
Tissue Transglutaminase but Not Microbial Transglutaminase Is Inhibited by Exogenous Oxidative Substances in Celiac Disease


Abstract
:1. Introduction
2. Results
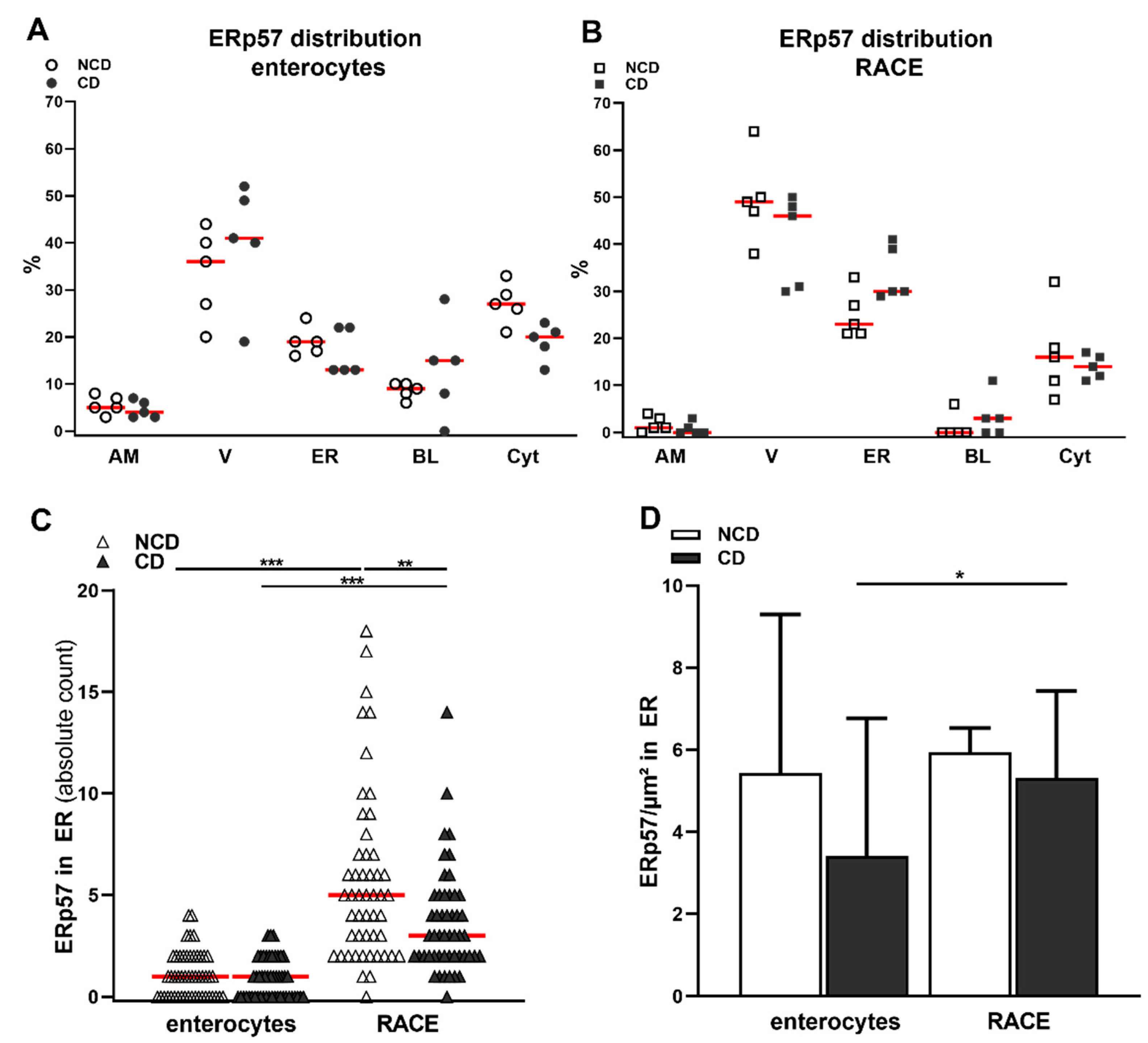
2.1. Localization of ERp57 within Intra- and Extracellular Compartments of Human Duodenal Epithelium
2.2. High Amounts of ERp57 Are Localized in the ER of RACE
2.3. Reduced Abundance of TG2 Inhibitory ERp57 within the ER of CD Biopsies
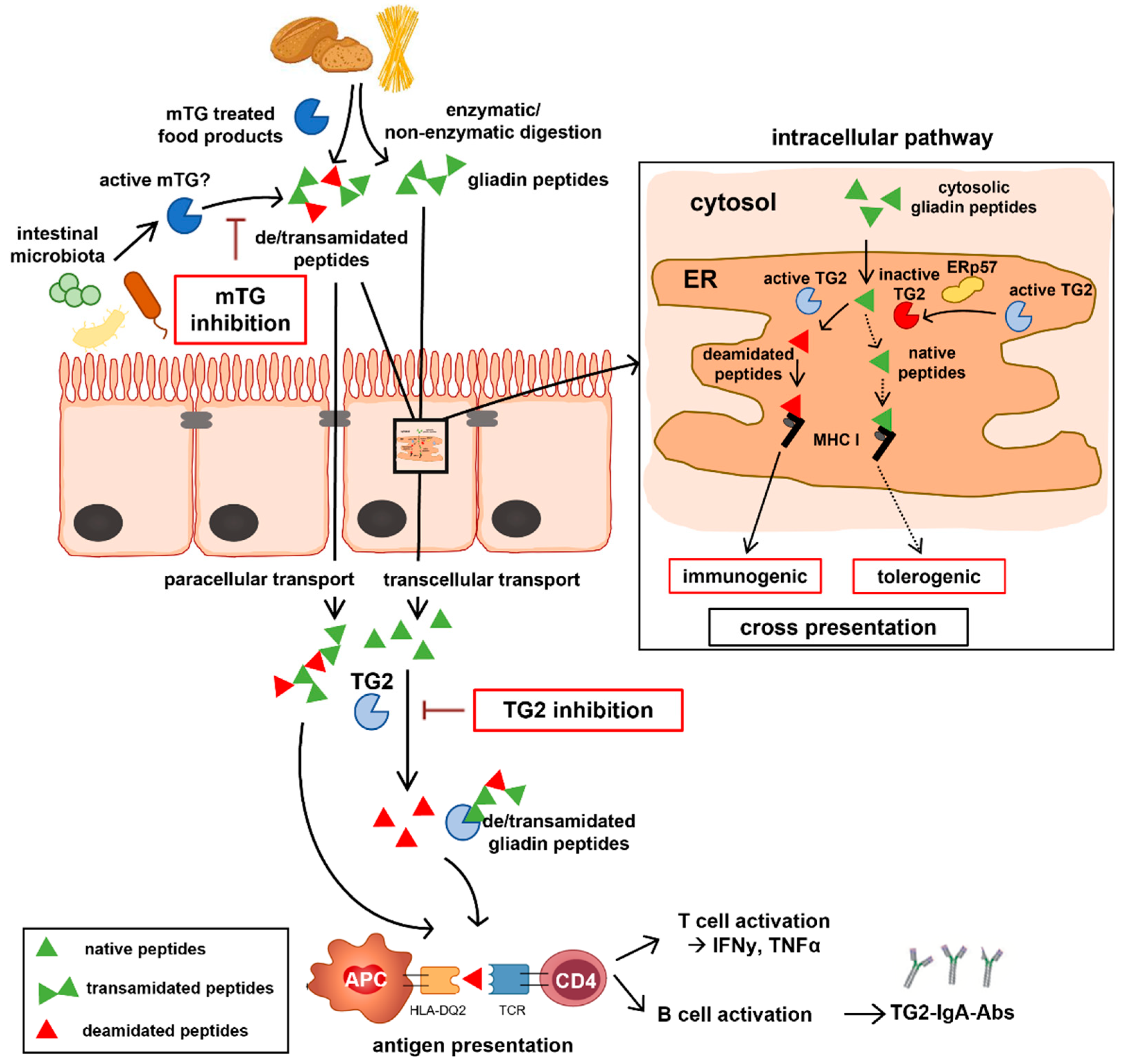
2.4. TG2 Is Regulated by Endogenous Oxidative Molecules
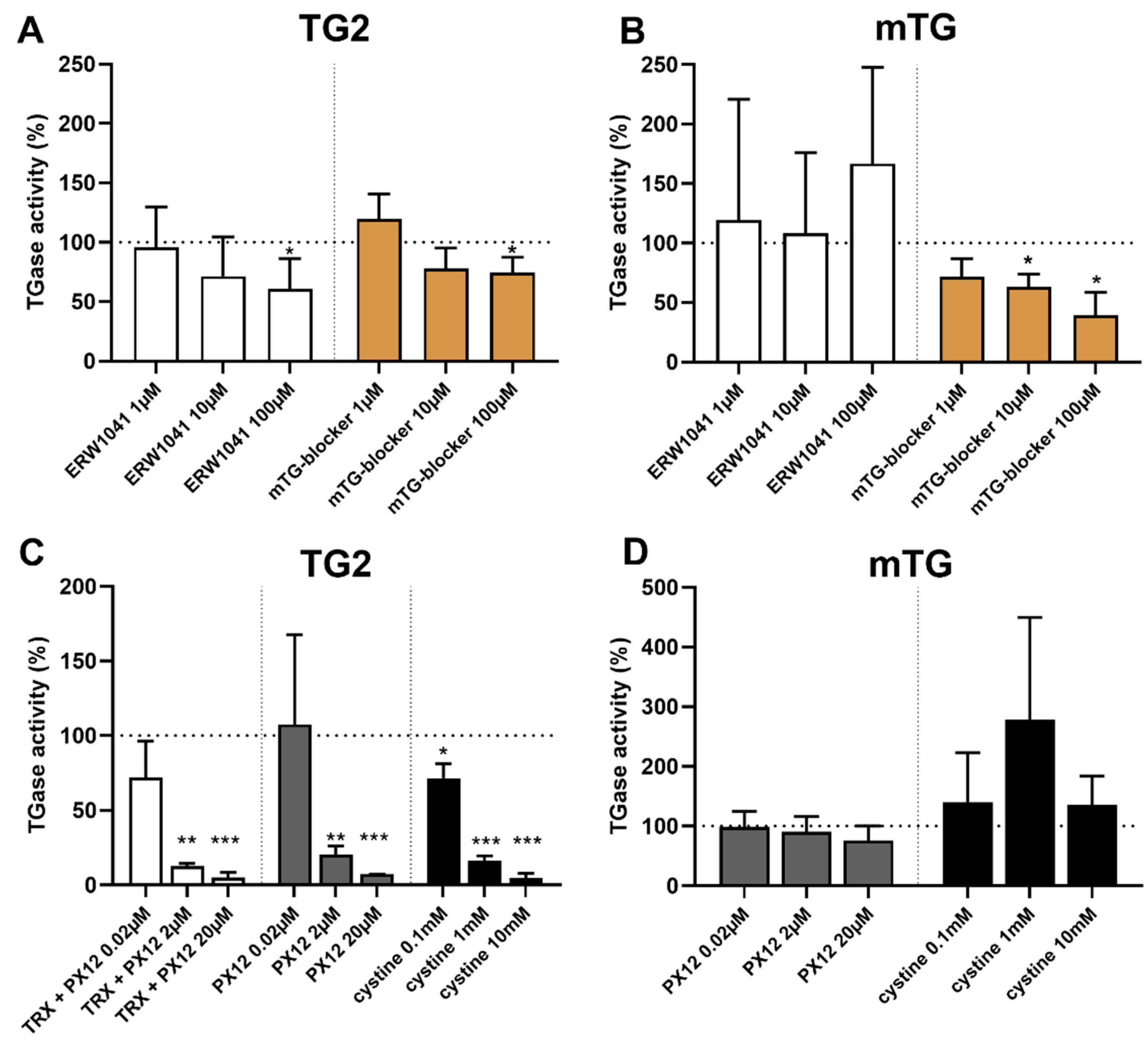
2.5. Selective Inhibition of TG2 and mTG by Exogenous Competitive Inhibitors
2.6. Oxidative Inhibition of TG2 by Exogenous PX12 and Cystine
3. Discussion
4. Materials and Methods
4.1. Patients’ Characteristics and Preparation of Biopsies
4.2. Electron Microscopy on Ultrathin Frozen Sections
4.3. Quantification of Subcellular Localization of ERp57
4.4. In Vitro Transamidation Activity of TG2 and mTG
4.5. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lebwohl, B.; Sanders, D.S.; Green, P.H.R. Coeliac disease. Lancet 2018, 391, 70–81. [Google Scholar] [CrossRef]
- Dieterich, W.; Ehnis, T.; Bauer, M.; Donner, P.; Volta, U.; Riecken, E.O.; Schuppan, D. Identification of tissue transglutaminase as the autoantigen of celiac disease. Nat. Med. 1997, 3, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Molberg, O.; McAdam, S.N.; Korner, R.; Quarsten, H.; Kristiansen, C.; Madsen, L.; Fugger, L.; Scott, H.; Norén, O.; Roepstorff, P.; et al. Tissue transglutaminase selectively modifies gliadin peptides that are recognized by gut-derived T cells in celiac disease. Nat. Med. 1998, 4, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Jabri, B.; Sollid, L.M. T Cells in Celiac Disease. J. Immunol. 2017, 198, 3005–3014. [Google Scholar] [CrossRef]
- Dekking, E.H.A.; van Veelen, P.A.; De Ru, A.A.; Kooy-Winkelaar, E.M.C.; Gröneveld, T.; Nieuwenhuizen, W.F.; Koning, F. Microbial transglutaminases generate T cell stimulatory epitopes involved in celiac disease. J. Cereal. Sci. 2008, 47, 339–346. [Google Scholar] [CrossRef]
- Skovbjerg, H.; Koch, C.; Anthonsen, D.; Sjöström, H. Deamidation and cross-linking of gliadin peptides by transglutaminases and the relation to celiac disease. Biochim. Biophys. Acta. 2004, 169, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Heil, A.; Ohsam, J.; Büchold, C.; Heil, A.; Ohsam, J.; Büchold, C.; Pasternack, R.; Yokoyama, K.; Kumazawa, Y.; Hils, M. Microbial transglutaminase has a lower deamidation preference than human tissue transglutaminase on a celiac disease relevant wheat gliadin T-cell epitope. J. Cereal. Sci. 2016, 70, 47–56. [Google Scholar] [CrossRef]
- Kieliszek, M.; Misiewicz, A. Microbial transglutaminase and its application in the food industry. A review. Folia Microbiol. 2014, 59, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Directive 2000/13/EC of the European Parliament and of the Council of 20 March 2000 on the Approximation of the Laws of the Member States Relating to the Labelling, Presentation and Advertising of Foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/DE/ALL/?uri=CELEX%3A32000L0013 (accessed on 25 July 2021).
- Lerner, A.; Aminov, R.; Matthias, T. Transglutaminases in Dysbiosis As Potential Environmental Drivers of Autoimmunity. Front. Microbiol. 2017, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.C.; Melkonian, A.V.; Ousey, J.A.; Khosla, C. Endoplasmic reticulum-resident protein 57 (ERp57) oxidatively inactivates human transglutaminase 2. J. Biol. Chem. 2018, 293, 2640–2649. [Google Scholar] [CrossRef] [Green Version]
- Plugis, N.M.; Palanski, B.A.; Weng, C.-H.; Albertelli, M.; Khosla, C.l. Thioredoxin-1 Selectively Activates Transglutaminase 2 in the Extracellular Matrix of the Small Intestine: IMPLICATIONS FOR CELIAC DISEASE. J. Biol. Chem. 2017, 292, 2000–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Stamnaes, J.; Klöck, C.; DiRaimondo, T.R.; Sollid, L.M.; Khosla, C. Activation of extracellular transglutaminase 2 by thioredoxin. J. Biol. Chem. 2011, 286, 37866–37873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiRaimondo, T.R.; Plugis, N.M.; Jin, X.; Khosla, C. Selective inhibition of extracellular thioredoxin by asymmetric disulfides. J. Med. Chem. 2013, 56, 1301–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turano, C.; Gaucci, E.; Grillo, C.; Chichiarelli, S. ERp57/GRP58: A protein with multiple functions. Cell. Mol. Biol. Lett. 2011, 1, 539–563. [Google Scholar] [CrossRef]
- Grommé, M.; Neefjes, J. Antigen degradation or presentation by MHC class I molecules via classical and non-classical pathways. Mol. Immunol. 2002, 39, 181–202. [Google Scholar] [CrossRef]
- Kovacsovics-Bankowski, M.; Rock, K.L. A Phagosome-to-Cytosol Pathway for Exogenous Antigens Presented on MHC Class I Mol. Sci. 1995, 267, 243–246. [Google Scholar] [CrossRef]
- Embgenbroich, M.; Burgdorf, S. Current Concepts of Antigen Cross-Presentation. Front. Immunol. 2018, 9, 1643. [Google Scholar] [CrossRef] [Green Version]
- Damo, M.; Wilson, D.S.; Watkins, E.A.; Hubbell, J.A. Soluble N-Acetylgalactosamine-Modified Antigens Enhance Hepatocyte-Dependent Antigen Cross-Presentation and Result in Antigen-Specific CD8+ T Cell Tolerance Development. Front. Immunol. 2021, 12, 555095. [Google Scholar] [CrossRef]
- Heath, W.R.; Carbone, F.R. Cross-presentation in viral immunity and self-tolerance. Nat. Rev. Immunol. 2001, 1, 126–134. [Google Scholar] [CrossRef]
- Gebru, Y.A.; Gupta, H.; Kim, H.S.; Eom, J.A.; Kwon, G.H.; Park, E.; Jeong, J.-J.; Won, S.-M.; Sharma, S.P.; Ganesan, R.; et al. T Cell Subsets and Natural Killer Cells in the Pathogenesis of Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2021, 22, 12190. [Google Scholar] [CrossRef]
- Ackerman, A.L.; Cresswell, P. Cellular mechanisms governing cross-presentation of exogenous antigens. Nat. Immunol. 2004, 5, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Stricker, S.; Laffolie, J.; de Rudloff, S.; Komorowski, L.; Zimmer, K.P. Intracellular Localization of Microbial Transglutaminase and Its Influence on the Transport of Gliadin in Enterocytes. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G. Fine Structure Immunocytochemistry; Springer: Berlin, Germany, 1993. [Google Scholar]
- Tye-Din, J.A.; Galipeau, H.J.; Agardh, D. Celiac Disease: A Review of Current Concepts in Pathogenesis, Prevention, and Novel Therapies. Front. Pediatr. 2018, 6, 350. [Google Scholar] [CrossRef] [PubMed]
- Arentz-Hansen, H.; Korner, R.; Molberg, O.; Quarsten, H.; Vader, W.; Kooy, Y.M.C.; Lundin, K.E.A.; Koning, F.; Roepstorff, P.; Sollid, L.M.; et al. The Intestinal T Cell Response to alpha-Gliadin in Adult Celiac Disease is Focused on a Single Deamidated Glutamine Targeted by Tissue Transglutaminase. J. Exp. Med. 2000, 191, 603–612. [Google Scholar] [CrossRef]
- Mazzarella, G.; Stefanile, R.; Camarca, A.; Giliberti, P.; Cosentini, E.; Marano, C.; Iaquinto, G.; Giardullo, N.; Auricchio, S.; Sette, A.; et al. Gliadin activates HLA class I-restricted CD8+ T cells in celiac disease intestinal mucosa and induces the enterocyte apoptosis. Gastroenterology 2008, 134, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Fan, S.; Zheng, J.; Huang, W.; Gasparetto, C.; Chao, N.J.; Hu, J.; Kang, Y. Inhibition of thioredoxin activates mitophagy and overcomes adaptive bortezomib resistance in multiple myeloma. J. Hematol. Oncol. 2018, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, R.K.; Kirkpatrick, D.L.; Belani, C.P.; Friedland, D.; Green, S.B.; Chow, H.S.; .Cordova, C.A.; Stratton, S.P.; Sharlow, E.R.; Baker, A.; et al. A Phase I pharmacokinetic and pharmacodynamic study of PX-12, a novel inhibitor of thioredoxin-1, in patients with advanced solid tumors. Clin. Cancer Res. 2007, 13, 2109–2114. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.F.; Dragovich, T.; Tate, W.R. The antitumor thioredoxin-1 inhibitor PX-12 (1-methylpropyl 2-imidazolyl disulfide) decreases thioredoxin-1 and VEGF levels in cancer patient plasma. J. Lab. Clin. Med. 2006, 147, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Sato, R.; Komura, T.; Ichikawa, H.; Hirashima, T.; Otake, S.; Akazawa, N.; Yazawa, T.; Abe, T.; Okada, T.; et al. Protective effect of the oral administration of cystine and theanine on oxaliplatin-induced peripheral neuropathy: A pilot randomized trial. Int. J. Clin. Oncol. 2020, 25, 1814–1821. [Google Scholar] [CrossRef]
- Miyachi, T.; Tsuchiya, T.; Oyama, A.; Tsuchiya, T.; Abe, N.; Sato, A.; Chiba, Y.; Kurihara, S.; Shibakusa, T.; Mikami, T. Perioperative oral administration of cystine and theanine enhances recovery after distal gastrectomy: A prospective randomized trial. J. Parenter. Enteral. Nutr. 2013, 37, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Schuppan, D.; Mäki, M.; Lundin, K.E.A.; Isola, J.; Friesing-Sosnik, T.; Taavela, J.; Popp, A.; Koskenpato, J.; Langhorst, J.; Hovde, Ø.; et al. Randomized Trial of a Transglutaminase 2 Inhibitor for Celiac Disease. N. Engl. J. Med. 2021, 385, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, B.; Molberg, Y.; Qiao, S.W.; Schmid, D.G.; von der Mu¨lbe, F.; Elgstøen, K.; Sollid, L.M. Gliadin T cell epitope selection by tissue transglutaminase in Celiac disease—Role of enzyme specificity and pH influence on the transamidation versus deamidation reactions. J. Biol. Chem. 2002, 277, 34109–34116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piper, J.L.; Gray, G.M.; Khosla, C. High selectivity of human tissue transglutaminase for immunoactive gliadin peptides: Implications for celiac sprue. Biochemistry 2002, 41, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Adachi, M.; Umeda, K.; Matsuura, A.; Nonaka, M.; Uchio, R.; Tanaka, J.; Motoki, M. Purification and Characteristics of a Novel Transglutaminase Derived from Microorganisms. Agric. Biol. Chem. 1989, 53, 2613–2617. [Google Scholar]
- Gianfrani, C.; Siciliano, R.A.; Facchiano, A.M.; Camarca, A.; Mazzeo, M.F.; Costantini, S.; Salvati, V.M.; Maurana, F.; Mazzarella, G.; Iaquinto, G.; et al. Transamidation of wheat flour inhibits the response to gliadin of intestinal T cells in celiac disease. Gastroenterology 2007, 133, 780–789. [Google Scholar] [CrossRef]
- Heredia-Sandoval, N.G.; Islas-Rubio, A.R.; Cabrera-Chavez, F.; de la Barca, A.M.C. Transamidation of gluten proteins during the bread-making process of wheat flour to produce breads with less immunoreactive gluten. Food Funct. 2014, 5, 1813–1818. [Google Scholar] [CrossRef]
- Luongo, D.; Maurano, F.; Bergamo, P.; Rossi, M. Microbial transglutaminase: A biotechnological tool to manage gluten intolerance. Anal. Biochem. 2020, 592, 113584. [Google Scholar] [CrossRef]
- Mazzeo, M.F.; Bonavita, R.; Maurano, F.; Bergamo, P.; Siciliano, R.A.; Rossi, M. Biochemical modifications of gliadins induced by microbial transglutaminase on wheat flour. Biochim. Biophys. Acta 2013, 1830, 5166–5174. [Google Scholar] [CrossRef]
- Husby, S.; Koletzko, S.; Korponay-Szabo, I.R.; Mearin, M.L.; Phillip, S.A.; Shamir, R.; Troncone, R.; Gierspiepen, K.; Branski, D.; Catassi, C.; et al. European Society for Pediatric Gastroenterology, Hepatology, and Nutrition guidelines for the diagnosis of coeliac disease. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 136–160. [Google Scholar] [CrossRef]
- Husby, S.; Koletzko, S.; Korponay-Szabó, I.; Kurppa, K.; Mearin, M.L.; Ribes-Koninckx, C.; Shamir, R.; Troncone, R.; Auricchio, R.; Castillejo, G.; et al. European Society Paediatric Gastroenterology, Hepatology and Nutrition Guidelines for Diagnosing Coeliac Disease 2020. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Tokuyasu, K.T. A technique for ultracryotomy of cell suspensions and tissues. J. Cell. Biol. 1973, 57, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Kersting, S.; Bruewer, M.; Schuermann, G.; Klotz, A.; Utech, M.; Hansmerten, M.; Krieglstein, C.F.; Senninger, N.; Schulzke, J.-D.; Naim, H.Y.; et al. Antigen Transport and Cytoskeletal Characteristics of a Distinct Enterocyte Population in Inflammatory Bowel Diseases. Am. J. Pathol. 2004, 165, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Schürmann, G.; Brüwer, M.; Klotz, A.; Schmid, K.W.; Senninger, N.; Zimmer, K.P. Transepithelial transport processes at the intestinal mucosa in inflammatory bowel disease. Int. J. Colorectal. Dis. 1999, 14, 41–46. [Google Scholar] [CrossRef] [PubMed]
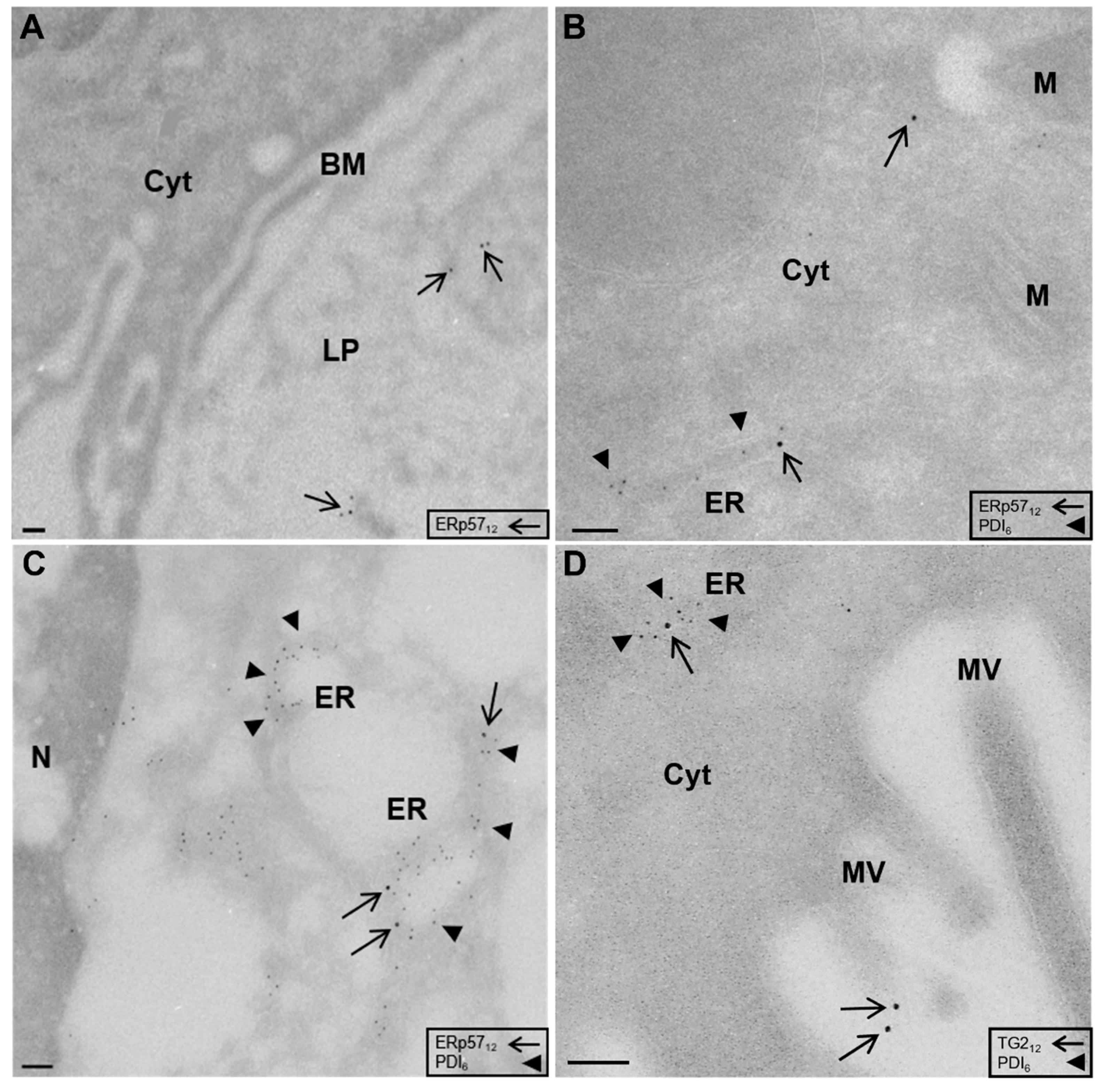



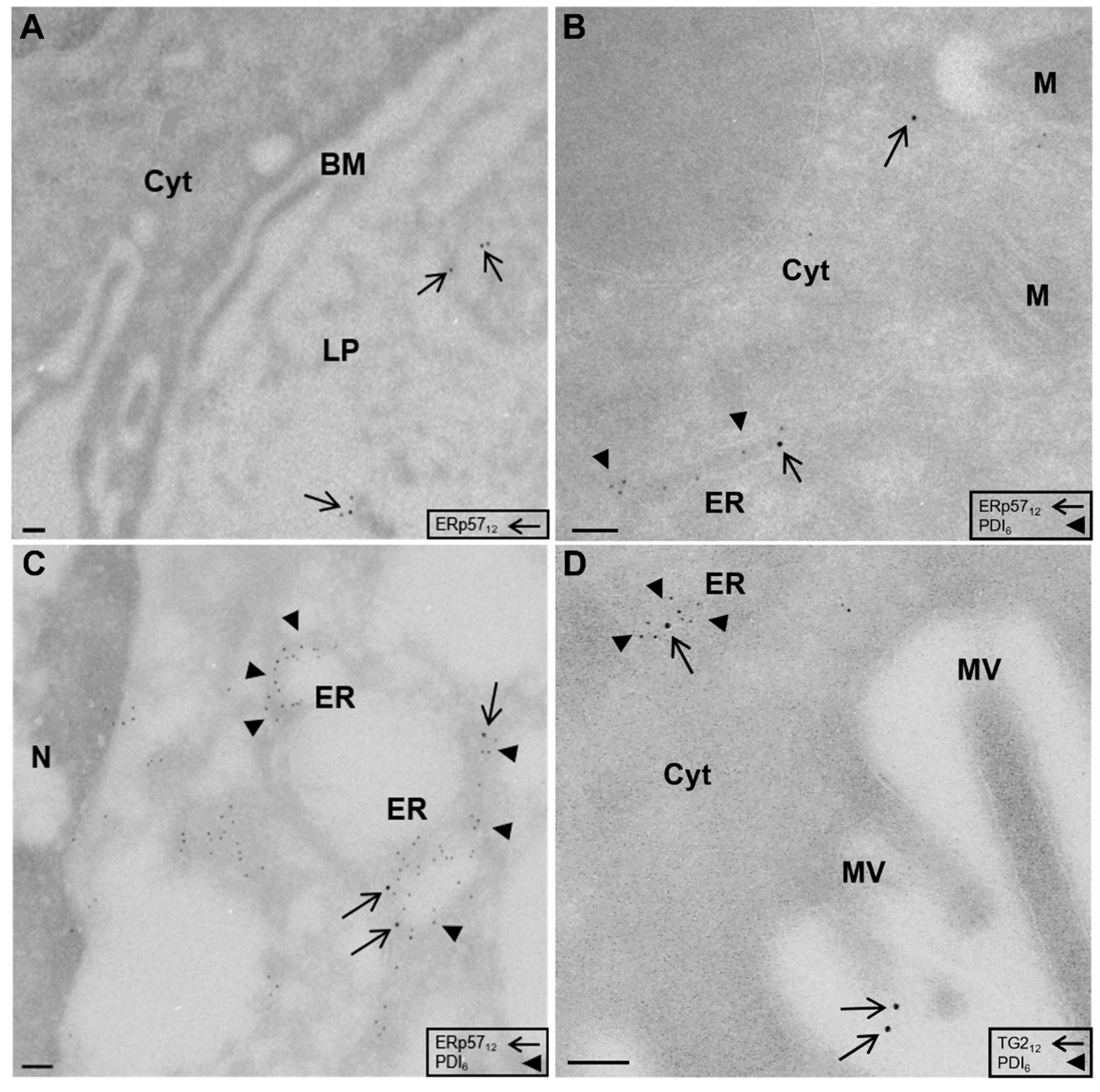{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enterocytes | RACE | Statistics | |
|---|---|---|---|
| labels per cell | median: 10, IQR 4 | median: 18, IQR 40 | p < 0.001 |
| surface of the ER | median: 0.7, IQR 0.4 µm | median 4.6, IQR 3 µm | p < 0.001 |
| apical membrane | median: 5, IQR 4% | median: 1, IQR 3% | p < 0.01 |
| vacuoles | mean: 37 ± 11% | mean: 45 ± 10% | p = 0.1 |
| endoplasmic reticulum | mean: 18 ± 4% | mean: 30 ± 7% | p < 0.001 |
| basal membrane | median: 10, IQR 28% | median: 0, IQR 11% | p < 0.01 |
| cytosol | mean: 23 ± 6% | mean: 15 ± 7% | p < 0.01 |
| Substance | Effect | Company |
|---|---|---|
| DTT (Dithiothreitol) | Stimulation/reduction | Sigma-Aldrich, St. Louis, MO, USA |
| TRX | Stimulation/reduction | Abcam, Cambridge, UK |
| L-cystine | Inhibition/oxidation | Sigma-Aldrich, St. Louis, MO, USA |
| ERp57 | Inhibition/oxidation | Novus Biologicals, Littleton, CO, USA |
| ERW1041 | Inhibition/competition | Sigma-Aldrich, St. Louis, MO, USA |
| mTG-blocker | Inhibition/competition | Zedira, Darmstadt, Germany |
| PX12 | Inhibition/oxidation | Sigma-Aldrich, St. Louis, MO, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stricker, S.; Rudloff, S.; De Laffolie, J.; Zimmer, K.-P. Tissue Transglutaminase but Not Microbial Transglutaminase Is Inhibited by Exogenous Oxidative Substances in Celiac Disease. Int. J. Mol. Sci. 2022, 23, 2248. https://doi.org/10.3390/ijms23042248
Stricker S, Rudloff S, De Laffolie J, Zimmer K-P. Tissue Transglutaminase but Not Microbial Transglutaminase Is Inhibited by Exogenous Oxidative Substances in Celiac Disease. International Journal of Molecular Sciences. 2022; 23(4):2248. https://doi.org/10.3390/ijms23042248
Chicago/Turabian StyleStricker, Sebastian, Silvia Rudloff, Jan De Laffolie, and Klaus-Peter Zimmer. 2022. "Tissue Transglutaminase but Not Microbial Transglutaminase Is Inhibited by Exogenous Oxidative Substances in Celiac Disease" International Journal of Molecular Sciences 23, no. 4: 2248. https://doi.org/10.3390/ijms23042248
APA StyleStricker, S., Rudloff, S., De Laffolie, J., & Zimmer, K. -P. (2022). Tissue Transglutaminase but Not Microbial Transglutaminase Is Inhibited by Exogenous Oxidative Substances in Celiac Disease. International Journal of Molecular Sciences, 23(4), 2248. https://doi.org/10.3390/ijms23042248