, X.; et al. Ferulic acid alleviates lipotoxicity-induced hepatocellular death through the SIRT1-regulated autophagy pathway and independently of AMPK and Akt in AML-12 hepatocytes. Nutr. Metab. 2021, 18, 1–13. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; Chen, H.; Yu, J.; Luo, Y.; Zheng, P.; He, J. Effects of dietary ferulic acid supplementation on growth performance and skeletal muscle fiber type conversion in weaned piglets. J. Sci. Food Agric. 2021, 101, 5116–5123. [Google Scholar] [CrossRef]
- Du, K.; Fang, X.; Li, Z. Ferulic acid suppresses interleukin-1β-induced degeneration of chondrocytes isolated from patients with osteoarthritis through the SIRT1/AMPK/PGC-1α signaling pathway. Immun. Inflamm. Dis. 2021, 2, 710–720. [Google Scholar] [CrossRef]
- de Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharm. 2014, 171, 3268–3282. [Google Scholar] [CrossRef] [Green Version]
- Lillioja, S.; Neal, A.L.; Tapsell, L.; Jacobs, D.R., Jr. Whole grains, type 2 diabetes, coronary heart disease, and hypertension: Links to the aleurone preferred over indigestible fiber. BioFactors 2013, 39, 242–258. [Google Scholar] [CrossRef] [Green Version]
- Bumrungpert, A.; Lilitchan, S.; Tuntipopipat, S.; Tirawanchai, N.; Komindr, S. Ferulic Acid Supplementation Improves Lipid Profiles, Oxidative Stress, and Inflammatory Status in Hyperlipidemic Subjects: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2018, 10, 713. [Google Scholar] [CrossRef] [Green Version]
- Cerdá, B.; Periago, P.; Espín, A.J.C.; Tomás-Barberán, F.A. Identification of Urolithin A as a Metabolite Produced by Human Colon Microflora from Ellagic Acid and Related Compounds. J. Agric. Food Chem. 2005, 53, 5571–5576. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Henning, S.M.; Zhang, Y.; Suchard, M.; Li, Z.; Heber, D. Pomegranate Juice Ellagitannin Metabolites Are Present in Human Plasma and Some Persist in Urine for Up to 48 Hours. J. Nutr. 2006, 136, 2481–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Istas, G.; Feliciano, R.P.; Weber, T.; Villalba, R.G.; Tomas-Barberan, F.; Heiss, C.; Rodriguez-Mateos, A. Plasma urolithin metabolites correlate with improvements in endothelial function after red raspberry consumption: A double-blind randomized controlled trial. Arch. Biochem. Biophys. 2018, 651, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Asgary, S.; Javanmard, S.H.; Zarfeshany, A. Potent health effects of pomegranate. Adv. Biomed. Res. 2014, 3, 100. [Google Scholar] [CrossRef]
- Velagapudi, R.; Lepiarz, I.; El-Bakoush, A.; Katola, F.O.; Bhatia, H.; Fiebich, B.L.; Olajide, O.A. Induction of Autophagy and Activation of SIRT-1 Deacetylation Mechanisms Mediate Neuroprotection by the Pomegranate Metabolite Urolithin A in BV2 Microglia and Differentiated 3D Human Neural Progenitor Cells. Mol. Nutr. Food Res. 2019, 63, e1801237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, N.; Das, A.; Biswas, N.; Gnyawali, S.; Singh, K.; Gorain, M.; Polcyn, C.; Khanna, S.; Roy, S.; Sen, C.K. Urolithin A augments angiogenic pathways in skeletal muscle by bolstering NAD+ and SIRT1. Sci. Rep. 2020, 10, 20184. [Google Scholar] [CrossRef] [PubMed]
- Andreux, P.A.; Blanco-Bose, W.; Ryu, D.; Burdet, F.; Ibberson, M.; Aebischer, P.; Auwerx, J.; Singh, A.; Rinsch, C. The mitophagy activator urolithin A is safe and induces a molecular signature of improved mitochondrial and cellular health in humans. Nat. Metab. 2019, 1, 595–603. [Google Scholar] [CrossRef]
- Fang, J.; Yan, Y.; Teng, X.; Wen, X.; Li, N.; Peng, S.; Liu, W.; Donadeu, F.X.; Zhao, S.; Hua, J. Melatonin prevents senescence of canine adipose-derived mesenchymal stem cells through activating NRF2 and inhibiting ER stress. Aging 2018, 10, 2954–2972. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, C.; Meng, C.-J.; Zhu, G.-Q.; Sun, X.-B.; Huo, L.; Zhang, J.; Liu, H.-X.; He, W.-C.; Shen, X.-M.; et al. Melatonin activates the Nrf2-ARE pathway when it protects against early brain injury in a subarachnoid hemorrhage model. J. Pineal Res. 2012, 53, 129–137. [Google Scholar] [CrossRef]
- Cristòfol, R.; Porquet, D.; Corpas, R.; Coto-Montes, A.; Serret, J.; Camins, A.; Pallàs, M.; Sanfeliu, C. Neurons from senescence-accelerated SAMP8 mice are protected against frailty by the sirtuin 1 promoting agents melatonin and resveratrol. J. Pineal Res. 2012, 52, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Sun, Y.; Cheng, L.; Jin, Z.; Yang, Y.; Zhai, M.; Pei, H.; Wang, X.; Zhang, H.; Meng, Q.; et al. Melatonin receptor-mediated protection against myocardial ischemia/reperfusion injury: Role of SIRT1. J. Pineal Res. 2014, 57, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, S.; Dong, Y.; Fan, C.; Zhao, L.; Yang, X.; Li, J.; Di, S.; Yue, L.; Liang, G.; et al. Melatonin prevents cell death and mitochondrial dysfunction via a SIRT1-dependent mechanism during ischemic-stroke in mice. J. Pineal Res. 2015, 58, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zhang, Y.; Zhang, F.; Xia, Y.; Liu, J.; Huang, R.; Wang, Y.; Hu, Y.; Wu, J.; Dai, C.; et al. CLOCK/BMAL1 regulates circadian change of mouse hepatic insulin sensitivity by SIRT1. Hepatology 2014, 59, 2196–2206. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, S.; Szumlanski, C.L.; Weinshilboum, R.M. Human liver nicotinamide N-methyltransferase. cDNA cloning, expression, and biochemical characterization. J. Biol. Chem. 1994, 269, 14835–14840. [Google Scholar] [CrossRef]
- Gebicki, J.; Sysa-Jedrzejowska, A.; Adamus, J.; Woźniacka, A.; Rybak, M.; Zielonka, J. 1-Methylnicotinamide: A potent anti-inflammatory agent of vitamin origin. Pol. J. Pharmacol. 2003, 55, 109–112. [Google Scholar]
- Chen, Y.; Zhang, J.; Li, P.; Liu, C.; Li, L. N1‑methylnicotinamide ameliorates insulin resistance in skeletal muscle of type 2 diabetic mice by activating the SIRT1/PGC1-α signaling pathway. Mol. Med. Rep. 2021, 23, 270. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Y.; Liu, C.; Li, L.; Li, P. N1-Methylnicotinamide Improves Hepatic Insulin Sensitivity via Activation of SIRT1 and Inhibition of FOXO1 Acetylation. J. Diabetes Res. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Miwa, T. Protective Effects of N1-Methylnicotinamide Against High-Fat Diet- and Age-Induced Hearing Loss via Moderate Overexpression of Sirtuin 1 Protein. Front. Cell. Neurosci. 2021, 15, 634868. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Y.; Liu, Y.; Tang, W.-Q.; Ji, C.-H.; Gu, J.-H.; Jiang, B. Antidepressant-like effects of 1-methylnicotinamide in a chronic unpredictable mild stress model of depression. Neurosci. Lett. 2020, 742, 135535. [Google Scholar] [CrossRef]
- Nejabati, H.R.; Samadi, N.; Shahnazi, V.; Mihanfar, A.; Fattahi, A.; Latifi, Z.; Bahrami-Asl, Z.; Roshangar, L.; Nouri, M. Nicotinamide and its metabolite N1-Methylnicotinamide alleviate endocrine and metabolic abnormalities in adipose and ovarian tissues in rat model of Polycystic Ovary Syndrome. Chem. Interact. 2020, 324, 109093. [Google Scholar] [CrossRef]
- Nejabati, H.R.; Samadi, N.; Roshangar, L.; Nouri, M. N1-methylnicotinamide as a possible modulator of cardiovascular risk markers in polycystic ovary syndrome. Life Sci. 2019, 235, 116843. [Google Scholar] [CrossRef]
- Fu, L.; Liu, C.; Chen, L.; Lv, Y.; Meng, G.; Hu, M.; Long, Y.; Hong, H.; Tang, S. Protective Effects of 1-Methylnicotinamide on Aβ(1-42)-Induced Cognitive Deficits, Neuroinflammation and Apoptosis in Mice. J. Neuroimmune Pharm. 2019, 14, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Nejabati, H.R.; Mihanfar, A.; Pezeshkian, M.; Fattahi, A.; Latifi, Z.; Safaie, N.; Valilo, M.; Jodati, A.R.; Nouri, M. N1-methylnicotinamide (MNAM) as a guardian of cardiovascular system. J. Cell. Physiol. 2018, 233, 6386–6394. [Google Scholar] [CrossRef]
- Mateuszuk, L.; Jasztal, A.; Maslak, E.; Gasior-Glogowska, M.; Baranska, M.; Sitek, B.; Kostogrys, R.; Zakrzewska, A.; Kij, A.; Walczak, M.; et al. Antiatherosclerotic Effects of 1-Methylnicotinamide in Apolipoprotein E/Low-Density Lipoprotein Receptor-Deficient Mice: A Comparison with Nicotinic Acid. J. Pharmacol. Exp. Ther. 2016, 356, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Moreno-Navarrete, J.M.; Wei, X.; Kikukawa, Y.; Tzameli, I.; Prasad, D.; Lee, Y.; Asara, J.M.; Fernández-Real, J.M.; Maratos-Flier, E.; et al. Nicotinamide N-methyltransferase regulates hepatic nutrient metabolism through Sirt1 protein stabilization. Nat. Med. 2015, 21, 887–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campagna, R.; Mateuszuk, L.; Wojnar-Lason, K.; Kaczara, P.; Tworzydło, A.; Kij, A.; Bujok, R.; Mlynarski, J.; Wang, Y.; Sartini, D.; et al. Nicotinamide N-methyltransferase in endothelium protects against oxidant stress-induced endothelial injury. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119082. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Gu, Y.; Shan, H.; Wenyi Mi, W.; Sun, J.; Shi, M.; Zhang, X.; Lu, X.; Han, F.; Gong, Q.; et al. O-GlcNAcylation of SIRT1 enhances its deacetylase activity and promotes cytoprotection under stress. Nat. Commun. 2017, 8, 1491. [Google Scholar] [CrossRef]
- McCarty, M.F.; O’Keefe, J.H.; DiNicolantonio, J.J. Glucosamine for the Treatment of Osteoarthritis: The Time Has Come for Higher-Dose Trials. J. Diet. Suppl. 2018, 16, 179–192. [Google Scholar] [CrossRef]
- Weimer, S.; Priebs, J.; Kuhlow, D.; Groth, M.; Priebe, S.; Mansfeld, J.; Merry, T.L.; Dubuis, S.; Laube, B.; Pfeiffer, A.F.; et al. D-Glucosamine supplementation extends life span of nematodes and of ageing mice. Nat. Commun. 2014, 5, 3563. [Google Scholar] [CrossRef] [Green Version]
- Janssens, G.E.; Houtkooper, R.H. Identification of longevity compounds with minimized probabilities of side effects. Biogerontology 2020, 21, 709–719. [Google Scholar] [CrossRef]
- Bell, G.A.; Kantor, E.D.; Lampe, J.W.; Shen, D.D.; White, E. Use of glucosamine and chondroitin in relation to mortality. Eur. J. Epidemiol. 2012, 27, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Katoh, A.; Kai, H.; Harada, H.; Niiyama, H.; Ikeda, H. Oral Administration of Glucosamine Improves Vascular Endothelial Function by Modulating Intracellular Redox State. Int. Heart J. 2017, 58, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Mattagajasingh, I.; Kim, C.-S.; Naqvi, A.; Yamamori, T.; Hoffman, T.A.; Jung, S.-B.; DeRicco, J.; Kasuno, K.; Irani, K. SIRT1 promotes endothelium-dependent vascular relaxation by activating endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 14855–14860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalkiadaki, A.; Guarente, L. Sirtuins mediate mammalian metabolic responses to nutrient availability. Nat. Rev. Endocrinol. 2012, 8, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yan, Y.; Gius, D.R.; Vassilopoulos, A. Metabolic regulation of Sirtuins upon fasting and the implication for cancer. Curr. Opin. Oncol. 2013, 25, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Kitada, M.; Kume, S.; Takeda-Watanabe, A.; Tsuda, S.-I.; Kanasaki, K.; Koya, D. Calorie restriction in overweight males ameliorates obesity-related metabolic alterations and cellular adaptations through anti-aging effects, possibly including AMPK and SIRT1 activation. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 4820–4827. [Google Scholar] [CrossRef]
- Jamshed, H.; Beyl, R.A.; Della Manna, D.L.; Yang, E.S.; Ravussin, E.; Peterson, C.M. Early time-restricted feeding improves 24-hour glucose levels and affects markers of the circadian clock, aging, and autophagy in humans. Nutrients 2019, 11, 1234. [Google Scholar] [CrossRef] [Green Version]
- McCarty, M.F. A note on “orthomolecular aids for dieting”—Myasthenic syndrome due to dl-carnitine. Med. Hypotheses 1982, 9, 661–662. [Google Scholar] [CrossRef]
- McCarty, M.F.; DiNicolantonio, J.J.; O’keefe, J.H. Ketosis may promote brain macroautophagy by activating Sirt1 and hypoxia-inducible factor-1. Med. Hypotheses 2015, 85, 631–639. [Google Scholar] [CrossRef]
- Hoffman, W.H.; Shacka, J.J.; Andjelkovic, A.V. Autophagy in the brains of young patients with poorly controlled T1DM and fatal diabetic ketoacidosis. Exp. Mol. Pathol. 2011, 93, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Liśkiewicz, D.; Liśkiewicz, A.; Grabowski, M.; Nowacka-Chmielewska, M.M.; Jabłońska, K.; Wojakowska, A.; Marczak, L.; Barski, J.J.; Małecki, A. Upregulation of hepatic autophagy under nutritional ketosis. J. Nutr. Biochem. 2021, 93, 108620. [Google Scholar] [CrossRef] [PubMed]
- Montiel, T.; Montes-Ortega, L.A.; Flores-Yáñez, S.; Massieu, L. Treatment with the Ketone Body D-β-hydroxybutyrate Attenuates Autophagy Activated by NMDA and Reduces Excitotoxic Neuronal Damage in the Rat Striatum In Vivo. Curr. Pharm. Des. 2020, 26, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Jiang, L.Q.; Deshmukh, A.S.; Mataki, C.; Coste, A.; Lagouge, M.; Zierath, J.R.; Auwerx, J. Interdependence of AMPK and SIRT1 for metabolic adaptation to fasting and exercise in skeletal muscle. Cell Metab. 2010, 11, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Wang, X.; Zhu, Y.; Li, Z.; Zhu, Y.-T.; Wu, J.-C.; Qin, Z.-H.; Xiang, M.; Lin, F. Exercise activates lysosomal function in the brain through AMPK-SIRT1-TFEB pathway. CNS Neurosci. Ther. 2019, 25, 796–807. [Google Scholar] [CrossRef]
- White, A.T.; Schenk, S. NAD(+)/NADH and skeletal muscle mitochondrial adaptations to exercise. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E308–E321. [Google Scholar] [CrossRef] [Green Version]
- McCarty, M.F. GCN2 and FGF21 are likely mediators of the protection from cancer, autoimmunity, obesity, and diabetes afforded by vegan diets. Med. Hypotheses 2014, 83, 365–371. [Google Scholar] [CrossRef]
- Dong, J.; Qiu, H.; Garcia-Barrio, M.; Anderson, J.; Hinnebusch, A.G. Uncharged tRNA Activates GCN2 by Displacing the Protein Kinase Moiety from a Bipartite tRNA-Binding Domain. Mol. Cell 2000, 6, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Masson, G.R. Towards a model of GCN2 activation. Biochem. Soc. Trans. 2019, 47, 1481–1488. [Google Scholar] [CrossRef] [Green Version]
- De Sousa-Coelho, A.L.; Marrero, P.F.; Haro, D. Activating transcription factor 4-dependent induction of FGF21 during amino acid deprivation. Biochem. J. 2012, 443, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, T.; Lin, V.Y.; Goetz, R.; Mohammadi, M.; Mangelsdorf, D.; Kliewer, S.A. Inhibition of Growth Hormone Signaling by the Fasting-Induced Hormone FGF21. Cell Metab. 2008, 8, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Fontana, L.; Weiss, E.P.; Villareal, D.T.; Klein, S.; Holloszy, J.O. Long-term effects of calorie or protein restriction on serum IGF-1 and IGFBP-3 concentration in humans. Aging Cell 2008, 7, 681–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castaño-Martinez, T.; Schumacher, F.; Schumacher, S.; Kochlik, B.; Weber, D.; Grune, T.; Biemann, R.; McCann, A.; Abraham, K.; Weikert, C. Methionine restriction prevents onset of type 2 diabetes in NZO mice. FASEB J. 2019, 33, 7092–7102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, Y.; Berglund, E.D.; Coate, K.; He, T.T.; Katafuchi, T.; Xiao, G.; Potthoff, M.; Wei, W.; Wan, Y.; et al. The starvation hormone, fibroblast growth factor-21, extends lifespan in mice. eLife 2012, 1, e00065. [Google Scholar] [CrossRef] [PubMed]
- Bartke, A. Minireview: Role of the Growth Hormone/Insulin-Like Growth Factor System in Mammalian Aging. Endocrinology 2005, 146, 3718–3723. [Google Scholar] [CrossRef] [Green Version]
- Bartke, A. Healthy Aging: Is Smaller Better?—A Mini-Review. Gerontology 2012, 58, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Patronek, G.J.; Waters, D.J.; Glickman, L.T. Comparative Longevity of Pet Dogs and Humans: Implications for Gerontology Research. J. Gerontol. Ser. A 1997, 52, B171–B178. [Google Scholar] [CrossRef] [Green Version]
- Willcox, B.J.; Willcox, D.C.; Todoriki, H.; Fujiyoshi, A.; Yano, K.; He, Q.; Curb, J.D.; Suzuki, M. Caloric restriction, the traditional Okinawan diet, and healthy aging: The diet of the world’s longest-lived people and its potential impact on morbidity and life span. Ann. N. Y. Acad. Sci. 2007, 1114, 434–455. [Google Scholar] [CrossRef] [Green Version]
- Kiechl, S.; Pechlaner, R.; Willeit, P.; Notdurfter, M.; Paulweber, B.; Willeit, K.; Werner, P.; Ruckenstuhl, C.; Iglseder, B.; Weger, S.; et al. Higher spermidine intake is linked to lower mortality: A prospective population-based study. Am. J. Clin. Nutr. 2018, 108, 371–380. [Google Scholar] [CrossRef]
- Eisenberg, T.; Abdellatif, M.; Schroeder, S.; Primessnig, U.; Stekovic, S.; Pendl, T.; Harger, A.; Schipke, J.; Zimmermann, A.; Schmidt, A.; et al. Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat. Med. 2016, 22, 1428–1438. [Google Scholar] [CrossRef]
- Eisenberg, T.; Abdellatif, M.; Zimmermann, A.; Schroeder, S.; Pendl, T.; Harger, A.; Stekovic, S.; Schipke, J.; Magnes, C.; Schmidt, A.; et al. Dietary spermidine for lowering high blood pressure. Autophagy 2017, 13, 767–769. [Google Scholar] [CrossRef]
- Puleston, D.J.; Zhang, H.; Powell, T.J.; Lipina, E.; Sims, S.; Panse, I.; Watson, A.S.; Cerundolo, V.; Townsend, A.; Rm, T.A.; et al. Autophagy is a critical regulator of memory CD8+ T cell formation. eLife 2014, 3, e03706. [Google Scholar] [CrossRef] [PubMed]
- Alsaleh, G.; Panse, I.; Swadling, L.; Zhang, H.; Richter, F.; Meyer, A.; Lord, J.; Barnes, E.; Klenerman, P.; Green, C.; et al. Autophagy in T cells from aged donors is maintained by spermidine, and correlates with function and vaccine responses. eLife 2020, 9, e57950. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Esparza, N.C.; Costa-Catala, J.; Comas-Baste, O.; Toro-Funes, N.; Latorre-Moratalla, M.L.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Occurrence of Polyamines in Foods and the Influence of Cooking Processes. Foods 2021, 10, 1752. [Google Scholar] [CrossRef] [PubMed]
- Pietrocola, F.; Lachkar, S.; Enot, D.P.; Niso-Santano, M.; Pedro, J.M.B.-S.; Sica, V.; Izzo, V.; Maiuri, M.C.; Madeo, F.; Mariño, G.; et al. Spermidine induces autophagy by inhibiting the acetyltransferase EP300. Cell Death Differ. 2014, 22, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Petrocelli, J.J.; Drummond, M.J. PGC-1α-Targeted Therapeutic Approaches to Enhance Muscle Recovery in Aging. Int. J. Environ. Res. Public Health 2020, 17, 8650. [Google Scholar] [CrossRef]
- Ding, Y.-W.; Zhao, G.-J.; Li, X.-L.; Hong, G.-L.; Li, M.-F.; Qiu, Q.-M.; Wu, B.; Lu, Z.-Q. SIRT1 exerts protective effects against paraquat-induced injury in mouse type II alveolar epithelial cells by deacetylating NRF2 in vitro. Int. J. Mol. Med. 2016, 37, 1049–1058. [Google Scholar] [CrossRef]
- Salminen, A.; Kauppinen, A.; Suuronen, T.; Kaarniranta, K. SIRT1 longevity factor suppresses NF-kappaB -driven immune responses: Regulation of aging via NF-kappaB acetylation? Bioessays 2008, 30, 939–942. [Google Scholar] [CrossRef]
- Yeung, F.; Hoberg, J.E.; Ramsey, C.S.; Keller, M.D.; Jones, D.R.; Frye, R.A.; Mayo, M.W. Modulation of NF-kappaB-dependent transcription and cell survival by the SIRT1 deacetylase. EMBO J. 2004, 23, 2369–2380. [Google Scholar] [CrossRef] [Green Version]
- Gracia-Sancho, J.; Villarreal, G., Jr.; Zhang, Y.; García-Cardeña, G. Activation of SIRT1 by resveratrol induces KLF2 expression conferring an endothelial vasoprotective phenotype. Cardiovasc. Res. 2009, 85, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Liu, X.; Feng, H.; Zhao, S.; Gao, H. Grape seed proanthocyanidin extracts enhance endothelial nitric oxide synthase expression through 5’-AMP activated protein kinase/Surtuin 1-Krüpple like factor 2 pathway and modulate blood pressure in ouabain induced hypertensive rats. Biol. Pharm. Bull. 2012, 35, 2192–2197. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yang, W.; Dai, H.; Deng, Z. Cardiovascular risk following metformin treatment in patients with type 2 diabetes mellitus: Results from meta-analysis. Diabetes Res. Clin. Pract. 2020, 160, 108001. [Google Scholar] [CrossRef] [PubMed]
- Monami, M.; Candido, R.; Pintaudi, B.; Targher, G.; Mannucci, E.; Monache, L.D.; Gallo, M.; Giaccari, A.; Masini, M.L.; Mazzone, F.; et al. Effect of metformin on all-cause mortality and major adverse cardiovascular events: An updated meta-analysis of randomized controlled trials. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Ponugoti, B.; Kim, D.-H.; Xiao, Z.; Smith, Z.; Miao, J.; Zang, M.; Wu, S.-Y.; Chiang, C.-M.; Veenstra, T.D.; Kemper, J.K. SIRT1 Deacetylates and Inhibits SREBP-1C Activity in Regulation of Hepatic Lipid Metabolism. J. Biol. Chem. 2010, 285, 33959–33970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Chen, X.; Xue, H.; Zhang, P.; Fang, W.; Chen, X.; Ling, W. Coenzyme Q10 attenuates high-fat diet-induced non-alcoholic fatty liver disease through activation of the AMPK pathway. Food Funct. 2019, 10, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.X.; van Tits, L.J.; Lohmann, C.; Arsiwala, T.; Winnik, S.; Tailleux, A.; Stein, S.; Gomes, A.P.; Suri, V.; Ellis, J.L. The Sirt1 activator SRT3025 provides atheroprotection in Apoe−/− mice by reducing hepatic Pcsk9 secretion and enhancing Ldlr expression. Eur. Heart J. 2015, 36, 51–59. [Google Scholar] [CrossRef]
- Wang, H.; He, F.; Liang, B.; Jing, Y.; Zhang, P.; Liu, W.; Zhu, B.; Dou, D. LincRNA-p21 alleviates atherosclerosis progression through regulating the miR-221/SIRT1/Pcsk9 axis. J. Cell Mol. Med. 2021, 25, 9141–9153. [Google Scholar] [CrossRef]
- Garcia-Peterson, L.M.; Li, X. Trending topics of SIRT1 in tumorigenicity. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129952. [Google Scholar] [CrossRef]
- Mccarty, M.F. The moderate essential amino acid restriction entailed by low-protein vegan diets may promote vascular health by stimulating FGF21 secretion. Horm. Mol. Biol. Clin. Investig. 2016, 30. [Google Scholar] [CrossRef]
- Noguchi, M.; Hirata, N.; Tanaka, T.; Suizu, F.; Nakajima, H.; Chiorini, J.A. Autophagy as a modulator of cell death machinery. Cell Death Dis. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Ikeda, S.; Zablocki, D.; Sadoshima, J. The role of autophagy in death of cardiomyocytes. J. Mol. Cell. Cardiol. 2021, 165, 1–8. [Google Scholar] [CrossRef]
- Button, R.W.; Luo, S.; Rubinsztein, D.C. Autophagic activity in neuronal cell death. Neurosci. Bull. 2015, 31, 382–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xilouri, M.; Stefanis, L. Autophagy in the central nervous system: Implications for neurodegenerative disorders. CNS Neurol. Disord. Drug Targets 2010, 9, 701–719. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Mitsunaga, S.; Yamazaki, M.; Hasebe, T.; Ishii, G.; Kojima, M.; Kinoshita, T.; Ueno, T.; Esumi, H.; Ochiai, A. Autophagy is activated in pancreatic cancer cells and correlates with poor patient outcome. Cancer Sci. 2008, 99, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
How nutraceuticals and diet can activate key mediators of autophagy—the initiating catalyst of autophagosome formation ULK1, and the transcription factors TFEB, FOXO1, ATF4 and CHOP, which promote expression of many proteins participating in autophagy. Sirt1 can activate some of these proteins (ATG5, LC3, Beclin1, etc.) via deacetylation. Sirt1 activators can include ferulic acid, melatonin, N1-methylnicotinamide, urolithin A, nicotinamide riboside, coenzyme Q10, glucosamine, aerobic exercise, and time-restricted feeding or intermittent fasting.
Figure 1.
How nutraceuticals and diet can activate key mediators of autophagy—the initiating catalyst of autophagosome formation ULK1, and the transcription factors TFEB, FOXO1, ATF4 and CHOP, which promote expression of many proteins participating in autophagy. Sirt1 can activate some of these proteins (ATG5, LC3, Beclin1, etc.) via deacetylation. Sirt1 activators can include ferulic acid, melatonin, N1-methylnicotinamide, urolithin A, nicotinamide riboside, coenzyme Q10, glucosamine, aerobic exercise, and time-restricted feeding or intermittent fasting.


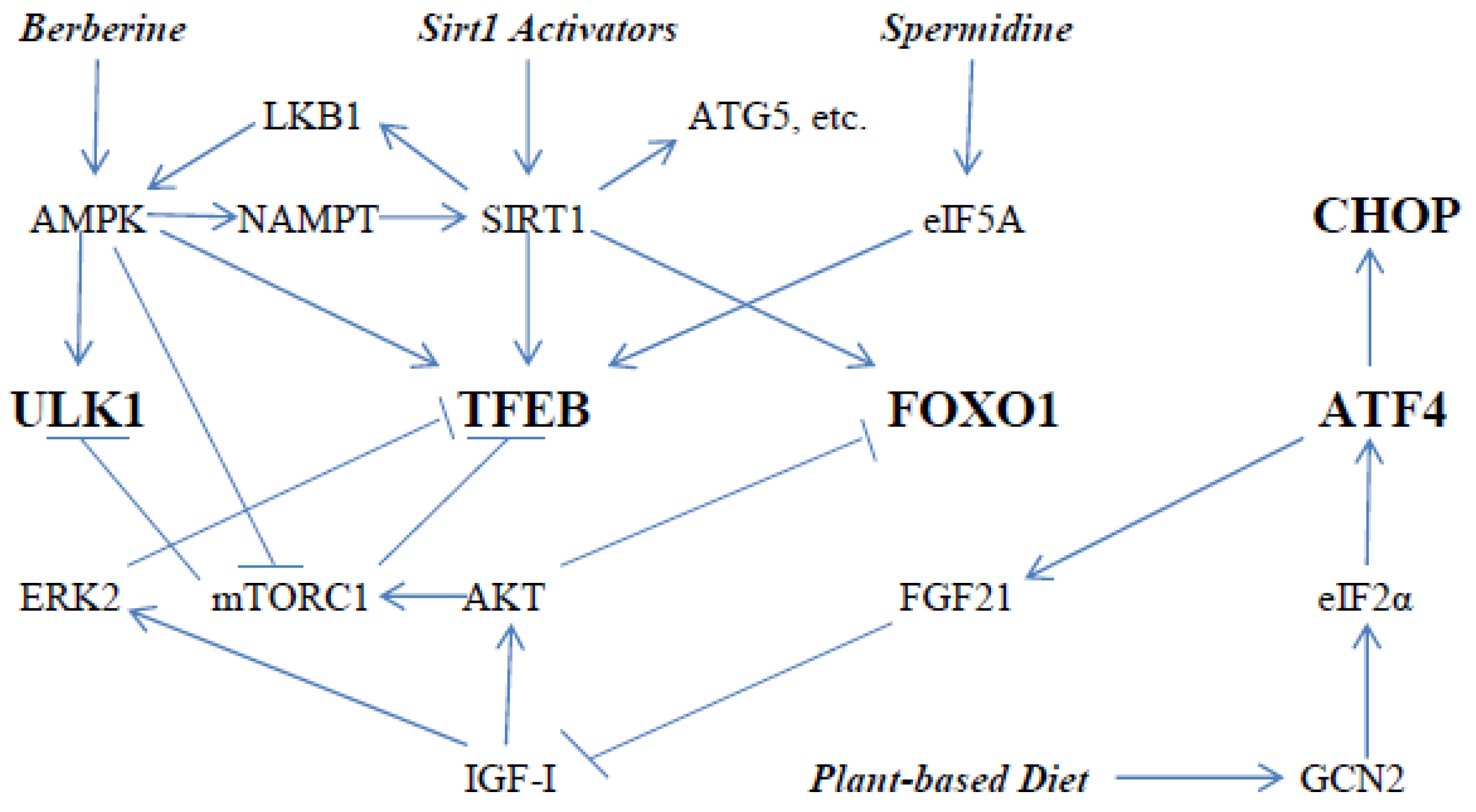{kind=link}
Table 1.
Nutraceuticals with potential for activation of autophagy.
| Nutraceutical | Target | Clinically Relevant Dosing * | References |
|---|---|---|---|
| Berberine | AMPK | 1000–2000 mg daily, divided doses | [60,61,62,63,64,65,66,67,68] |
| Ferulic Acid | Sirt1 | 500–1000 mg daily, divided doses | [85,86,87,88,89,90,91,92] |
| Melatonin | Sirt1 | 5–20 mg at bedtime | [104,105,106,107,108,109] |
| N1-methylnicotinamide | Sirt1 | Not established ** | [121] |
| Urolithin A | Sirt1 | Not established ** | [101,102,103] |
| Nicotinamide Riboside | Sirt1 | 500–2000 mg daily, divided doses | [75,76,77] |
| Coenzyme Q10 | Sirt1 | 100–300 mg as ubiquinol daily | [78] |
| Glucosamine | Sirt1 | 1.5–3 g once daily | [123,124] |
| Spermidine | EIF5A *** | 10–30 mg daily, divided doses **** | [39,40,41] |
* These are not represented as ideal doses for activation of the defined targets, but rather doses which in clinical experience have been sufficient to exert physiological effects. ** While these agents are produced in the human body and can be presumed to be safe at physiological levels, and have been made available as nutraceuticals, there is too little published clinical experience with them at present to define worthwhile clinical doses. *** Required for efficient translation of TFEB mRNA. **** Since natural diets provide a spermidine intake in the range of 20 mg daily, it is presumed that 10–30 mg of supplemental spermidine daily will meaningfully impact spermidine status.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
McCarty, M.F. Nutraceutical and Dietary Strategies for Up-Regulating Macroautophagy. Int. J. Mol. Sci. 2022, 23, 2054. https://doi.org/10.3390/ijms23042054
AMA Style
McCarty MF. Nutraceutical and Dietary Strategies for Up-Regulating Macroautophagy. International Journal of Molecular Sciences. 2022; 23(4):2054. https://doi.org/10.3390/ijms23042054
Chicago/Turabian StyleMcCarty, Mark F. 2022. "Nutraceutical and Dietary Strategies for Up-Regulating Macroautophagy" International Journal of Molecular Sciences 23, no. 4: 2054. https://doi.org/10.3390/ijms23042054
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.